Марк Д. Фершильд. Модели цветового восприятия
Подождите немного. Документ загружается.


ка снабжает сигналами одну ганглиозную клетку, тогда как палочки, посылая
сигнал ганглиозной клетке, объединяют ответ от сотен рецепторов, обеспечи
вая тем самым повышенную чувствительность (но за счет остроты зрения). Ска
занное частично объясняет и то, каким образом информация от такого большо
го количества рецепторов передается через один миллион ганглиозных клеток.
Рис. 1.6 иллюстрирует также высокую концентрацию колбочковых рецепто
ров в центральной ямке. Колбочки крайне редко встречаются в периферийной
сетчатке, в то время как в центральной ямке совсем не встречаются палочки.
Благодаря отсутствию палочек, в центральной ямке освобождается полезное
пространство, используемое для обеспечения высокой остроты зрения при по
мощи колбочковой системы.
И последняя особенность, которую иллюстрирует рис. 1.6, это слепое пят
но — область, лежащая на 12–15° латеральнее центральной ямки, в которой на
чинается зрительный нерв и в которой вследствие этого отсутствуют фоторе
цепторы. Одна из причин, по которой слепое пятно остается незамечаемым, со
стоит в том, что оно находится на противоположных сторонах поля зрения в ка
ждом из двух глаз. Однако любопытно, что, когда один глаз закрыт, слепое пят
но попрежнему остается незаметным.
Рис. 1.7 можно использовать для доказательства существования слепого
пятна. Чтобы увидеть слепое пятно, закройте левый глаз, а взгляд правого за
фиксируйте на кресте рисунка 1.7 (а), затем меняйте расстояние до книжки,
пока точка справа от креста не исчезнет, попав на слепое пятно. Обратите вни
мание на то, что вы видите, когда исчезает точка: вместо темной области появ
ляется участок чистой бумаги — это пример феномена, именуемого заполнени
ем: поскольку мозг с данной позиции больше не получает сигнала об изменении
зрительного стимула, он заполняет пустоту наиболее вероятным стимулом,
в данном случае — однородной белой бумагой. Сила такого заполнения может
39
ГЛАВА 1 ЦВЕТОВОЕ ЗРЕНИЕ ЧЕЛОВЕКА
Рис. 1.6 Плотность (рецепторы на кв. мм) палочковых и колбочковых фоторецепторов как
функция залегания в сетчатке человека.

быть проиллюстрирована рис. 1.7 (b), предназначенным для исследования сле
пого пятна: закройте левый глаз, а взгляд правого зафиксируйте на кресте, за
тем меняйте расстояние просмотра до тех пор, пока прорезь в линии не исчез
нет, то есть пока она не попадет на слепое пятно, — вместо прорези вы будете
видеть непрерывную линию, поскольку теперь это наиболее вероятный зри
тельный стимул. Если вы хотите выполнить эти упражнения, закрыв левый
глаз, то просто переверните рисунок вверх ногами, чтобы поймать слепое пятно
на другой стороне поля зрения.
Феномен заполнения углубляет наше понимание механизмов работы зри
тельной системы: сигналы, переданные ганглиозным клеткам, несут информа
цию только о локальных изменениях в зрительных стимулах, причем дейст
венна лишь информация о пространственных и временн
¢
ых переходах (то есть
о границах), передаваемая в мозг. Восприятием этот код интерпретируется пу
тем анализа характера изменений и заполняется соответствующим однород
ным восприятием до поступления сигнала о новом переходе, что обеспечивает
гигантскую экономию в полосе пропускания передаваемых сигналов. Можно
даже говорить о некоем подобии группового кодирования (сжатия), которое
иногда применяется в технологии цифровых изображений.
1.3 ОБРАБОТКА ЗРИТЕЛЬНОГО СИГНАЛА
Процесс нейронной обработки зрительной информации на уровне сетчатки
сложен, а на дальнейших стадиях он становится значительно (если не бесконеч
но) более сложным. Данный раздел — это краткий обзор путей, по которым идет
зрительная информация, и имеет смысл начать его с общей карты этих путей.
Оптическое изображение на сетчатке вначале преобразуется в химические
и электрические сигналы фоторецепторов, а затем эти сигналы обрабатывают
ся клетками нейронной сети сетчатки (горизонтальными, биполярными, амак
риновыми и ганглиозными клетками, описанными выше). Аксоны ганглиоз
ных клеток формируют зрительный нерв, который идет к т.н. латеральному
40
ГЛАВА 1 ЦВЕТОВОЕ ЗРЕНИЕ ЧЕЛОВЕКА
Рис. 1.7 Стимулы, используемые для иллюстрации наличия слепого пятна и феномена «запол
нения». Закройте левый глаз. Зафиксируйте взгляд правого глаза на кресте и меняйте дистан
цию просмотра до тех пор, пока точка (а) или прорезь в линии (b) не упадeт на ваше слепое пят
но. Обратите внимание на ощущения в каждом случае.
коленчатому телу (lateral geniculate nucleus — LGN) в таламусе
1
. Клетки LGN,
получив входной сигнал от ганглиозных клеток, отсылают его в т.н. первую
зрительную область (V1) затылочных долей коры головного мозга, в которых
обработка информации становится невероятно сложной. В коре головного моз
га было найдено почти 30 зрительных областей, названные затем как V2, V3,
V4, MT и т.д. Сигналы от этих областей отсылаются к некоторым другим облас
тям и обратно. Корковая обработка включает в себя множество стадий: пря
мую, обратную и латеральную обработки; гдето в недрах этой сети и формиру
ется наше окончательное восприятие. Коекакие детали этих процессов описа
ны в следующих параграфах.
Свет, воздействующий на сетчатку, абсорбируется фотопигментом различ
ных фоторецепторов (в палочках, в частности, это родопсин). После поглоще
ния фотона родопсин меняет свою структуру, происходит цепная химическая
реакция, итогом которой является закрытие ионных каналов в клеточной стен
ке, продуцирующее электрический сигнал, основанный на разности относи
тельных концентраций различных ионов (к примеру, натриевых и карбонат
ных) вне и внутри клетки. Аналогичные процессы протекают в колбочках, ро
допсин которых состоит из опсина и ретиналя: колбочки всех трех типов име
ют сходную фотопигментную структуру, но у колбочковых опсинов («конопси
нов») немного разные молекулярные формулы, что ведет к отличиям в спек
тральной чувствительности колбочек. Каждый тип колбочек (L, M или S)со
держит свою форму «конопсина».
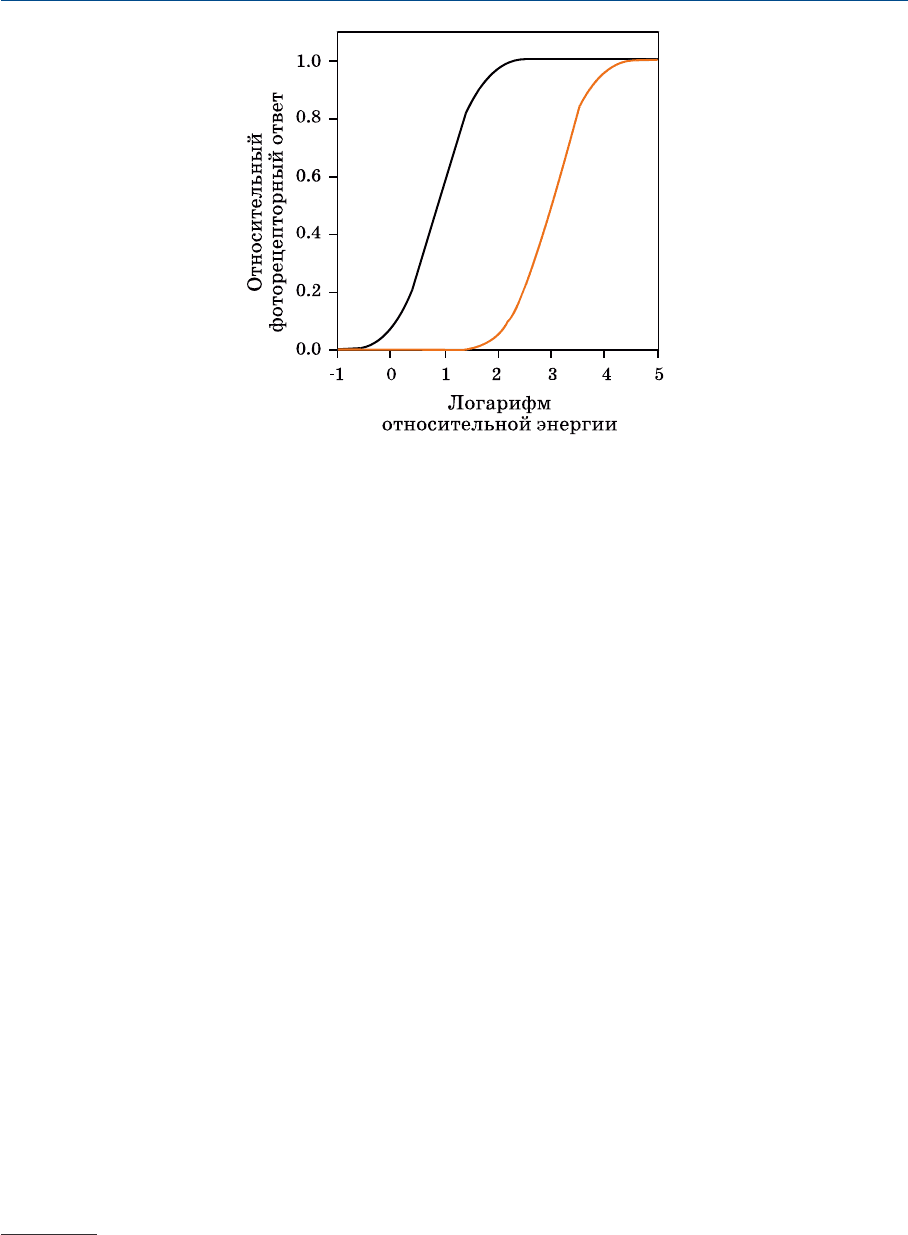
На рис. 1.8 показаны относительные ответы фоторецепторов как функция
световой экспозиции сетчатки. Любопытно, что характеристики этих функций
подобны характеристикам многих устройств ввода и вывода изображений.
41
ГЛАВА 1 ЦВЕТОВОЕ ЗРЕНИЕ ЧЕЛОВЕКА
Рис. 1.8 Относительные энергетические ответы палочковых и колбочковых фоторецепторов.
1
Таламус — подкорковый отдел головного мозга. Латеральное коленчатое тело — подструк
тура таламуса. — Прим. пер.

В нижней части графика виден порог, ниже которого рецепторы не отвечают.
Затем следует более или менее линейный участок кривых, и за ним — финаль
ный участок — зона насыщения (верхний конец кривых). Приведенные кри
вые описывают рецепторный фототок и демонстрируют самую первую стадию
зрительной обработки. В дальнейшем сигналы обрабатываются нейронами сет
чатки и синапсами. Результатом этой обработки является преобразованный
сигнал, который ганглиозные клетки отправляют по зрительному нерву в мозг.
Рецептивные поля
По ряду причин (таких, как необходимость подавления шумов и обеспече
ние высокой скорости передачи) амплитудномодулированные сигналы фото
рецепторов конвертируются ганглиозными клетками и вышестоящими отде
лами мозга в частотномодулированное представление. Сегодня несомненно то,
что величина сигнала от возбужденной нервной клетки представлена количе
ством пиков напряжения в секунду, а не разностью потенциалов по обе стороны
клеточной стенки.
Чтобы получить представление о физиологических свойствах нервных кле
ток сетчатки, необходимо познакомиться с концецпией т.н. рецептивных полей.
Рецептивное поле фоторецептора — это графическое представление неко
ей области поля зрения, на которую отвечает данная клетка. На схеме рецеп
тивного поля обычно указывают характер ответа различных областей этого
поля (к примеру, положительный ответ, отрицательный ответ, спектральный
сдвиг). Фактически рецептивное поле фоторецептора — это маленькая круглая
область, указывающая размер и местоположение персональной чувствитель
ности данного рецептора (в поле зрения).
На рис. 1.9 показаны некоторые варианты рецептивных полей ганглиозных
клеток, иллюстрирующие антагонизм центра и окружения, характерный для
данного уровня зрительной обработки. Рецептивное поле на рис. 1.9 (а) иллю
стрирует положительный центральный ответ на положительный входной сиг
нал от единичной колбочки: положительный ответ окружен т.н. отрицатель
ным ответом окружения, который, как правило, запускается отрицательным
входным сигналом от соседних колбочек. Таким образом, ответ от данной ганг
42
ГЛАВА 1 ЦВЕТОВОЕ ЗРЕНИЕ ЧЕЛОВЕКА
Рис. 1.9 Типичные антагонистичные рецептивные поля «центр — окружение»: (а) — централь
ное; (b) периферийное.

лиозной клетки задается совокупностью положительных и отрицательных
входных сигналов от множества колбочек, в результате чего ганглиозная клет
ка не просто отвечает на точечный свет, но выступает в роли т.н. краевого дат
чика (фактически — датчика «пятен»). Читатели, знакомые с цифровой обра
боткой изображений, могут провести аналогию между ответом ганглиозной
клетки и результатом операции свертки с заданным ядром, применяемой для
обнаружения края.
На рис. 1.9 (b) показано, что ответы с противоположными полярностями рав
новероятны: ответ, показанный на рис. 1.9 (а), рассматривают как ответ цен
тральной ганглиозной клетки, тогда как ответ, показанный на рис. 1.9 (b), — на
зывают ответом периферийной ганглиозной клетки. Зачастую центральные
и периферийные клетки имеют одну и ту же пространственную локализацию,
питаемую одними и теми же фоторецепторами, в результате чего повышается
динамический диапазон системы.
Отметим, что ганглиозные клетки, показанные на рис. 1.9, не будут выда
вать ответ на однородные поля (то есть тогда, когда положительные и отрица
тельные сигналы сбалансированы). Сказанное выше иллюстрирует один из ас
пектов компрессии изображения, выполняемого сетчаткой, благодаря которо
му головной мозг не перегружается избыточной информацией — передаются
данные лишь об изменениях в окружающем мире. Отметим, что пространст
венная обработка информации в зрительной системе — это базис влияния фона
на цветовое восприятие.
Рис. 1.9 иллюстрирует пространственную оппонентность ответов ганглиоз
ных клеток, а на рис. 1.10 показано, что в дополнение к пространственной оп
понентности к ответам ганглиозных клеток часто прибавляется спектральная
оппонентность. На рис. 1.10 (а) показан краснозеленый оппонентный ответ
в центре, питаемый положительным сигналом от Lколбочки, при этом окру
жение питаемо отрицательным сигналом от Mколбочек. На рис. 1.10 (b) пока
зана периферийная версия данной клетки. Таким образом, прежде чем зри
тельная информация покинет сетчатку, происходит ее обработка, радикально
влияющая на цветовое восприятие. Рисунки 1.9 и 1.10 демонстрируют типич
ные рецептивные поля ганглиозных клеток, и хотя существует множество
43
ГЛАВА 1 ЦВЕТОВОЕ ЗРЕНИЕ ЧЕЛОВЕКА
Рис. 1.10 Примеры краснозеленых (а) и зеленокрасных (b) спектрально и пространственно
антагонистичных рецептивных полей.

иных типов и вариантов ответа ганглиозных клеток, все они работают по сход
ным принципам.
На пути к первичной зрительной коре сигналы проходят через латеральное
коленчатое тело (LGN). Аксоны ганглиозных клеток заканчиваются в LGN,
создавая с его клетками синапсы, что обеспечивает непосредственный («один
в один») обмен информацией между ганглиозными и LGNклетками. Таким об
разом, рецептивные поля LGNклеток идентичны таковым у ганглиозных кле
ток. Возможно, LGN действует только как ретранслятор сигналов. Однако ве
роятно и то, что LGN все же выполняет некую зрительную функцию, принимая
на себя нейронные проекции из коры, которые, в свою очередь, могут отвечать
за некоторые виды коммутации или за адаптационный механизм обратной свя
зи. Аксоны LGNклеток подходят к первой зрительной области коры головного
мозга (V1).
Обработка в V1;области
В V1области коры головного мозга кодирование зрительной информации
существенно усложняется: подобно тому как исходящие сигналы от различ
ных фоторецепторов сравниваются и объединяются для реализации ответов
ганглиозной клетки, выходящие сигналы от LGNклеток сравниваются и объе
диняются для производства кортикальных (корковых) ответов. Когда сигналы
попадают в цепь кортикальной обработки, эта обработка повторяется несколь
ко раз с резко возрастающим уровнем сложности, и в итоге рецептивные поля
начинают терять свое значение.
В V1 встречаются клетки, которые отвечают лишь на стимулы определенно
го вида, например:
— края и полоски различной ориентации;
— вход от одного глаза, второго и обоих глаз;
— различные пространственные частоты;
— различные временн
¢
ые частоты;
— специфические пространственные местоположения;
— комбинации перечисленных стимулов.
Кроме того, обнаруживаются клетки, которые линейно комбинируют сиг
налы от клеток LGN (и от прочих клеток с нелинейным суммированием).
Все варианты ответов необходимы для восприятия размера, формы, место
положения, движения, интенсивности и цвета. Учитывая сложность корти
кальных ответов в клетках V1, нетрудно представить себе, до какой степени
сложны зрительные ответы, получаемые из сети, состоящей примерно из
30 зрительных областей.
На рис. 1.11 показана схема малой доли связей между различными извест
ными кортикальными областями. Отметим, что рис. 1.11 демонстрирует связи
именно между областями, но не между клетками: при этом существует пример
но 109 нейронов коры, выполняющих персональные зрительные функции,
деятельность которых очень сложно объяснить простыми словами. Однако
функция отдельной клетки, как правило, не имеет значения, поскольку раз
44
ГЛАВА 1 ЦВЕТОВОЕ ЗРЕНИЕ ЧЕЛОВЕКА

личные виды восприятия распространяются на целые пакеты клеток, разбро
санных по всей коре головного мозга.
Более мы не будем говорить о физиологии и в следующих разделах опишем
лишь некоторые общие перцепционные и психофизические свойства зритель
ной системы, что поможет нам глубже понять механизм ее работы.
1.4 МЕХАНИЗМЫ ЦВЕТОВОГО ЗРЕНИЯ
В истории нашей науки было много теорий, пытавшихся объяснить меха
низмы цветового зрения. Краткий обзор некоторых наиболее современных
концепций поможет разобраться в концепции текущей.
Трихроматическая теория
В конце XIXго века была разработана т.н. трихроматическая теория цвето
вого зрения, основанная на трудах Максвелла, Юнга и Гельмгольца, которые
догадывались о существовании трех типов рецепторов, чувствительных к лу
чам примерно красной, примерно зеленой и примерно синей областей спектра.
Трихроматическая теория предполагала, что три вида рецепторов формируют
три изображения окружающего мира, которые затем передаются в мозг, где со
отношения сигналов каждого изображения сравниваются, на основании чего
и возникает цветовое ощущение. Трихроматическая (трехрецепторная) приро
да цветового зрения не вызывала сомнений, но идея трех изображений, посы
лаемых в мозг, оказалась несостоятельной и не смогла объяснить ряд зритель
ных феноменов.
45
ГЛАВА 1 ЦВЕТОВОЕ ЗРЕНИЕ ЧЕЛОВЕКА
Рис. 1.11 Фрагментарная диаграмма, иллюстрирующая многопоточность обработки визуаль
ной информации в зрительной коре головного мозга. Информация может передаваться по каж
дой связи в обоих направлениях.

Оппонентная теория Геринга
В тот же период Геринг предложил т.н. оппонентную теорию цветового зре
ния, основанную на множестве субъективных наблюдений за цветовым вос
приятием. Эти наблюдения включали восприятие цветового тона и симультан
ного (одновременного) контраста, т.н. постобразы и аномалии цветового зре
ния. Геринг отметил, что определенные цветовые тона никогда не воспринима
ются одновременно, к примеру: цветовое ощущение никогда не описывается
как зеленокрасное или желтосинее, в то время как комбинации красного
с желтым, красного с синим, зеленого с желтым и зеленого с синим восприни
маются легко. Это обстоятельство навело Геринга на мысль, что существует не
кая фундаментальная причина, по которой цвета в краснозеленых и желто
синих парах противопоставлены друг другу. Наблюдения касались и симуль
танного контраста, суть которого в том, что одни и те же объекты, расположен
ные на красном фоне, воспринимаются чуть более зелеными, на зеленом фоне
чуть более красными, на желтом фоне чуть более синими, а на синем фоне чуть
более желтыми.
Рис. 1.12 иллюстрирует оппонентную природу визуальных постобразов. По
стобраз от красного стимула будет зеленым, от зеленого — красным, от желто
го — синим и от синего — желтым (отметим, что постобразы легко объяснить
и в логике комплементарных [дополнительных] цветов как следствие адапта
ции трихроматической системы). Обосновывая оппонентную теорию, Геринг
упомянул лишь о светлотемных постобразах, но при этом ничего не сказал
о хроматических). Наконец, Геринг заметил, что лица, страдающие аномалия
46
ГЛАВА 1 ЦВЕТОВОЕ ЗРЕНИЕ ЧЕЛОВЕКА
Рис. 1.12 Стимулы, предназначенные для демонстрации оппонентных постобразов. Зафикси
руйте взгляд на черной точке в месте стыковки четырех окрашенных квадратов. Через 30 се
кунд переместите и зафиксируйте взгляд на черной точке равномерного белого поля. Обратите
внимание на цвета возникших постобразов и сравните их с цветами оригинальных стимулов.

ми цветового зрения, теряют способность различать цветовые тона в краcно
зеленых и желтосиних парах.
Наблюдения Геринга ведут нас к пониманию механизма обработки цветовой
информации зрительной системой. Геринг тоже предполагал существование
трех типов рецепторов, но считал, что они работают по принципу биполярного
ответа на светлыйтемный, красныйзеленый и желтыйсиний. В то время та
кое утверждение казалось физиологически абсурдным, и оппонентная теория
Геринга не получила признания.
Современная теория оппонентных цветов
В середине XXго века оппонентная теория возродилась, поскольку стали по
являться подтверждающие ее экспериментальные данные, к примеру: Свэтичин
(1956) обнаружил оппонентные сигналы при электрофизиологических измере
ниях ответов в сетчатке золотой рыбки (которые оказались трихроматически
ми!), Де`Валойс с коллегами (1958) обнаружил подобные оппонентные физиоло
гические ответы в LGNклетках макаки, Джеймсон и Гурвич (1955) представили
количественные психофизические данные, полученные на основе т.н. безотте
ночных экспериментов с людьми. Эксперименты Джеймсона и Гурвича позволи
ли измерить относительные спектральные чувствительности оппонентных пу
тей. Все эти данные, объединенные с огромным количеством дополнительных
исследований, привели к развитию современной оппонентной теории цветового
зрения, которую иногда называют стадийной теорией (см. рис. 1.13).
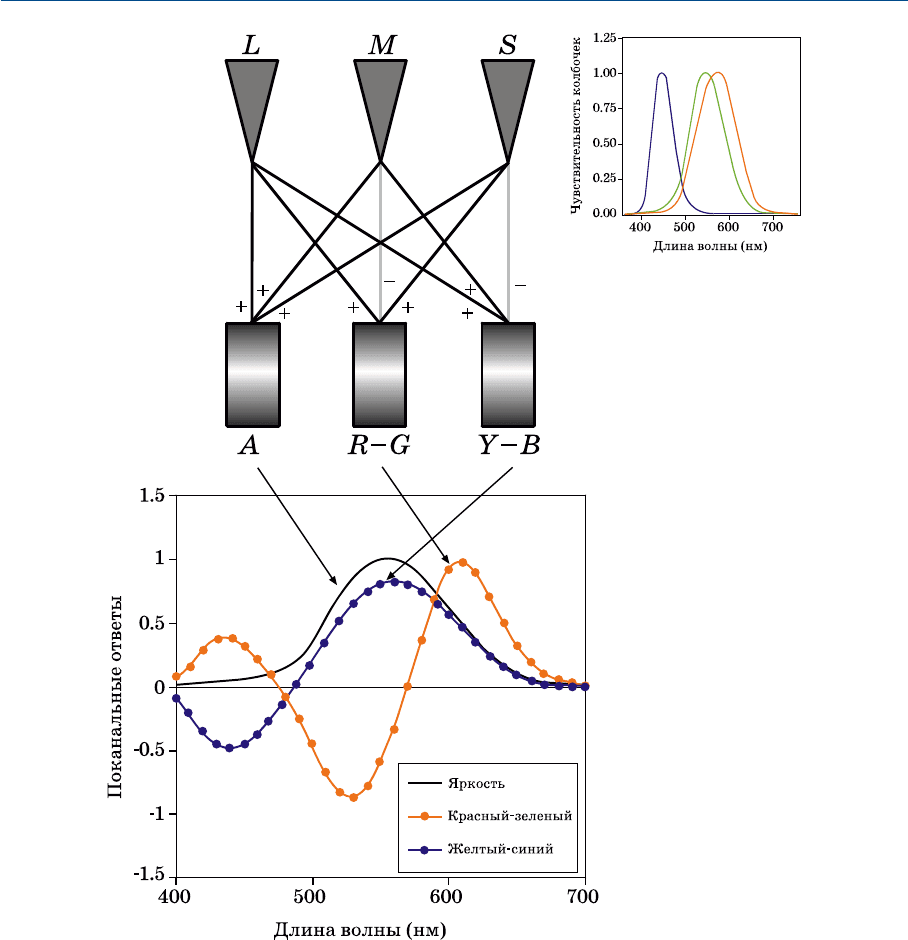
Рис. 1.13 демонстрирует, что на первой стадии цветового зрения рецепторы,
как и предполагали Максвелл, Юнг и Гельмгольц, действительно трихромати
ческие. Однако вопреки простой трихроматической теории три «цветоделен
ных» изображения не передаются непосредственно в мозг: вместо этого нейро
ны сетчатки (и возможно, более высокие отделы зрительной системы) кодиру
ют цвет через оппонентные сигналы. Выходные сигналы от всех трех типов
колбочек суммируются (L+M+S) с целью получения ахроматического ответа,
который до тех пор соответствует кривой CIE V(l), пока суммация берется
в пропорции к относительной заселенности сетчатки тремя типами колбочек.
Различия в колбочковых сигналах позволяют сконструировать красно
зеленые (L–M+S) и желтосиние (L+M–S) оппонентные сигналы.
Преобразование LMSсигналов в оппонентные приводит к декорреляции
цветовой информации, идущей по трем каналам, тем самым обеспечивая более
эффективную передачу сигналов и снижая шумовые помехи. Три оппонентных
пути, разумеется, имеют индивидуальные пространственные и временн
¢
ые ха
рактеристики, очень важные для прогнозирования цветового восприятия (но
их мы обсудим в разделе 1.5).
Значимость преобразования трихроматических сигналов в оппонентные от
ражена в формулировках всех моделей цветового восприятия. В рис. 1.13
включена не только схематическая диаграмма нейронного «телеграфа», кото
рый продуцирует оппонентные ответы, но также и относительные спектраль
ные чувствительности оппонентных механизмов до и после оппонентного коди
рования.
47
ГЛАВА 1 ЦВЕТОВОЕ ЗРЕНИЕ ЧЕЛОВЕКА
Механизмы адаптации
Заметим, что было бы неправомерно рассматривать обработку цветовых сиг
налов человеческой зрительной системой лишь как статическую: необходимо
еще разобрать динамические приспособительные механизмы, которые служат
оптимизации визуального ответа в тех или иных условиях просмотра, то есть
изучить различные виды адаптации.
Для понимания процессов цветового восприятия необходимо рассмотреть
механизмы темновой, световой и хроматической адаптаций.
48
ГЛАВА 1 ЦВЕТОВОЕ ЗРЕНИЕ ЧЕЛОВЕКА
Рис. 1.13 Схематическая иллюстрация кодирования колбочковых сигналов в оппонентные
цветовые сигналы в зрительной системе человека.