Сафонов А., Хоботкова Л., Хижняк Н. Загальна екологія
Подождите немного. Документ загружается.

61
Для пружньої стійкості кількісною характеристикою слугує сила впливу, після припинення якого
система здатна порвернутися до вихі дного стану, а також швидкість, з якою система повертається до
попереднього стану.
11.2. Механізми регуляції екологічних процесів
Всі біо- і екосистеми є саморегулівними системами зі здатністю до самоорганізації та
накопичення негентропії.
В основі регуляції будь-яких процесів лежить механізм зворотнього зв’язку. Якщо вихідні
параметри системи впливають на вхідні, то система регулюється петлею зворотнього зв’язку. Якщо
зростання вихідних параметрів призводить до зростання вхідних, має місце позитивний зворотній зв’язок,
у протилежному випадку – негативний. Реалізований механізм негативного зворотнього зв’язку ми маємо
нагоду споглядати в будові механізму регуляції рівня води в надунітазному бачкові. Коли рівень води в
ньому сягає мінімуму (при спусках води), максимально відчиняється отвір, з якого наповнюється водою
бачок. В міру його наповнення поплавок піднімається і через важіль зменшує отвір водотічного сифону, аж
поки не зачинить його зовсім. Тобто чим вищий рівень води в
бачку, тим повільніше йде його наповнення
водою, і нарешті при певному рівні тік води припиняється.
Позитивний зворотній зв’язок можна спостерігати на всіляких пожежах - чим більше полум’я
охоплює довкілля, тим швидше воно поширюється, аж поки не вигорить все, що здатне горіти, після чого
пожежа припиняється.
Стосовно живої матерії, то
позитивний зворотній зв’язок має місце при j - подібному рості
популяції, при спалахах чисельності видів, при пандеміях тощо.
Негативний зв’язок спостерігається на кожному кроці – збільшення щільності популяції вище
певного рівня призводить, як правило, до зниження темпів росту особин і темпу росту популяції в цілому.
Так, при S - подібному рості популяції 3- тя і 4- та фази росту є взірцями негативного зворотнього зв’язку;
він лежить також в основі регуляції ферментативної активності, різноманітних фізіологічних процесів
тощо. Так, активний розвиток фітопланк-тону призводить до зменшення проникненості світла, в результаті
чого сповільнюється подальший його розвиток. Внесення в грунт азотних добрив значною мірою інгібує
активність азотфіксаторів. Часто неврахування механізмів саморегуляції спотворює результати
екологічних досліджень. Так, при визначенні продукції за склянковим методом, відразу після того, як вода
опиняється в замкненому просторі, запускаються механізми регуляції фотосинтетичної активності:
концентрація СО
2
зменшується, як і концентрація всіх біогенів, що використовуються в процесі
фотосинтезу, що неминуче знижує активність фотосинтетичної діяльності рослин; з іншого боку, зростання
концентрації кисню в свою чергу, інгібує фотосинтез. Окрім цього, як показано експериментами з
радіоактивними мітками, навколо фітопланктону протягом кількох хвилин чи навіть секунд утворюється
простір, позбавлений біогенів, що відразу блокує фотосинтез: в природніх умовах це компенсується
турбулентніс-тю водних мас, переміщенням фітопланктону, потоком метаболітів від зоопланктерів тощо.
Таким чином, чим вищий рівень фотосинтезу, тим істотніше спотворюються результати його вимірів за
методом світлих і чорних склянок.
Згідно закону Ле-Шательє-Брауна будь-яке відхилення параметрів системи від рівноважного їх
стану запускає механізми, що стабілізують стан системи.
На популяційному рівні вкрай важливий механізм регуляції розмірної структури грунтується на
виділенні гідробіонтами видоспецифічних екзометаболітів – речовин, що істотно впливають на темп росту
представників свого виду і майже на впливають на особин іншого виду. Це явище детально досліджене на
молоді риб, личинках амфібій тощо. В усіх випадках при певній концентрації організмів спостерігається
максимальний темп росту, причому значно вищий, ніж при меншій концентрації організмів (типовий
позитивний зворотній зв’язок !) Подальше зростання щільності особин призводить до згначної
диференціації в розмірах та темпові росту: група особин, що виділяються крупними розмірами, має значно
вищий темп росту, ніж більшість особин. Причому
, якщо цих дебелих представників вилучити, то іх місце
тут же займуть інші особини з числа дрібних. Але якщо дрібні і крупні просторово розмежовані (сидять в
різних акваріумах, але вода циркулює між ними, то різниця в рості та його темпах зберігається) – тут ми
маємо, з одного боку, негативний звороній зв’язок (бо в цілому темп росту значно сповільнюється), а з
іншого – «спробу» популяції шляхом дифереціації свого розмірного складу уникнути гострої конкуренції за
трофічні ресурси (аж до канібалізму з боку крупних особин).
62
Досить цікавими є механізми регуляції метаболічних процесів у пойкілотермних тварин.
Вважається, що “холоднокровні” тварини мають таку температуру тіла, як температура зовнішнього
середовища. Частково це так, хоч добре відомо, що у комах в польоті температура тіла значно вища від
температури оточуючого повітря.
Окрім того, багато які пойкілотермні істоти в різний час доби в залежності від свого фізіологічного
стану обирають певні ділянки з тією чи іншою температурою. Так, рептилії полюбляють погрітися на
сонечку. Але значно менше відомо, що риби також істотно змінюють рівень свого метаболізму,
перебуваючи в певних горизонтах води з різною температурою.
Так, молодь (подкаменщика) Cottus extensus першого року життя з озера Бір (США) живиться
виключно в придонних шарах, де температура води складає 4-5
0
С остракодами і придонними веслоногими
ракоподібними (циклопами) Але з настанням сутінок молодь переміщується в шари води з температурою
13-16
0
С, де і тримається до світанку. Перебуваючи в шарах води з температурою на 9
0
С вищою, ніж в
місцях живлення, молодь таким чином підвищую рівень свого метаболізму ( а відповідно живлення і росту)
втричі (Wurtsbaugh, Neverman, 1988).
Прагнення до більш теплих ділянок після споживання їжі притаманне багатьом рептиліям,
амфібіям і рибам, що сприяє активації процесів травлення і пришвидшує метаболічні процеси в цілому. З
іншого боку, для риб показано значення переміщення в більш холодні горизонти води для більш
економного використання наявних кормових ресурсів (шляхом зниження рівня метаболізму в умовах
більш низьких температур).
Дослідження термопреферендумів у цьогорічок ляща (Базаров, Голованов,2000) показало, що
рівень кінцевого термопреферендума молоді ляща при голодуванні знижується на 13-16
0
С у порівнянні з
контролем (у ситих особин) і складає 13-15
0
С. Внесення корму, не залежно від сезону року, призводить до
вибору молоддю ляща літніх температур (26
0
С і вище), які, вочевидь, є оптимальтними для
життєдіяльності і відповідають їх еколого-фізіологічному статусу. При цьому слід відзначити, що реакції
вибору зони оптимуму (чи кінцевого термопреферендуму) проявляється однаковою мірою і у ситих особин
і на початкових стадіях голодування. Лише після 7-9 добового голодування виявляється тенденція до
вибору голодуючими особинами більш холодних ділянок тепературного градієнта. Фізіологічний стан риб
(зокрема, високий чи низький рівень нагодованості..накормленности істотно впливає на характер і
динаміку реакції. Якщо на початку експеримента як ситі, так і голоні риби (коефіціент вгодованості за
Фультоном 1,56) обирали зони кінцевого термопреферендуму на рівні 29-30
0
С, то після голодування
протягом 43 дібііі відновленні годування молоді (коефіціент вгодованості за Фультоном 1,15) відповідний
рівень був на 4
0
С нижчим. Інтервал же обираємих температур у ситих особин був вужчим, а у голодних
розширювався в 2-3 рази, що можна тлумачити як пошук їжі навіть в несприятливих температурних
умовах.
На екосистемному рівні регуляція знаходить свій прояв і в екологічній сукцесії – і авто- і
гетеротрофна сукцесії йдуть у напрямку досягнення термодинамічної рівноваги системи, коли вся
доступна системі енергія (енергія, асимільована системою) йде на підтримання її впорядкованості. Тобто
величина чистої продукції системи прагне до нуля, а вся енергія йде на підтримання негентропії.
Це знаходить свій прояв і у взаємозв’язку речовинно-енергетичних і інформаційних процесів в
екосистемі. Стабільність системи забезпечується їх збалансованістю і узгодженістю.
Кількісною характеристикою регуляторних можливостей системи є порівняння відносних змін
екологічного фактору і параметрів системи. Так, оцінюючи можливість осморегуляторних механізмів, ми
порівнюємо зміну солоності (у скільки разів ) і концентрації солей в тканинних рідинах організму. На
екосистемному рівні можна порівнювати рівень освітленості й рівень фотосинтезу, ступінь забруднення
середовища (концентрацію забруднюючих речовин) і значення найбільш вагомих параметрів екосистеми –
інформаційних, речовинно-енергетичних.
Азовське море завжди вважалося одним з найбільш продуктивних (в тому числі
рибопродуктивних) у світі. В 1930-1960 рр. Вилови риби в ньому досягали 85 кг/га на рік, головним чином
за рахунок тюльки, судака, ляща і хамси. Максималдьна глибина Азовського моря – 14м, солоність
близько 10,5%
о
. Останніми десятиліттями Азовське море опинилося під могутнім антропогенним впливом,
пов’язаним з зарегулюванням стоку Дону і Кубані, що знайшло своє відображення на водному режимі
моря, зростання об’єму стічних вод промислового і сільськогосподарського походження, інтенсифікація
судоходства та іншими, що підірвало запаси бентосу і риб-бентофагів. Цьому сприяли і
63
геологорозвідувальні роботи по пошуку нафти та газу, звалювання в море забруднених грунтів з акваторій
портів і судноходних каналів (дампінг) та ін.
В період інтенсивної хімізації сільського господарства на півдні Росії і в Україні (друга половина
60-х-80-ті роки), зі стоком Дону і Кубані зріс притік добрив і пестицидів в море. Внаслідок цього відбулася
евтрофікація і обумовлене нею “цвітіння” води, а також накопичення залишків пестицидів в трофічних
ланцюгах і акумуляція їх в тілі риб, особливо хижаків. Тяжко вдарила по рибним запасам розорювання
(пойменніх лугов) – нерестовищ риб і засмоктування великої кількості мальків в різноманітні водозабірні
споруди. І все жє таки навіть за цих несприятливих умов, екосистема Азовського моря виявилася досить
стійкою, а загальний вилов риби досягав 250 тис. Т/рік, або в середньому 66 кг/га– головним чином за
рахунок дрібних пелагічних видів– хамси і тюльки. Антропогенний прес на пелагіаль виявився меншим, ніж
на бенталь і планктон Азовського моря продовжував розвиватись досить інтенсивно.
В 1990-ті роки, у зв’язку з економічною кризою та спадом виробництва масштаб антропогенного
впливу на море знизився, токсикогенний стік з полів втратив своє колишнє значення і можна було
сподіватися, що рибопродуктивність Азовського моря почне зростати. Проте відбулося несподіване: в
Азовське море з Чорного, разом з водними масами відбулося вторгнення занесенего з
Атлантичного
океану гребінця мнеміопсис (Mnemiopsis leidyi A.Agassiz). Знайшовши тут чудові харчові ресурси у вигляді
зоопланктону, а також іхтіопланктону. В короткий термін кормова база пелагічних риб виявилася
підірваною, промислові запаси впали до критичного рівня. Рибна промисловість Росії і України в цьому
регіоні понесла величезні, небачені раніше збитки.
В Чорному морі поодинокі екземпляри цього виду гребінця
були помічені в 1982 році, проте вже в
кінці 80-х років його загальна біомаса в морі досягла одного мільярда тон. З Чорного моря мнеміопсис
завдяки системі течій, розселився в сусідні моря – Азовське. Мармурове, Егейське і Середземне.
Слідом за мнеміопсисом в Азовське море вселився гребінець Beroe ovata, який є антагоністом
мнеміопсиса і він вже починає обмежувати його кількість.
Загалом підраховані збитки, нанесені мнеміопсисом рибному промислу – 43 млн долларів США
щорічно. Обгрунтовано метод регуляції щільності мнеміопсиса шляхом вселення його вида-антагоніста –
Beroe ovata.
11.3. Іформаційна структура екосистем
Крім потоків енергії і колообігів речовин, екосистемам притаманні розвинені інформаційні мережі,
які включають потоки фізичних і хімічних сигналів, що забезпечують взаємозв’язки елементів системи між
собою і функціювання її як єдиного цілого. Саме тому є всі підстави вважати, що екосистеми мають
кібернетичну природу (від грецького – kibernētikē – мистецтво управління). Проте слід підкреслити, що, на
відміну від створених людиною кібернетичних пристроїв, її управляючі функції зосереджені всередині неї
дифузно (а не направлені назовні і спеціалізовані).
Надлишковість (коли яка-небудь функція може виконуватися кількома компонентами) підвищує
стабільність системи.
Література
Бигон М., Харпер Дж., Таунсенд К. Экология. Особи, популяции, сообщества: В 2-х т. Пер. с англ. – М.: Мир, 1989. Т.1.– 667 с,
Т.2 – 477 с.
Гиляров А.М. Популяционная экология. – М.: МГУ, 1990. – 191 с.
Дажо Р. Основы экологии. – М.: Прогресс, 1975. – 414 с.
Джефферс Дж. Введение в системный анализ: применение в экологии.–М.: Мир, 1981.– 252 с.
Джиллер
Структура сообществ и экологическая ниша.-М.: Мир, 1988.–184 с.
Константинов А.С. Общая гидробиология. – М.: Высш. шк., 1986.– 472 с.
Кучерявий Екологія. – Львів: Світ, 2000.– 500 с.
Лархер В. Экология растений. – М.: Мир, 1978. –185 с.
Одум Ю. Основы экологии. – М.: Мир, 1975.–742 с.
Одум Ю. Экология. – М.: Мир, 1986. – Т.1-2.
Пианка Э. Эволюционная экология.– М.: Мир, 1981.– 400 с.
Реймерс
Н.Ф. Экология (теории, законы, правила, принципы и гипотезы). – М.: Журнал «Россия Молодая», 1994. – 367 с.
Уиттекер Р. Сообщества и экосистемы.– М.: Прогресс, 1980. – 327 с.
Федоров В.Д., Гильманов Т.Г. Экология. – М.: МГУ, 1980. – 464 с.
Шмидт-Ниельсен К. Физиология животных. Приспособления и среда.– М.: Мир, 1982. – Т. 1.– 416 с.– Т.2.– 384 с.
Яблоков А.В. Популяционная биология
.– М.: Высшая школа, 1987.– 303 с.
64
Вступ до лабораторного практикуму з загальної екології
МЕТОДОЛОГІЧНІ ОСОБЛИВОСТІ СУЧАСНОЇ ЕКОЛОГІЇ
Екологія, як комплексна наука використовує досить широкий арсенал методів, які на перший погляд
властиві іншим фундаментальним наукам. Завдяки широкому арсеналу методів екології відбувається
тісний зв’язок екології з іншими науками.
Згідно Ю.А. Злобіна (1998), методи екології можна розділити на три основні групи:
Методи збору інформації. Класичні методи дослідження стану екологічних об’єктів (включають у
себе всі методи, які застосовують природничі науки), що спрямовані на накопичення фактичного
матеріалу про складові компоненти досліджуваної ділянки екосистеми. Біосфери.
Методи обробки отриманої інформації. Дана група методів спрямована на узагальнення
отриманої інформації шляхом систематизації певних параметрів складових компонентів досліджуваної
ділянки екосистеми. Сучасна обчислювальна техніка дає можливість обробляти велику кількість
фактичного матеріалу, що, в свою чергу, робить великий фактичний матеріал більш доступним для
узагальнення. Слід відмітити, що при певних екологічних дослідженнях статистична обробка є необхідною
умовою достовірності отриманих результатів.
Методи інтерпретації отриманих результатів. Методи моделювання. Важким етапом будь-
яких екологічних досліджень є здатність аналізу отриманих результатів, побудова певної моделі стану
екосистеми. Такий підхід дає змогу прогнозувати зміни, які можуть відбуватися на досліджуваній ділянці
під впливом певних екологічних факторів або під впливом діяльності людини. На основі абстрагування
результатів досліджень можна робити словесні описи екосистем (вербальні моделі), побудувати схеми
взаємозв’язків компонентів (графічні моделі), робити спробу опису екосистеми за допомогою
математичних формул (математичні моделі). Безапеляційне вживання методів моделювання неможливе
через непередбачуваність процесів, які відбуваються в екосистемах, залежить від “великих” та “малих”
циклів. Як правило планетарної природи. Тільки після накопичення достатньої кількості інформації про
закономірності існування Всесвіту, побудовані моделі будуть найбільш наближені до реального стану
екосистем. Введення змінних величин не дає можливість вирішувати дані проблеми, математичні
формули стають більш ускладненими. Спрощення та вилучення змінних компонентів веде до втрати
достовірності самої моделі.
Методологічною засадою сучасної екології є комплексне використання натурних спостережень,
вимірювань і досліджень, експериментальних лабораторних досліджень, екологічного картування і
моделювання. Більшості екологічних досліджень притаманний системний підхід. У сучасних екологічних
дослідженнях широко використовують методи інших наук – хімії, фізики, геології, біології, математики
тощо. Ці методи можна об’єднати в кілька груп:
1. Методи реєстрації та оцінки якості довкілля, насамперед різни типи екологічного моніторингу,
зокрема біомоніторинг і біоіндикаця, дистанційний і аерокосмічний моніторинг.
2. Методи кількісного обліку організмів і методи оцінки біомаси та продуктивності рослин і тварин.
3. Вивчення особливостей впливу різних екологічних чинників на життєдіяльність організмів (як
складні й тривалі спостереження в природі, так і, частіше, експерименти в лабораторних умовах –
токсикологічні, біохімічні, біофізичні та ін.).
4. Методи вивчення взаємозв’язків між організмами в багатовидових угрупованнях.
5. Методи математичного моделювання екологічних явищ і процесів, а також екосистем (останнє –
набагато складніше), імітаційне моделювання; моделювання від локальних до регіональних і глобальних
екологічних процесів і ситуацій.
6. Створення геоінформаційних систем і технологій для розв’язання екологічних питань різних
масштабів і в різних сферах діяльності.
7. Комплексний еколого-економічний аналіз стану різних об’єктів, територій, галузей виробництва.
8. Геоекологічні методи дослідження, геоекологічний моніторинг з метою зменшення негативного
впливу забруднювачів на довкілля.
9. Технологічні методи екологізації різних виробництв з метою зменшення їх негативного впливу на
довкілля.
10. Медико-екологічні методи вивчення впливу різних чинників на здоров’я людей.
65
11. Методи екологічного контролю стану довкілля: екологічна експертиза, екологічний аудит,
екологічна паспортизація.
У рамках лабораторного практикуму переважно відбувається ознайомлення з класичними
методами екологічних досліджень.
Практична робота №1
Тема: Визначення параметрів навколишнього середовища
Мета: Набути практичних навичок визначення фізичних параметрів навколишнього середовища
Завдання: ознайомитись із приладами для вимірювань значень деяких факторів навколишнього
середовища, замалювати прилади, провести виміри температури, вологості, освітленості, швидкості вітру,
тиску.
Матеріали: термометри (ртутний, спиртовий), психрометр Асмана, люксметр, анемометр, секундомір,
барометр.
Теоретичні відомості
Живі організми знаходяться під постійним впливом абіотичних факторів, що
створюють фізико-хімічні умови
навколишнього середовища. Для вимірювання параметрів навколишнього середовища використовують різноманітні
фізичні прилади.
Температура. Для вимірювання температури повітряного, ґрунтового, водного середовища використовують
термометри. Принцип дії термометра базується на властивості речовини змінювати об’єм в залежності від
температури. В залежності від вимог до проведення досліджень, використовують спиртовий, ртутний або
біметалічний термометри. Для вимірювання температури ґрунту використовують термометр-щуп, що дозволяє
робити заміри температури на різній глибині від поверхні. Одиниці виміру – градуси за Цельсієм (°С).
Для вимірювання вологості повітря застосовують аспіраційний психрометр Асмана. Психрометр
складається із двох термометрів – сухого і змоченого. Датчик одного із термометрів обмотаний батистом, який
змочений водою.
Психрометр обладнаний вентилятором, при ввімкненні якого термометри обдуваються повітрям.
Внаслідок випаровування води змочений термометр показує нижчу температуру, ніж сухий. Чим менша вологість,
тим більша інтенсивність випаровування і ти нижчими є показники змоченого термометра. Різниця температур між
змоченим і сухим термометром буде характеризувати вологість повітря. Одиниці виміру – відсоткове
співвідношення у повітрі (%).
Для
визначення освітленості поверхні використовується фізичний прилад – люксметр, принцип дії якого
базується на фізичних властивостях фотоелементу. В залежності від освітленості стрілка люксметра показує різні
величини, що і буде характеризувати освітленість. Одиниці виміру – люкс (лк).
Швидкість вітру вимірюють за допомогою фізичного приладу – анемометра. Принцип дії анемометра
базується на основі обертання “крила” приладу
за певний період часу. Після встановлення анемометра у
повітряному потоці і фіксації даних усіх шкал через однакові інтервали часу (10-15 сек) одночасно включають
механізм приладу і секундомір. Визначення проводять протягом 1-2 хв. Після завершення вимірів анемометр та
секундомір зупиняють і фіксують дані обох приладів. Швидкість вітру визначають за графіком, що додається до
анемометра
. Для цього по вертикальній вісі графіку відшуковують значення шкали лічильника анемометра за одну
секунду. З цієї точки проводиться горизонтальна лінія до перетинання з графіком, потім від точки перетину до
горизонтальної вісі. Показники на горизонтальній вісі і будуть характеризувати швидкість вітру в м/сек.
Приблизну оцінку сили вітру можна провести і
за видимими проявами (шкала Бофорта).
Для визначення тиску повітря застосовують барометр. Одиниці виміру – мм ртутного стовпа або кПа.
Хід роботи
Після ознайомлення із принципами роботи приладів, студенти визначають фізичні параметри навколишнього
середовища. Для цього студенти розділяються на групи із 2-3 (5) чоловік, кожна з яких проводить дослідження
окремих параметрів навколишнього середовища. Результати зводяться у формі таблиць для декількох досліджень.
На основі отриманих результатів роблять висновки.
Питання для самоконтролю
1. У чому
полягає принцип дії термометра?
2. За яким принципом діє психрометр Асмана?
3. Чому температура змоченого термометра нижче від сухого?
4. Який принцип дії люксметра і в яких одиницях вимірюється освітленість?
5. У чому полягає принцип дії анемометра і в яких одиницях вимірюється швидкість вітру?
6. На основі якої властивості волосу
базується принцип дії барометра?
66
Практична робота №2
Тема: Визначення параметрів навколишнього середовища протягом
довгого періоду часу
Мета: Набути практичних навичок у організації та проведенні вимірів фізичних параметрів
навколишнього середовища за допомогою автоматичних приладів реєстрації
Завдання: ознайомитись із приладами автоматичного вимірювання значень деяких факторів
навколишнього середовища, принципом їх роботи, провести виміри температури, вологості, тиску протягом
тривалого періоду часу.
Матеріали: термограф, гігрограф, барограф, термометр, психрометр Асмана, барометр, чорнило,
міліметровий
папір.
Теоретичні відомості
Для тривалих досліджень фізичних параметрів навколишнього середовища застосовують різноманітні
прилади автоматичної реєстрації тих чи інших показників. Автоматичний прилад для реєстрації складається із
датчика, механізму передачі даних, барабану, механізму обертання барабану. В залежності від того, які фізичні
параметри необхідно отримати, застосовують такі прилади: термограф (для автоматичної фіксації температури),
барограф (для автоматичної реєстрації змін тиску), гігрограф (для автоматичної реєстрації зміни вологості).
Відмінності у побудові цих приладів полягають тільки у механізмах датчиків. У гігрографа датчик вологості –
пучок (35-40 шт.) знежиреного людського волосу, захищеного від пошкодження спеціальним пристроєм. Датчик
термографа – біметалічна пластинка, захищена від пошкоджень спеціальним пристроєм. Принцип роботи
барографа базується на роботі
комплекту анероїдних мембран і компенсаторної біметалічної пластини.
Принцип дії автоматичних пристроїв широко використовується і при інших екологічних дослідженнях.
Зокрема, вдалим є використання цих пристроїв для вивчення частоти відвідування гнізд птахами під час
вигодовування пташенят, вивчення активності дрібних ссавців тощо. Простота у виготовленні, надійність роботи та
досить висока точність автоматичних приладів
забезпечує їх широке використання.
Хід роботи
Після ознайомлення із принципами роботи пристроїв автоматичної реєстрації, студенти за допомогою
термометру, психрометра Асмана, барометра калібрують шкалу на спеціальному міліметровому папері. Після
калібрування і визначення точки початку досліду міліметровий папір заправляється на барабан, чорнилом
наповнюється перо, запускається годинниковий механізм обертання барабану. На наступне заняття,
після зупинки
годинникового механізму, зніміть міліметровий папір і проведіть розшифровку отриманих результатів.
Отримані результати занотуйте у вигляді графіку. Зробіть висновок.
Питання для самоконтролю
1. Для чого використовують автоматичні пристрої реєстрації?
2. В чому полягає принцип роботи автоматичних пристроїв?
3. Які відмінності у роботі термографа, барографа, гігрографа?
4. Яку інформацію можна
отримати за допомогою автоматичних приладів?
Практична робота №3
Тема: Дослідження ґрунтів як складового елементу біотопу
Мета: Ознайомитись із основними методами вивчення ґрунтів
Завдання: дослідити ґрунтові характеристики двох ділянок, які відрізняються біотопічно.
Матеріали: лопата, лінійка, поліетиленові пакети, ваги.
Теоретичні відомості
Едафічні фактори – це ґрунтові умови, що впливають на життя і поширення живих організмів. Ґрунт має
трифазну структуру, оскільки складається із твердих частинок, води і повітря.
Потужність
ґрунту – це товщина в сантиметрах. До генетичних горизонтів відносяться наступні: лісова
підстилка, перегнійно-акумулятивний, або гумусовий, підзолистий горизонт, материнська порода.
Гумус обумовлює появу чорного або сірого кольору, окис заліза – червоне, жовте, іржасте забарвлення.
Кварц і шпат білий, заболочені ґрунти зеленуваті.
За механічним складом розрізняють глинисті ґрунти, суглинки, піщані, супіщані. Глинисті ґрунти
розтираються
руками важко, супіщані – легко. Суглинки при розтиранні утворюють тонкий порошок, в якому відчуваються піщинки.
За структурою ґрунти можна розділити на горіхуваті та пластинчасті. За щільністю – на рихлі та щільні.

67
Хід роботи
Перш ніж розпочати проведення розрізу, слід оглянути місцевість. Розріз не повинен закладатися поблизу
доріг, канав та у нетипових для мікрорельєфу ділянках. На вибраній ділянці прокопайте ґрунтовий розріз так, щоб
одна стінка його була рівною, а протилежна була у вигляді сходинок. Бічні також мають бути вертикальними. При
прокопуванні ґрунт
слід викидувати в сторони, щоб не забруднити лицевої стінки, за якою і буде робитися опис
горизонтів. Після прокопування визначте потужність горизонтів у сантиметрах, використовуючи для цього лінійку.
Візуально оцініть і опишіть колір ґрунту. Розітріть ґрунт і дайте характеристику його механічному складу. На зрізі
дайте оцінку структури та щільності ґрунту. Результати оформіть
у вигляді таблиці.
Таблиця
Генетичний
горизонт
Потужність (см) Колір Механічний склад Структура Щільність
Питання для самоконтролю
1. До яких екологічних факторів відносяться ґрунтові умови?
2. Що таке ґрунт, які його складові компоненти?
3. Як відрізнити генетичні горизонти на розрізі?
4. Що таке гумус, чим він утворений?
ВИЗНАЧЕННЯ ГРАНУЛОМЕТРИЧНОГО СКЛАДУ ТА ВОДОПІДЙОМНОЇ ЗДАТНОСТІ ҐРУНТІВ
Мета: набуття практичних навичок визначення гранулометричного складу ґрунту в польових умовах
та визначення висоти капілярного підйому води в ґрунтах різних типів.
Матеріал та обладнання: фарфорові чашки, скляні трубки висотою 30 см та діаметром 1,5 – 2 см,
марля, ванночки або кювети, фільтрувальний папір, дерев’яні дощечки, середні проби грунтів різних типів
(дерново-слабопідзолистий піщаний,
лучний на алювіальних відкладах, темно-сірий опідзолений на лесовидних
суглинках, типовий малогумусний чорнозем на лесовидних породах).
Пояснення до завдання
Гранулометричний (механічний) склад ґрунтів має велике агрономічне значення. З ним безпосередньо
пов’язана фільтраційна та водозатримувальна здатність, швидкість просихання, тепловий режим, накопичувальна
здатність ґрунту тощо. Найбільш поширений метод визначення гранулометричного складу
ґрунту в польових умовах
є метод джгута (проба на скачування).
Згідно з цим методом склад ґрунту визначають за наступними показниками:
- піщаний – не скочується в шнур або кульку.
- супіщаний – при розкачуванні сплющується, утворюючи неміцні кульки.
- легкосуглинковий - при розкачуванні утворює неміцний шнур, який розпадається на окремі джгутики.
- середньосуглинковий – при розкачуванні утворює хороший шнур, який розламуються при згортанні в
кільце.
- важкосуглинковий – при розкачуванні утворює хороший шнур, який при згортанні в кільце дає тріщини.
- глинистий – при розкочуванні утворює міцний шнур, який легко згортається в кільце, не утворюючи тріщин
Велике значення для властивостей ґрунту має вода,
яка знаходиться в ньому у формах: хімічно зв’язаній,
пароподібній, плівковій (фізично зв’язаній), капілярній та гравітаційній.
Водопідйомна здатність – здатність ґрунту викликати капілярний підйом води. Вона залежить від
механічного складу та агрегованості ґрунту. Максимальна висота підйому ґрунтової вологи по капілярах щодо
піщаних ґрунтів становить 0,5 – 0,7 м, суглинкових - 3-5 м.
Хід роботи
1.
Визначення гранулометричного складу ґрунту
Взяти почергово невеликі частини проб ґрунтів, висипати до фарфорової чашки та намочити водою до
тістоподібного стану. З отриманого «тіста» скатати спочатку кульку діаметром 1,5-2 см, який потім розкатати в шнур
та згорнути в кільце діаметром близько 2 см (на долоні або навколо пальця). За запропонованими вище
показниками, визначити гранулометричний склад ґрунту.
2.
Визначення водопідйомної здатності ґрунтів
Скляні трубки обв’язати з нижнього кінця марлею, а через верхній вільний кінець насипати ґрунт з різних проб до
висоти приблизно 20 см. Закріпити трубки вертикально, поставити на дощечку з фільтрувальним папером, кінці
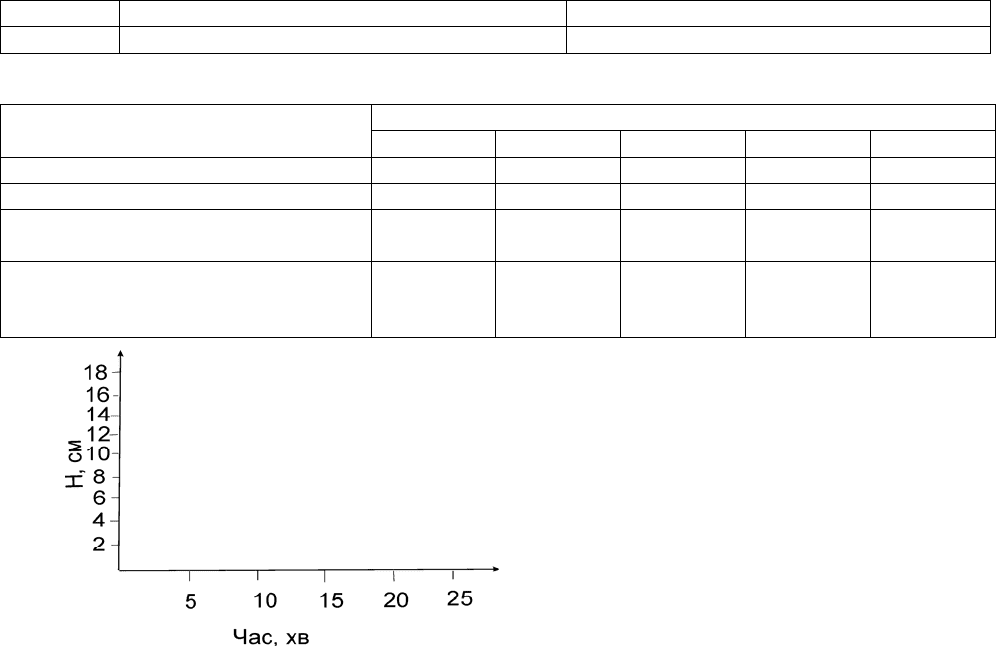
якого опустити у ванночку з водою. Зробити виміри висоти підйому вологи в трубочках з ґрунтом через 5 хв, 10 хв,
15 хв, 20 хв, 25 хв.
68
Фіксація результатів
1. Занести результати завдання 1 до таблиці. Зробити висновки щодо механічного складу ґрунтів різних
типів.
2. Записати результати завдання 2 до таблиці. За цими даними побудувати графіки (див. рис.). Зробити
висновки щодо швидкості підйому капілярної води в різних ґрунтах.
Гранулометричний склад ґрунту
№ зразка Результати скачування Гранулометричний склад
Водопідйомна здатність ґрунтів
Висота підйому вологи в ґрунтах, см
Ґрунт
5 хв 10 хв 15 хв 20 хв 25 хв
Дерново-слабопідзолистий піщаний
Лучний ґрунт на алювіальних відкладах
Темно-сірий опідзолений ґрунт на
лесовидних суглинках
Типовий
малогумусний чорнозем на лесових
породах
Рис. Приклад побудови графіка висоти капілярного підйому води в ґрунті.
Практична робота № 4
Тема: Методи виміру фонової радіоактивності, гамма та бета випромінювання
Завдання: використовуючи дозиметр-радіометр, провести заміри фонової радіації.
Матеріали: дозиметр-радіометр "Припять"
Теоретична частина
Усі види іонізуючого випромінювання підрозділяють на дві групи: електромагнітне випромінювання (до яких
відноситься рентгенівське і гама-випромінювання) та корпускулярне випромінювання (випромінювання різного роду
ядерних частинок).
У природі є багато джерел природного іонізуючого випромінювання. Перш за все
– це ізотопи багатьох
елементів, що знаходяться у складі гірських порід та мінералів. Головним є калій–40 та вуглець–14.
Несприятливість біологічної дії радіоактивних речовин пов’язана не тільки з їхньою разовою дією, а із здатністю
акумулюватися в організмі. Стронцій–90 накопичується в кістках, йод–131 – в щитовидній залозі, цезій–137
включається в активний метаболізм, витісняючи
азот.
Космічне випромінювання також є джерелом радіаційного фону. Космічні промені дають трохи менше
половини зовнішнього опромінення, яке утримує населення від природних джерел радіації. Космічні промені в
основному приходять до нас із глибин Всесвіту, але деяка їх частина утворюється на Сонці під час сонячних
спалахів. Космічні промені можуть досягати поверхні Землі або
взаємодіяти з її атмосферою, породжуючи вторинне
випромінювання та сприяючи утворенню різноманітних радіонуклеїдів.
69
Небезпека, що пов’язана з атомною енергетикою та атомним озброєнням, була яскраво продемонстрована
аварією на Чорнобильській АЕС у 1986 р. У результаті аварії в навколишнє середовище потрапили радіоактивні
ізотопи свинцю-239, цезію-137, стронцію-90, плутонію-240.
Загальний фон радіоактивного випромінювання на території України складає – 0,005-0,06 мР/год.
Хід роботи
Включіть прилад, для чого перемикач живлення
переведіть в положення "Вкл". На цифровому табло повинні
індукуватись:
Включення приладу повинно супроводжуватись коротким звуковим сигналом. При природньому фоновому
випроміненні прилад повинен подавати 1-6 звукових сигналів у хвилину. Із збільшенням дози гамма
випромінювання пропорційно виростає частота звукових сигналів.
Для випромінювання потужності експозиційної дози гамма випромінювання зробить наступні кроки:
1) перевірте, чи
закрита задня кришка приладу, при необхідності закрийте її;
2) переведіть перемикач режиму роботи в положення "МД" (крайнє ліве положення);
3) включіть прилад перемикачем живлення і натисніть кнопку "пуск/стоп". При цьому на цифровому
табло повинні з’явитись крапки після кожного розряду і розпочатись відлік імпульсів.
4) через 20-25 секунд вимірювання закінчиться
, що супроводжується звуковим сигналом, а на цифровому
табло зафіксується число з однією комою. Даний показник приладу буде відповідати потужності експозиційної дози
гамма-випромінювання, виміряної у мР/год. Для виконання повторного заміру достатньо ще раз натиснути на кнопку
"пуск". Після закінцення випромінювання вимкніть прилад. Для отримання більш точних даних проведіть
вимірювання
3-5 разів, розрахуйте середнє арифметичне.
Для вимірювання щільності бета-випромінювання, зробіть наступні кроки:
1) перевірте, чи закрита задня кришка приладу, при необхідності закрийте її;
2) переведіть перемикач режиму роботи в положення "МД" (крайнє ліве положення);
3) піднесіть прилад поверхнею задньої кришки до досліджуваної поверхні на відстань 0,5-1,0
см і натисніть кнопку "пуск
";
4) виконайте замір і занотуйте показник індикатора;
5) відкрийте задню кришку приладу;
6) виконайте замір при відкритій задній кришці, запишіть показник індикатора;
7) закрийте задню кришку, вимкніть прилад;
8) за формулою: q = K
s
(Nγ + β + Nγ) розрахуйте величину щільності потоку бета-випромінювання.
Одиниці виміру - част./см
2
хв.
На основі проведених вимірів зробить висновок.
Питання для самоконтролю
1. Що таке альфа-випромінювання?
2. Що таке бета-випромінювання?
3. Чим шкідливе радіоактивне випромінювання?
4. З чого складається фонове радіоактивн випромінювання?
0.000 0000
або
0.0.0.0.
70
ПИТАННЯ ПИСЬМОВОГО ТЕОРЕТИЧНОГО ТУРУ ОЛІМПІАДИ З ЗАГАЛЬНОЇ ЕКОЛОГІЇ ДОННУ
1. Екологія як наука: класичне і сучасне тлумачення, схеми внутрішньої будови.
2. Концептуальні основи неоекології: визначення, призначення, класифікація.
3. Біосфера: склад, межі та функціонування.
4. Екологічні фактори: визначення, класифікація.
5. Екосистема: будова та основні процеси функціонування.
6. Поняття обмежуючого фактору та
екологічної толерантності. Закон Шелфорда.
7. Чим спричинена поява “озонових дір”? заходи з подолання цього катастрофічного
явища.
8. Шумове забруднення довкілля: визначення, рівні, наслідки, застережні заходи.
9. Екологічні “закони” Б. Коммонера. Сформулювати та проілюструвати прикладами.
10. Контроль і управління водними ресурсами: основні поняття, показники. Вимоги до
якості води.
11. Екологічна безпека: визначення
, фактори та міри з її забезпечення у регіонах України.
12. Рух ксенобіотиків за трофічними ланцюгами та їх вплив на стан здоров’я людини.
13. Екологія геоенергоаномальних зон.
14. Дати визначення і навести відомі вам класифікації природних ресурсів.
15. Дати розгорнуту характеристику одного з основних засобів очищення стічних вод.
16. Дати
розгорнуту характеристику одного з основних засобів очищення пило-газових
викидів промислових підприємств.
17. Антропогенна євтрофікація внутрішніх водоймищ: принципи, механізм і наслідки.
18. Класифікація та основні ознаки території і об’єктів природно-заповідного фонду
України.
19. Засоби рекультивації порушених земель.
20. Екологічні і соціальні проблеми гідроелектростанцій, побудованих на рівнинних ріках.
21. Екологічні
проблеми енергетики та шляхи їх розв’язання.
22. Дати характеристику альтернативних джерел енергії в екологічному аспекті.
23. Принципи санітарного зонування промислових підприємств.
24. Дати розгорнуту характеристику засобам переробки твердих відходів.
25. Екологічне нормування. Сутність нормативів ГДВ і ГДС.
26. Дати стислу характеристику екологічного права як механізму управління в системі
природокористування.
27. Захист довкілля від викидів автомобільного транспорту.
28. Дати стислу характеристику об’єктів і засобів екологічної експертизи.
29. Концепція екологічної освіти і виховання в Україні.
30. Роль і форми участі громадськості щодо вирішення екологічних проблем в Україні.