Албертс Б., Брей Д. и др. Молекулярная биология клетки. Том 1
Подождите немного. Документ загружается.


281
дезаминирования цитозина в урацил, частота которых, согласно оценкам, достигает 100 на один геном в сутки (рис. 5-31). Содержащиеся в ДНК
основания изменяются также под влиянием реакционноснособных метаболитов, нарушающих их нормальное спаривание, а также под действием
ультрафиолетовой радиации Солнца, которая может вызвать образование ковалентной связи между двумя соседними остатками пиримидиновых
оснований в ДНК (образование димеров тимина; рис. 5-32). Все перечисленное выше - это лишь небольшая часть тех многочисленных изменений,
которые происходят спонтанно в нашей ДНК. Большинство из них должно было бы привести либо к выпадению одной или нескольких пар
оснований в дочерней цепи ДНК после цикла репликации, либо к замене пары оснований (например, каждое дезаминирование С → U должно было
бы вызвать в конце концов замену пары C-G на пару Т-А, поскольку U ведет себя сходно с Т и образует комплементарную пару с А). Как мы уже
знаем, такие изменения, если бы они происходили достаточно часто, имели бы роковые последствия для живых организмов.
5-14
5.2.7. Стабильность генов обеспечивается репарацией ДНК [21]
Хотя в ДНК любой клетки человека под влиянием тепловой энергии происходят ежедневно тысячи случайных изменений, за год в
каждой клетке накапливается (если только вообще накапливается) лишь очень небольшое число стабильных изменений нуклеотидной
последовательности ДНК. Мы знаем теперь, что среди множества случайных замен оснований в ДНК лишь одна на тысячу приводит к
возникновению мутации, все же остальные повреждения очень эффективно ликвидируются в процессе репарации ДНК. Все репарационные
механизмы основаны на том, что в клетке имеются две копии генетической информации - по одной в каждой из двух цепей молекулы ДНК. Если
нуклеотидная последовательность одной из цепей случайно оказывается измененной, информация не утрачивается, поскольку вторая ее копия
хранится в нуклеотидной последовательности другой цепи ДНК. Из схемы на рис. 5-33 видно, что основной путь репарации ДНК включает три
этапа.
1. Измененный участок поврежденной цепи ДНК распознается и удаляется при помощи специфических ферментов, носящих название
ДНК-репарирующих нуклеаз; они осуществляют гидролиз фосфодиэфирных связей между поврежденными нуклеотидами и остальной частью
молекулы ДНК, в результате чего в спирали ДНК в этом месте возникает брешь.
2. Другой фермент, ДНК-полимераза, связывается с 3'-концом поврежденной цепи ДНК и заполняет эту брешь путем присоединения
одного нуклеотида за другим, копируя информацию, содержащуюся в «хорошей» (матричной) цепи.
3. В заключение фермент, называемый ДНК-лигазой, «сшивает» ДНК и тем самым завершает восстановление интактной молекулы.
Рис. 5-32. Образование тиминового димера распространенный тип
повреждения ДНК под действием ультрафиолетовых лучей (в
частности, пол действием солнечного света). Подобный димер
способны образовать два любых соседних пиримидиновых основания
(С или Т).
Рис. 5-33. Три этапа репарации ДНК. На первом этапе вырезается
поврежденный участок, на втором и третьем этапах происходит
восстановление исходной нуклеотидной последовательности ДНК.
ДНК-полимераза заполняет брешь, возникшую вследствие удаления
поврежденной части цепи (2-й этап), а ДНК-лигаза сшивает разрыв,
оставшийся в «исправленной» цепи (3-й этап). Сшивание
осуществляется путем восстановления разорванной фосфодиэфирной
связи (см. рис. 5-35).
282
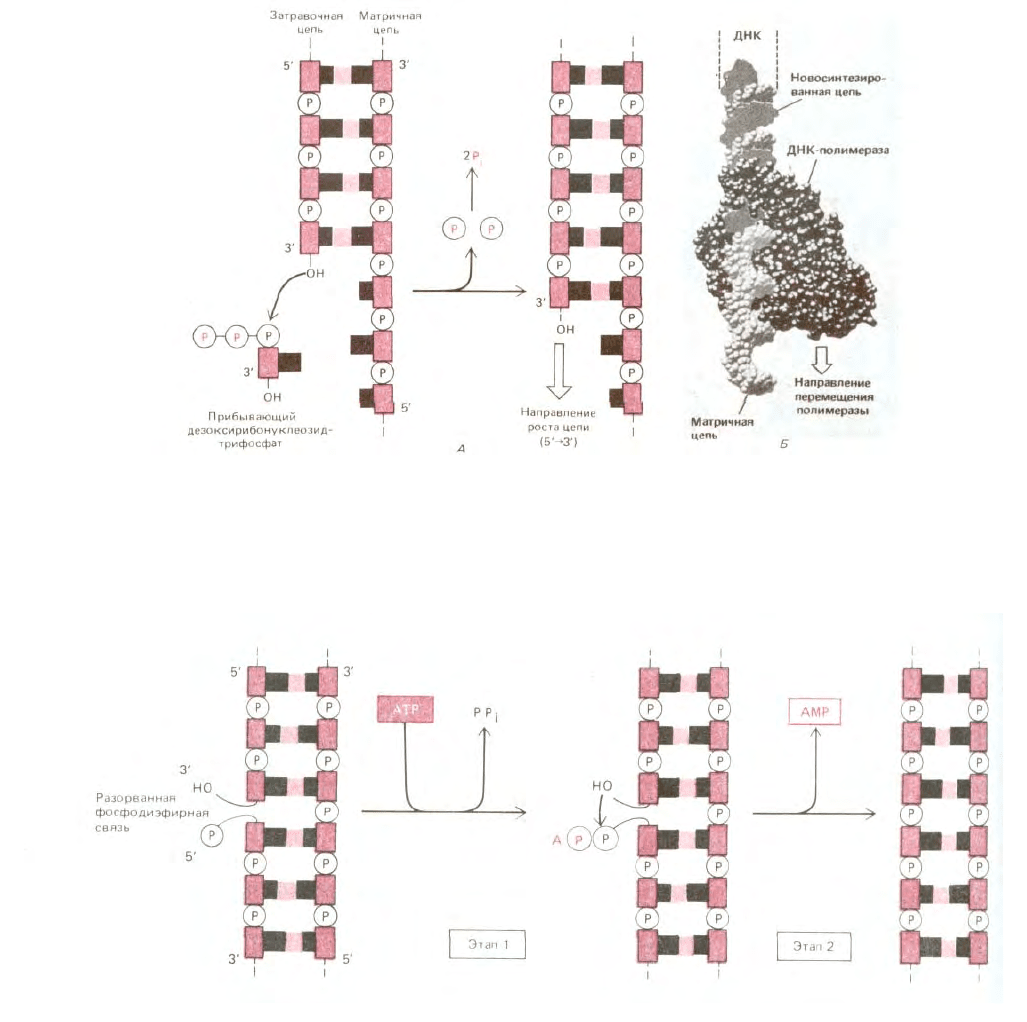
Рис. 5-34. Фермент ДНК-полимераза. А. Реакция, катализируемая ДНК-полимеразой. Этот фермент катализирует поэтапное присоединение
дезоксирибонуклеотидов к 3'-концу полинуклеотидной (затравочной) цепи, спаренной с другой (матричной) полинуклеотидной цепью. Таким
образом, новая цепь ДНК растет в направлении 5' → 3'. Поскольку каждый прибывающий дезоксирибонуклеозидтрифосфат должен спариться с
матричной цепью, для того чтобы его могла узнать ДНК-полимераза, именно матричная цепь определяет, какой из четырех возможных
дезоксирибонуклеотидов (А, С, G или Т) присоединится к 3'-концу синтезируемой цепи. Как и в случае РНК-полимеразы, движущей силой реакции
служит значительное выгодное изменение свободной энергии (см. рис. 5-2). Б. Структура ДНК-полимеразы Е. coli, определенная методом
рентгеноструктурного анализа. ДНК-полимераза изображена здесь в момент участия в синтезе ДНК. (С любезного разрешения Тот Steitz.)

283
← Рис. 5-35. Фермент ДНК-лигаза восстанавливает разорванную фосфодиэфирную связь. Из схемы видно, что ДНК-лигаза использует сначала
молекулу АТР, для того чтобы активировать в точке разрыва 5'-конец поврежденной цепи (1-й этап), и лишь после этого образует новую связь (2-й
этап). Энергетически невыгодная реакция сшивания разрыва осуществляется, таким образом, благодаря сопряжению с энергетически выгодным
процессом гидролиза АТР. У больных синдромом Блума (одно из наследственных заболеваний) обнаружена частичная недостаточность ДНК-
лигазы. В связи с этим у них нарушена репарация повреждений ДНК и как следствие повышена частота заболевания раком.
Ферменты ДНК-полимераза и ДНК-лигаза играют важную роль в метаболизме ДНК: оба этих фермента участвуют не только в
репарации, но и в репликации ДНК. Катализируемые ими реакции иллюстрируют соответственно рис. 5-34 и 5-35.
5-18
5-19
5.2.8. Различные типы повреждений в ДНК распознаются разными ферментами [22]
Способ, каким в процессе репарации осуществляется удаление поврежденного участка, зависит от типа повреждения. Например, при
апуринизации (наиболее часто встречающемся повреждении ДНК) один из остатков дезоксирибозы лишается ранее находившегося при нем
основания (см. рис. 5-31). Фермент АП-эндонуклеаза быстро распознает данный остаток дезоксирибозы и разрывает в этом измененном участке
цепи фосфодиэфирную связь. После этого поврежденный нуклеотид удаляется и правильная последовательность нуклеотидов восстанавливается
при помощи механизма, представленного на рис. 5-33.
Другой, близкий к этому, путь репарации связан с участием особого набора ферментов, называемых ДНК-гликозилазами. Каждый из этих
ферментов узнает какой-либо один определенный тип измененных оснований в ДНК и катализирует гидролитическое отщепление такого
основания. Существует, как полагают, не менее шести типов ферментов, входящих в эту группу. Среди них имеются ферменты, удаляющие
дезаминированный цитозин, дезаминированный аденин, алкилированные основания разных типов, основания с разомкнутым кольцом и основания,
в которых двойная углерод-углеродная связь заменена простой. Общий для всех случаев механизм проиллюстрирован на рис. 5-36 конкретным
примером. Здесь представлено удаление дезаминированного цитозина. Сначала фермент урацил-ДНК-гликозилаза удаляет измененное основание
(урацил). Дезоксирибозу, утратившую бывшее при ней основание, узнает другой фермент-АП-эндонуклеаза. Поскольку это тот самый фермент,
который узнает апуринизированные участки ДНК, восстановление правильной последовательности идет далее тем же путем, который мы уже
описали для случая апуринизации. В итоге U, возникший вследствие случайного дезаминирования, вновь замещается на С. Важность процесса
удаления из ДНК случайно дезаминированных оснований удалось продемонстрировать непосредственно на конкретных примерах. Один из них
касается бактериальных штаммов, у которых вследствие мутации отсутствует фермент урацил-ДНК-гликозилаза. Выяснилось, что у таких мутантов
частота спонтанных замен C-G на Т-А (в норме низкая) возрастает приблизительно в 20 раз.
В клетках имеется особый путь для удаления почти любого типа повреждения в ДНК, затрагивающего очень большой ее участок. Такие
обширные повреждения возникают, например, при ковалентных взаимо-

284
действиях между основаниями ДНК и объемистыми углеводородами, в частности бензпиреном, обладающим канцерогенными свойствами. К ним
же относятся и различные пиримидиновые димеры (Т-Т, Т-С и С-С), возникающие под действием солнечных лучей (см. рис. 5-32). В подобных
случаях крупный мультиферментный комплекс узнает не какое-либо одно специфическое изменение основания, а обширное повреждение двойной
спирали ДНК. Фосфодиэфирные связи поврежденной цепи по обе стороны от повреждения разрываются и измененный участок удаляется весь
целиком. После этого восстановление нормальной последовательности происходит как обычно.
О роли репарационных процессов свидетельствует тот факт, что клетки затрачивают большую часть своих ресурсов на производство
репарационных ферментов. Обширные исследования, проведенные на дрожжах, выявили у них свыше 50 различных генов, кодирующих такие
ферменты. Не менее сложны пути репарации ДНК у человека. Выяснилось, что у больных с пигментной ксеродермой нарушен процесс репарации
обширных повреждений, в котором, как показывает генетический анализ, участвует не менее 7 различных генных продуктов. У таких больных в
клетках накапливаются пиримидиновые димеры, что приводит к тяжелому поражению кожи, включая рак.
5-15
5.2.9. Клетки синтезируют репарирующие ферменты в ответ на повреждение ДНК [23]
В процессе эволюции клетки выработали много различных механизмов, обеспечивающих их выживание в этом мире, полном
всевозможных опасностей. Часто какое-нибудь резкое воздействие среды активирует целый набор именно тех генов, продукты которых способны
защитить клетки от этого воздействия. Всем клеткам присущ, например, такой механизм, как реакция на тепловой шок; ее можно наблюдать в
клетках, подвергшихся действию чрезмерно высоких температур. При этом индуцируется синтез особых «шоковых» белков; часть из них, по-
видимому, помогает стабилизировать и репарировать другие клеточные белки, частично денатурированные тепловым шоком.
Во многих клетках существуют также механизмы, дающие им возможность синтезировать ферменты для репарации ДНК, так сказать, в
аварийных ситуациях, в ответ на серьезные повреждения ДНК. Среди примеров такого рода лучше всего изучен SOS-ответ (SOS-репарация) у Е.
со/г. У этой бактерии любое нарушение репликации ДНК, вызванное ее повреждением, ведет к появлению сигнала (таким сигналом служит, по-
видимому, избыток одноцепочечной ДНК), усиливающего транскрипцию более чем 15 различных генов, многие из которых кодируют белки,
участвующие в репарации ДНК. Сигнал активирует у Е. coli белок (см. разд. 5.4.4), который затем разрушает другой белок - отрицательный
регулятор активности генов (репрессор). Действие этого репрессора заключается в подавлении у Е. coli транскрипции всего набора генов,
участвующих в SOS-ответе. Изучение бактериальных мутантов с различными нарушениями SOS-репарации показало, что новосинтезированные
белки обусловливают два эффекта. Во-первых, их индукция повышает выживаемость клеток: если мутанты, у которых синтез таких ферментов
нарушен, подвергнуть действию тех или иных агентов, вызывающих повреждение ДНК (например, ультрафиолетовых лучей), то процент погибших
клеток окажется необычно высоким. Во-вторых, некоторые из индуцированных белков вызывают временное повышение частоты мутаций,
вследствие чего генетическая изменчивость бактериальной популяции возрастает. Выгода здесь, видимо, заключается в том, что

285
таким путем увеличивается шансы на появление мутантной клетки с повышенной приспособленностью.
Существуют и другие индуцируемые системы репарации ДНК. Известно, например, что одна из них у бактерий активируется
присутствием в ДНК метилированных нуклеотидов. Аналогичная система функционирует в клетках дрожжей. Есть сведения, что и некоторые
высшие эукариотические клетки адаптируются к повреждениям ДНК аналогичным путем.
5.2.10. Особенности структуры и химические свойства двойной спирали ДНК облегчают ее репарацию
Молекула ДНК имеет структуру, по-видимому, наилучшим образом приспособленную для репарации. Если гипотеза о том, что РНК
появилась в процессе эволюции раньше, чем ДНК верна (см. разд. 1.1.7), возникает вопрос, почему присутствующий в РНК урацил (U) был в ДНК
заменен на тимин (Т). Очевидно, это можно объяснить тем, что механизм, осуществляющий удаление дезаминированных остатков цитозина (рис. 5-
36), не смог бы функционировать, если бы четвертым нуклеотидом в ДНК был урацил, а не тимин (т. е. не 5-метилурацил). Спонтанное
дезаминирование С дает U, и потому фермент, узнающий и удаляющий такие случайно возникшие остатки U, наряду с ними удалял бы и остатки U,
которые были бы нормальными компонентами этой ДНК.
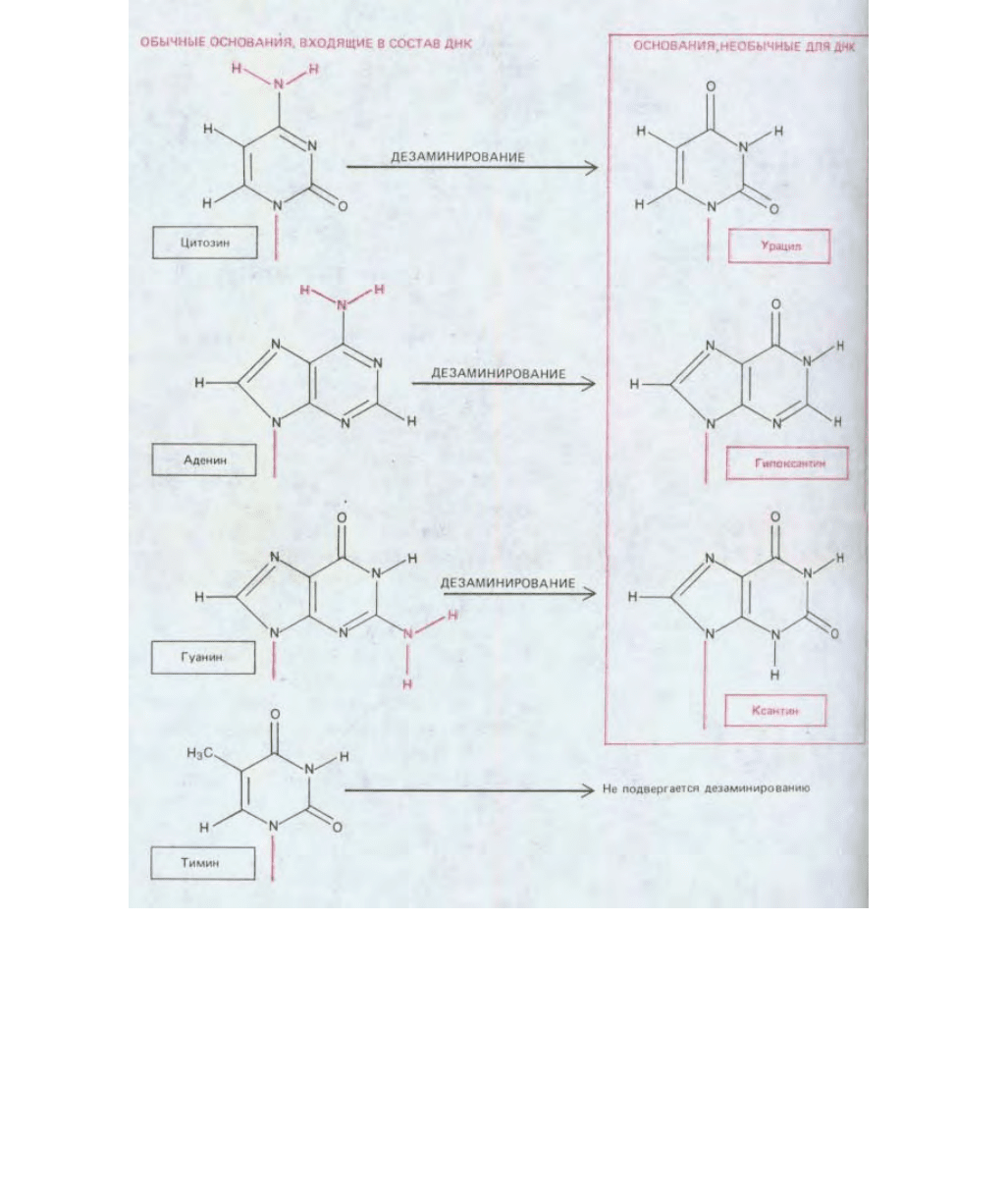
Аналогичным образом обстоит дело и в другом случае, а именно в выборе гуанина вместо гипоксантина. Простейший пурин,
специфически спаривающийся с С, - это гипоксантин. Но гипоксантин является непосредственным продуктом дезаминирования А (рис. 5-37).
Добавив к гипоксантину вторую аминогруппу, эволюция создала гуанин, который не может образоваться из А в результате его спонтанного
дезаминирования. Таким образом, любое возможное дезаминирование в ДНК ведет к появлению необычного основания, которое именно в силу
своей необычности может быть сразу же распознано и удалено специальной ДНК-гликозилазой (рис. 5-37).
Итак, сама химическая природа оснований гарантирует, что дезаминирование не останется незамеченным. Однако точная репарация (а
вместе с тем и радикальное решение шрёдингеровской дилеммы) возможна благодаря существованию двух копий генетической информации,
каждая из которых представлена одной из двух цепей двойной спирали ДНК. Лишь в случае крайне маловероятного события, а именно
одновременного повреждения обоих членов одной и той же пары оснований, в клетке не окажется ни одной правильной копии, которая могла бы
служить матрицей для репарации ДНК.
Генетическая информация может также храниться в одноцепочечной ДНК или РНК, и некоторые очень мелкие вирусы обладают одно-
цепочечными геномами, содержащими лишь несколько тысяч нуклеотидов. Описанные выше механизмы не в состоянии осуществлять репарацию
таких нуклеиновых кислот, и потому частота мутаций у этих вирусов весьма велика. Лишь организмы с совсем крошечными геномами могут
позволить себе хранить генетическую информацию не в двойной спирали ДНК, а в иных структурах.
Рис. 5-36. Путь репарации ДНК с участием урацил-ДНК-гликозилазы, восстанавливающий в цепи ДНК цитозин после его случайного
дезаминирования. После действия ДНК-гликозилазы сахарофосфат, утративший бывшее при нем основание, удаляется из цепи АП-эндонуклеазой,
тем же ферментом, который участвует и в репарации апуринизированных участков. Далее следуют этапы, показанные на рис. 5-33. В названии
«АП-эндонуклеаза» отражен тот факт, что данный фермент распознает в спирали ДНК любой участок, содержащий остаток дезоксирибозы,
утративший бывшее при нем основание. Утраченное основание может быть либо пурином (апуринизированные участки), либо пиримидином
(апиримидинизированные участки).
286
Рис. 5-37. Продукты спонтанного дезаминирования различных оснований ДНК. Все эти продукты дезаминирования необычны в составе ДНК и
распознаются именно по этой причине.
Заключение
Судить о надежности сохранения нуклеотидных последовательностей ДНК у высших эукариот можно, исходя из скорости изменения
аминокислотных последовательностей второстепенных белков и нуклеотидных последовательностей ДНК на протяжении эволюционного
времени. Эта надежность столь велика, что за год в геноме млекопитающего, насчитывающем 3 • 10
9
пар оснований, в среднем происходит всего
лишь 10-20 замен оснований, затрагивающих клетки зародышевой линии. В то же время в геноме такого размера из-за неизбежных процессов
химического распада ежедневно повреждаются тысячи нуклеотидов ДНК. Генетическая информация может надежно храниться в
нуклеотидных последовательностях ДНК лишь потому, что широкий набор различных репарирующих ферментов осуществляет непрерывный
«осмотр» ДНК и удаляет из нее поврежденные нуклеотиды.
Процесс репарации ДНК основан на том, что генетическая информация представлена в этой молекуле двумя копиями - по одной в
каждой из двух цепей двойной спирали ДНК. Благодаря этому случайное повреждение в одной из цепей может быть удалено репарирующим
ферментом и данный участок цепи ресинтезирован в своем нормальном виде за счет информации, содержащейся в неповрежденной цепи.

287
5.3. Механизмы репликации ДНК [24]
Живые организмы должны не только поддерживать целостность нуклеотидных последовательностей ДНК путем ее репарации, но еще и
очень точно воспроизводить свою ДНК перед каждым клеточным делением. При репликации ДНК скорость полимеризации колеблется в пределах
от 500 нуклеотидов в 1 с у бактерий приблизительно до 50 нуклеотидов у млекопитающих. Ясно, что ферменты, катализирующие процесс
репликации, должны работать и точно, и быстро. Быстрота и точность достигаются с помощью особого мультиферментного комплекса,
направляющего процесс репликации. Этот комплекс, состоящий из нескольких различных белков, представляет собой сложный и совершенный
«аппарат репликации».
5.3.1. Репликация ДНК, как и ее репарация, основана на комплементарном спаривании оснований [25]
Матричная активность ДНК проявляется в том, что ее нуклеотидная последовательность копируется (целиком или частично) путем
комплементарного спаривания оснований (А с Т или G с С) в виде комплементарной последовательности нуклеотидов ДНК или РНК. Этот процесс
предполагает узнавание каждого нуклеотида в ДНК свободным (неполимеризованным) комплементарным нуклеотидом и обязательное разделение
(хотя бы на время) двух цепей ДНК, с тем чтобы в каждом основании группы, играющие роль доноров и акцепторов при образовании водородных
связей, оказались доступными для комплементарного спаривания. Таким образом поступающие одиночные нуклеотиды выстраиваются в
определенном порядке вдоль матричной цепи ДНК для ферментативной полимеризации, продуктом которой является новая полинуклеотидная
цепь. В 1957 г. был открыт первый фермент, катализирующий процесс полимеризации нуклеотидов; он был назван ДНК-полимеразой. Было
показано, что субстратами ДНК-полимеразы служат дезоксирибонуклеозидтрифосфаты, полимеризующиеся на одно-цепочечной ДНК-матрице
(двухступенчатый механизм этой полимеризации представлен на рис. 5-34 в связи с обсуждением процесса репарации ДНК). Позже была выделена
и РНК-полимераза, для которой субстратами служат рибонуклеозидтрифосфаты.
Во время репликации ДНК каждая из двух ее старых цепей служит матрицей для образования новой цепи. Поэтому чрезвычайно длинная
нуклеотидная последовательность клеточной ДНК реплицируется, как это принято называть, «полуконсервативно» и каждая из двух дочерних
клеток получает при клеточном делении новую двойную спираль ДНК, состоящую из одной старой и одной новой цепи (см. рис. 3-11).
5-23
5.3.2. Репликационная вилка асимметрична [26]
Исследования, проведенные в начале 1960-х годов на реплицирующихся хромосомах, в которые в качестве импульсной метки вводили
радиоактивный предшественник ДНК
3
Н-тимидин, выявили особую четко ограниченную область репликации, перемещающуюся вдоль
родительской спирали ДНК. Эта активная область из-за своей Y-образной формы была названа репликационной вилкой. Именно в ней с
помощью мультиферментного комплекса, содержащего ДНК-полимеразу, синтезируются дочерние молекулы ДНК.
В то время казалось вполне вероятным, что простейший механизм репликации ДНК заключается в непрерывном росте обеих новых цепей
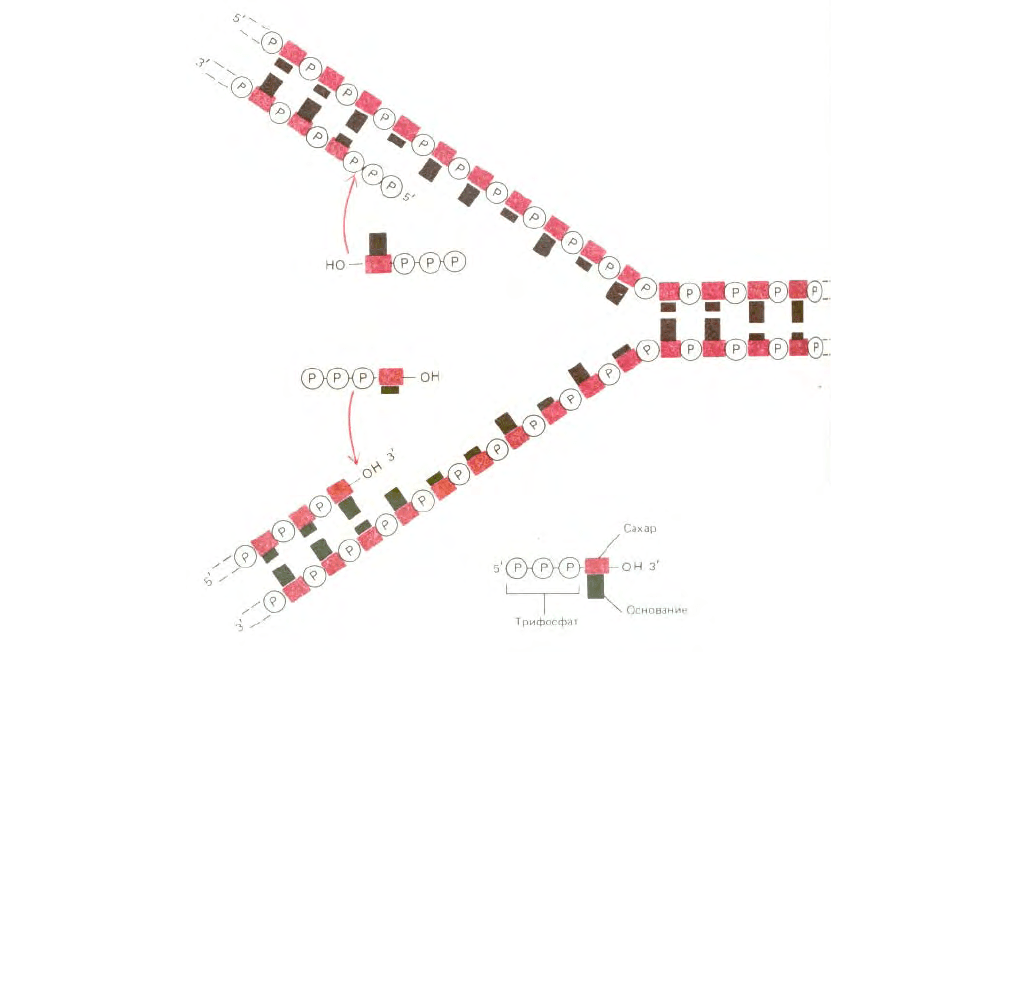
288
Рис. 5-38. На первый взгляд простейшим механизмом репликации ДНК представляется механизм, изображенный на этой (неверной!) схеме. Обе
дочерние цепи должны были бы при этом расти непрерывно за счет присоединения нуклеотидов соответственно в 5' → 3' - направлении (на рисунке
- внизу) и 3' → 5' - направлении (на рисунке - вверху). Однако фермента, который бы катализировал присоединение нуклеотидов в направлении 3' →
5', не существует.
нуклеотид за нуклеотидом по мере перемещения репликационной вилки от одного конца молекулы ДНК к другому. Однако, поскольку две цепи в
спирали ДНК антипараллельны, одна из дочерних цепей должна расти в направлении 5' → 3', а другая - в направлении 3' → 5'. В таком случае
репликационной вилке потребовалось бы две разные ДНК-полимеразы. Одна из них наращивала бы цепь в направлении 5' → 3' (рис. 5-34); при этом
каждый поступающий мономер (дезоксирибонуклеозидтрифосфат) приносит с собой необходимую для его присоединения к цепи энергию (ее
носителем является трифосфатная группа). Другая ДНК-полимераза, перемещающаяся в направлении 3' → 5', должна катализировать «рост с
головы»; в этом случае энергию, необходимую для присоединения каждого очередного нуклеотида, должен нести конец растущей цепи ДНК. В
действительности такой (3' → 5') ДНК-полимеразы не существует (рис. 5-38), хотя биохимикам известны некоторые другие процессы
полимеризации, протекающие по типу «роста с головы» (см. рис. 2-34), Каким же образом происходит рост цепи в направлении 3' → 5'?
Возможный ответ на этот вопрос подсказали в конце 1960-х годов эксперименты с радиоактивно меченными предшественниками ДНК. Если
растущие клетки получают всего на несколько секунд высокорадиоактивный
3
Н-тимидин, то метка включается лишь в ДНК, синтезированную в
самый последний момент, т. е. в ту ее часть, которая следует непосредственно за репликационной вилкой. Этим методом избирательного введения
метки было выявлено, что при репликации бактериальной ДНК в области репликационной вилки образуются и какое-то время существуют
фрагменты, насчитывающие от 1000 до 2000 нуклеотидов (впоследствии за ними закрепилось название «фраг-
289
Рис. 5-39. Строение репликационной вилки. Обе дочерние цепи строятся в направлении 5' → 3'. Для этого отстающая цепь ДНК должна
синтезироваться в виде ряда коротких фрагментов (фрагменты Оказаки).
менты Оказаки»; у эукариот они гораздо короче: от 100 до 200 нуклеотидов). Несколько позже было показано, что синтез этих фрагментов ДНК
идет только в направлении 5' → 3'; синтезированные фрагменты соединяются затем в длинные цепи ДНК под действием того же фермента, который
сшивает разрывы в спирали ДНК во время ее репарации, т.е. под действием ДНК-лигазы (см. рис. 5-35).
Репликационная вилка асимметрична (рис. 5-39). Из двух синтезируемых дочерних цепей ДНК одна строится непрерывной, а другая
прерывистой. Первую называют ведущей (или лидирующей), а вторую - отстающей. Наращивание второй цепи отстает, потому что образование
каждого фрагмента Оказаки оказывается возможным лишь после того, как продвижение ведущей цепи откроет соответствующий участок матрицы.
Хотя в целом вся эта цепь строится в направлении 3' → 5', каждый из ее фрагментов синтезируется в направлении 5' → 3'. Благодаря тому что ДНК
на отстающей части вилки строится при помощи механизма, работающего прерывисто по типу «шитья назад иголкой» (backstitching), в
репликационной вилке не требуется никакого другого фермента, кроме (5' → 3')-ДНК-полимеразы.
5-24
5.3.3. Высокая точность репликации ДНК предполагает наличие механизма, осуществляющего коррекцию [27]
Точность копирования при репликации ДНК столь велика, что в среднем на каждые 1-10
9
комплементарных пар, образующихся в
процессе воспроизведения генома млекопитающих, насчитывающего 3-10
4
пар оснований (см. разд. 9.1.3), приходится приблизительно одна
ошибка. Точность эта значительно превосходит ту, какую следует ожидать, учитывая, что во время репликации образуются не только обычные
комплементарные пары оснований. В нормальной ДНК возникают на короткое время с частотой 10
-4
-10
-5
редкие таутомерные формы всех четырех
ее оснований. Эти формы образуют неправильные пары. Так, редкая таутомерная форма С спаривается с А вместо G, в результате чего возникает
мутация (рис. 5-40). Таким образом высокая точность
Рис. 5-40. Пример возникновения при репликации ДНК неправильной пары оснований: находясь в термодинамически невыгодной таутомерной
форме, цитозин легко образует водородные связи с аденином.

290
Рис. 5-41. Схема, поясняющая, как протекает процесс
коррекции (устранение ошибок) при синтезе ДНК,
катализируемом ДНК-полимеразами у бактерий.
Предполагается, что аналогичный механизм коррекции
действует и в эукариотических клетках.
репликации ДНК определяется наличием механизмов, осуществляющих
коррекцию, т. е. устраняющих подобные ошибки.
Один из важных механизмов коррекции зависит от особых свойств
ДНК-полимеразы. В отличие от РНК-полимераз ДНК-полимеразы не могут
начать синтез новой полинуклеотидной цепи, просто связав друг с другом 3'-ОН-
конец какой-либо полинуклеотидной цепи, которая должна быть спарена с
матричной цепью ДНК; ДНК-полимеразы способны только добавлять новые
нуклеотиды к уже имеющемуся 3'-ОН-концу полинуклеотидной цепи (см. рис. 5-
34). Эту предобразованную цепь, к которой добавляются нуклеотиды, называют
затравкой или праймером. Молекулы ДНК с затравкой, у которой 3'-ОН-конец
не спарен, не могут служить матрицами. Бактериальные ДНК-полимеразы
способны, однако, с ними работать. Вступив в контакт с такими молекулами
ДНК, они используют присущую им (3' → 5')-экзонуклеазную активность и
отщепляют (путем гидролиза) любые неспаренные нуклеотиды на затравочном
конце. Отщепляется ровно столько нуклеотидов, сколько требуется для того,
чтобы у затравки появился спаренный конец и образовалась активная матрица.
Действуя таким образом, ДНК-полимераза выступает в роли
«самокорректирующего» фермента: она устраняет свои собственные ошибки,
возникающие в процессе полимеризации. Рис. 5-41 поясняет, как этот тип
коррекции может использоваться для удаления неправильных пар С—А,
образуемых редкой таутомерной формой цитозина.
Потребность в правильно спаренном конце как раз и наделяет ДНК-
полимеразу способностью исправлять свои собственные ошибки. Такой
фермент, очевидно, мог бы начать синтез ДНК при полном отсутствии затравки,
только утратив способность различать спаренный и неспаренный концы. В то же
время РНК-полимеразы, участвующие в транскрипции генов (см. разд. 5.1.1),
судя по всему не нуждаются в самокоррекции, потому что ошибки транскрипции
не передаются следующему поколению и случайно возникшие дефектные
молекулы особой роли не играют. РНК-полимеразы могут начинать синтез
новых полинуклеотидных цепей в отсутствие затравки, причем ошибки
встречаются с частотой 10
-4
как при синтезе РНК, так и при трансляции, т.е. при
переводе нуклеотидных последовательностей мРНК в аминокислотные
последовательности белков.
5-22
5.3.4. Репликация ДНК в направлении 5' → 3' обеспечивает
эффективную коррекцию
Весьма вероятно, что однонаправленность репликации ДНК (5' → 3')
объясняется высокими требованиями к точности процесса. Если бы
существовала ДНК-полимераза, присоединяющая
дезоксирибонуклеозидтрифосфаты к синтезируемой полинуклеотидной цепи
таким образом, что эта цепь росла в направлении 3' → 5', то активирующую
трифосфатную группировку нес бы растущий 5'-конец цепи, а не поступающий
мононуклеотид. В этом случае ошибки полимеризации не могли бы устраняться
простым гидролизом, потому что появление свободного 5'-конца немедленно
обрывало бы синтез ДНК. Ясно, что основание, только что неправильно
спарившееся на 3'-конце, устранить гораздо легче, чем такое же основание,
присоединившееся к 5'-концу