Benton M.J. Introduction to paleobiology and the fossil record
Подождите немного. Документ загружается.


18 INTRODUCTION TO PALEOBIOLOGY AND THE FOSSIL RECORD
cooperation of many people. The spectacular
Burgess Shale fauna (Gould 1989; Briggs
et al. 1994) was found by the geologist Charles
Walcott in 1909. The discovery was partly by
chance: the story is told of how Walcott and
his wife were riding through the Canadian
Rockies, and her horse supposedly stumbled
on a slab of shale bearing beautifully pre-
served examples of Marrella splendens, the
“lace crab”. During fi ve subsequent fi eld
seasons, Walcott collected over 60,000 speci-
mens, now housed in the National Museum
of Natural History, Washington, DC. The
extensive researches of Walcott, together with
those of many workers since, have docu-
mented a previously unknown assemblage of
remarkable soft-bodied animals. The success
of the work depended on new technology
in the form of high-resolution microscopes,
scanning electron microscopes, X-ray photog-
raphy and computers to enable three-dimen-
sional reconstructions of fl attened fossils. In
addition, the work was only possible because
of the input of thousands of hours of time in
skilled preparation of the delicate fossils, and
in the production of detailed drawings and
descriptions. In total, a variety of government
and private funding sources must have con-
tributed hundreds of thousands of dollars to
the continuing work of collecting, describing
and interpreting the extraordinary Burgess
Shale animals.
The Burgess Shale is a dramatic and unusual
example. Most paleontological research is
more mundane: researchers and students may
spend endless hours splitting slabs, excavating
trenches and picking over sediment from
deep-sea cores under the microscope in order
to recover the fossils of interest. Laboratory
preparation may also be tedious and long-
winded. Successful researchers in paleontol-
ogy, as in any other discipline, need endless
patience and stamina.
Modern paleontological expeditions go all
over the world, and require careful negotia-
tion, planning and fund-raising. A typical
expedition might cost anything from
US$20,000 to $100,000, and fi eld paleontol-
ogists have to spend a great deal of time plan-
ning how to raise that funding from government
science programs, private agencies such as the
National Geographic Society and the Jurassic
Foundation, or from alumni and other spon-
sors. A typical high-profi le example has been
Box 1.4 Giant dinosaurs from Madagascar
How do you go about fi nding a new fossil species, and then telling the world about it? As an example,
we choose a recent dinosaur discovery from the Late Cretaceous of Madagascar, and tell the story
step by step. Isolated dinosaur fossils had been collected by British and French expeditions in the
1880s, but a major collecting effort was needed to see what was really there. Since 1993, a team,
led by David Krause of SUNY-Stony Brook, has traveled to Madagascar for nine fi eld seasons with
funding from the US National Science Foundation and the National Geographic Society. Their work
has brought to light some remarkable new fi nds of birds, mammals, crocodiles and dinosaurs from
the Upper Cretaceous.
One of the major discoveries on the 1998 expedition was a nearly complete skeleton of a titanosau-
rian sauropod. These giant plant-eating dinosaurs were known particularly from South America and
India, though they have a global distribution, and isolated bones had been reported from Madagascar
in 1896. The new fossil was found on a hillside in rocks of the Maevarano Formation, dated at about
70 million years old, in the Mahajanga Basin. The landscape is rough and exposed, and the bones were
excavated under a burning sun. The fi rst hint of discovery was a series of articulated tail vertebrae, but
as the team reported, “The more we dug into the hillside, the more bones we found”. Almost every
bone in the skeleton was preserved, from the tip of the nose, to the tip of the tail. The bones were exca-
vated and carefully wrapped in plaster jackets for transport back to the United States.
Back in the laboratory, the bones were cleaned up and laid out (Fig. 1.12). Kristi Curry Rogers
worked on the giant bones for her PhD dissertation that she completed at SUNY-Stony Brook in
2001. Kristi, and her colleague Cathy Forster, named the new sauropod Rapetosaurus krausei in

PALEONTOLOGY AS A SCIENCE 19
(a)
(b)
Figure 1.12 Finding the most complete titanosaur, Rapetosaurus, in Madagascar: (a) Kristi Curry
Rogers (front right) with colleagues excavating the giant skeleton; (b) after preparation in the lab,
the whole skeleton can be laid out – this is a juvenile sauropod, so not as large as some of its
relatives. (Courtesy of Kristi Curry Rogers.)
2001. It turned out to be different from titanosaurians already named from other parts of the world,
and the specimen was unique in being nearly complete and in preserving the skull, which was
described in detail by Curry Rogers and Forster in 2004. Its name refers to “rapeto”, a legendary
giant in Madagascan folklore. To date, Rapetosaurus krausei is the most complete and best-preserved
titanosaur ever discovered.
Kristi Curry Rogers is now Curator and Head of Vertebrate Paleontology at the Science Museum
of Minnesota, where she continues her work on the anatomy and relationships of sauropod dino-
saurs, and on dinosaur bone histology. Read more about her at http://www.blackwellpublishing.
com/paleobiology/. You can fi nd out more about Rapetosaurus in Curry Rogers and Forster (2001,
2004) and at http://www.blackwellpublishing.com/paleobiology/.
Continued
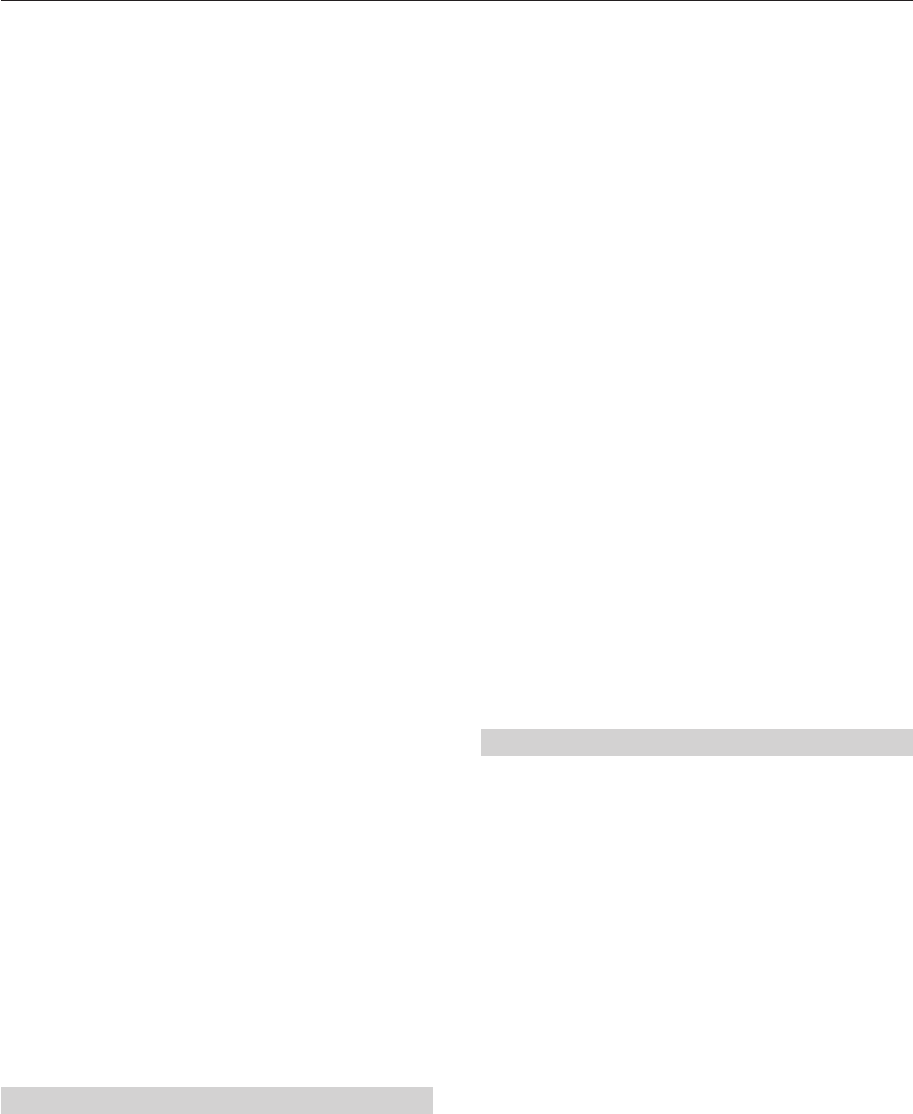
20 INTRODUCTION TO PALEOBIOLOGY AND THE FOSSIL RECORD
a long-running program of study of dinosaurs
and other fossil groups from the Cretaceous
of Madagascar (Box 1.4).
Field expeditions attract wide attention,
but most paleontological research is done in
the laboratory. Paleontologists may be moti-
vated to study fossils for all kinds of reasons,
and their techniques are as broad as in any
science. Paleontologists work with chemists
to understand how fossils are preserved and
to use fossils to interpret ancient climates and
atmospheres. Paleontologists work with engi-
neers and physicists to understand how
ancient animals moved, and with biologists to
understand how ancient organisms lived
and how they are related to each other.
Paleontologists work with mathematicians
to understand all kinds of aspects of
evolution and events, and the biomechanics
and distribution of ancient organisms. Pale-
ontologists, of course, work with geologists
to understand the sequence and dating of
the rocks, and ancient environments and
climates.
But it seems that, despite centuries of study,
paleobiologists have so much to learn. We
don’t have a complete tree of life; we don’t
know how fast diversifi cations can happen
and why some groups exploded onto the scene
and became successful and others did not; we
don’t know the rules of extinction and mass
extinction; we don’t know how life arose
from non-living matter; we don’t know why
so many animal groups acquired skeletons
500 million years ago; we don’t know why
life moved on to land 450 million years ago;
we don’t know exactly what dinosaurs did;
we don’t know what the common ancestor of
chimps and humans looked like and why the
human lineage split off and evolved so fast to
dominate the world. These are exciting times
indeed for new generations to be entering this
dynamic fi eld of study!
Review questions
1 What kinds of evidence might you look
for to determine the speed and mode of
locomotion of an ancient beetle? Assume
you have fossils of the whole body, includ-
ing limbs, of the beetle and its fossilized
tracks.
2 Which of these statements is in the form
of a scientifi c hypothesis that may be
tested and could be rejected, and which
are non-scientifi c statements? Note,
scientifi c hypotheses need not always be
correct; equally, non-scientifi c statements
might well be correct, but cannot be
tested:
• The plant Lepidodendron is known
only from the Carboniferous Period.
• The sabertoothed cat Smilodon ate
plant leaves.
• Tyrannosaurus rex was huge.
• There were two species of Archaeop-
teryx, one larger than the other.
• Evolution did not happen.
• Birds and dinosaurs are close relatives
that share a common ancestor.
3 Do you think scientists should be cautious
and be sure they can never be contra-
dicted, or should they make statements
they believe to be correct, but that can be
rejected on the basis of new evidence?
4 Does paleontology advance by the discov-
ery of new fossils, or by the proposal and
testing of new ideas about evolution and
ancient environments?
5 Should governments invest tax dollars in
paleontological research?
Further reading
Briggs, D.E.G. & Crowther, P.R. 2001. Palaeobiology
II. Blackwell, Oxford.
Bryson, B. 2003. A Short History of Nearly Everything.
Broadway Books, New York.
Buffetaut, E. 1987. A Short History of Vertebrate Pal-
aeontology. Croom Helm, London.
Cowen, R. 2004. The History of Life, 4th edn. Black-
well, Oxford.
Curry Rogers, K. & Forster, C.A. 2001. The last of the
dinosaur titans: a new sauropod from Madagascar.
Nature 412, 530–4.
Curry Rogers, K. & Forster, C.A. 2004. The skull of
Rapetosaurus krausei (Sauropoda: Titanosauria)
from the Late Cretaceous of Madagascar. Journal of
Vertebrate Paleontology 24, 121–44.
Dong Z.-M. & Currie, P.J. 1996. On the discovery of
an oviraptorid skeleton on a nest of eggs at Bayan
Mandahu, Inner Mongolia, People’s Republic of
China. Canadian Journal of Earth Sciences 33,
631–6.
Foote, M. & Miller, A.I. 2006. Principles of Paleontol-
ogy. W.H. Freeman, San Francisco.
Fortey, R. 1999. Life: A Natural History of the First
Four Billion Years of Life on Earth. Vintage Books,
New York.

PALEONTOLOGY AS A SCIENCE 21
Hammer, O. & Harper, D.A.T. 2005. Paleontological
Data Analysis. Blackwell, Oxford.
Kemp, T.S. 1999. Fossils and Evolution. Oxford Uni-
versity Press, Oxford.
Mayr, E. 1991. One Long Argument; Charles Darwin
and the Genesis of Modern Evolutionary Thought.
Harvard University Press, Cambridge, MA.
Palmer, D. 2004. Fossil Revolution: The Finds that
Changed Our View of the Past. Harper Collins,
London.
Rudwick, M.J.S. 1976. The Meaning of Fossils: Epi-
sodes in the History of Paleontology. University of
Chicago Press, Chicago.
Rudwick, M.J.S. 1992. Scenes from Deep Time: Early
Pictorial Representations of the Prehistoric World.
University of Chicago Press, Chicago.
References
Alvarez, L.W., Alvarez, W., Asaro, F. & Michel, H.V.
1980. Extraterrestrial causes for the Cretaceous-
Tertiary extinction. Science 208, 1095–108.
Beringer, J.A.B. 1726. Lithographiae wirceburgensis,
ducentis lapidum fi guatorum, a potiori insectifor-
mium, prodigiosis imaginibus exornatae specimen
primum, quod in dissertatione inaugurali physico-
historica, cum annexis corollariis medicis. Fuggart,
Wurzburg, 116 pp.
Briggs, D.E.G., Erwin, D.H. & Collier, F.J. 1994. The
Fossils of the Burgess Shale. Smithsonian Institution
Press, Washington.
Curry Rogers, K. & Forster, C.A. 2001. The last of the
dinosaur titans: a new sauropod from Madagascar.
Nature 412, 530–4.
Curry Rogers, K. & Forster, C.A. 2004. The skull of
Rapetosaurus krausei (Sauropoda: Titanosauria)
from the Late Cretaceous of Madagascar. Journal of
Vertebrate Paleontology 24, 121–44.
Darwin, C.R. 1859. On the Origin of Species by Means
of Natural Selection, or the Preservation of Favoured
Races in the Struggle for Life. John Murray, London,
502 pp
Gould, S.J. 1989. Wonderful Life. The Burgess Shale
and the Nature of History. Norton, New York.
Hammer, O. & Harper, D.A.T. 2005. Paleontological
Data Analysis. Blackwell, Oxford.
Hunter, W. 1768. Observations on the bones commonly
supposed to be elephant’s bones, which have been
found near the river Ohio, in America. Philosophical
Transactions of the Royal Society 58, 34–45.
Norell, M.A., Clark, J.M., Chiappe, L.M. & Dashzeveg,
D. 1995. A nesting dinosaur. Nature 378, 774–6.
Norell, M.A., Clark, J.M., Dashzeveg, D. et al. 1994.
A theropod dinosaur embryo and the affi nities of
the Flaming Cliffs Dinosaur eggs. Science 266,
779–82.

Chapter 2
Fossils in time and space
Key points
• Scientists began to study the order and sequence of geological events during the Renais-
sance when artists rediscovered perspective.
• Lithostratigraphy is the establishment of rock units, forming the basis for virtually all
geological studies; lithostratigraphic units are displayed on maps and measured
sections.
• Biostratigraphy, using zone fossils, forms the basis for correlation and it can now be
investigated using a range of quantitative techniques.
• Chronostratigraphy, global standard stratigraphy, is the division of geological time into
workable intervals with reference to type sections in the fi eld.
• Cyclostratigraphy and sequence stratigraphy can provide more refi ned frameworks that
can also help understand biological change.
• Geochronometry is based on absolute time, measured in years before present by a range
of modern, quantitative techniques.
• Paleobiogeography provides basic data to suggest and test plate tectonic and terrane
models.
• Changes in geography allowed faunas and fl oras to migrate, and major groups to radiate
and go extinct.
• The rhythmic joining and break up of continents through time has been associated with
climate and diversity change.
• Fossils from mountain belts are signifi cant in constraining the age and origin of tectonic
events; fossil data have also provided estimates for fi nite strain and thermal
maturation.
FOSSILS IN TIME AND SPACE 23
The Earth is immensely old, and the distribu-
tion of continents and oceans has changed
radically over time. Early paleontologists did
not know these things, and so they tried to
pack the whole of the history of life into a
relatively short span of time, vizualizing trilo-
bites or dinosaurs inhabiting a world that was
much as it is today.
Life on Earth, however, has been evolving
for up to 4 billion years, and there has been a
complex story of fossil groups coming and
going, and continents moving from place to
place. How do we develop geographic and
temporal frameworks that are accurate and
reliable enough to chart the distributions of
fossil organisms through time and space? For-
tunately, paleogeographers and stratigraphers
are now equipped with a range of high-tech
methods, virtually all computer-based, that
provide a greater consensus for models describ-
ing the distributions of the continents, oceans
and their biotas throughout geological time.
Fossils also store information on the fi nite
strain and thermal maturation of rocks located
in the planet’s mountain belts, allowing the
tectonic history of these ranges to be recon-
structed; thermal maturation information is
important in identifying the levels of thermal
maturity of rocks and the gas and oil windows
in hydrocarbon exploration. In some cases
fossil shells also contain isotopes and other
geochemical information that can identify
changes in global climate (see p. 111).
FRAMEWORKS
Six distinct aspects of Tuscany we there-
fore recognize, two when it was fl uid,
two when level and dry, two when it was
broken; and as I prove this fact concern-
ing Tuscany by inference from many
places examined by me, so do I affi rm it
with reference to the entire earth, from
the descriptions of different places con-
tributed by different writers.
Nicolaus Steno (1669) The
Prodromus of Nicolaus Steno’s
Dissertation Concerning a Solid Body
Enclosed by Process of Nature
Within a Solid
Before the distributions of fossils in time
and space can be described, analyzed and
interpreted, fossil animals and plants must be
described in their stratigraphic context. A
rock stratigraphy is the essential framework
that geologists and particularly paleontolo-
gists use to accurately locate fossil collections
in both temporal and spatial frameworks. It
seems, not surprisingly, that like a fi ne bottle
of Italian wine, this can be traced back to the
sunny, pastel landscapes of Tuscany and the
Renaissance.
Leonardo’s legacy
The origin of modern stratigraphy can be
traced back to Leonardo da Vinci and his
drawings. Pioneer work by the Danish poly-
math Nicolaus Steno (Niels Stensen) in north-
ern Italy, during the late 17th century (see
p. 11), established the simple fact that older
rocks are overlain by younger rocks if the
sequence has not been inverted (Fig. 2.1a).
His law of superposition of strata is funda-
mental to all stratigraphic studies. In addi-
tion, Steno established in experiments that
sediments are deposited horizontally and
rock units can be traced laterally, often for
considerable distances; remarkably simple
concepts to us now, but earth shattering at
the time. But what has this got to do with
da Vinci?
Leonardo da Vinci (1452–1519) is famous
for many things, and his contributions to
science are refreshingly modern when we look
back at them. In his art, da Vinci essentially
rediscovered geological perspective, some 200
years before Steno, during the Renaissance
(Rosenberg 2001). In his drawing of the hills
of Tuscany, da Vinci portrayed a clear sequence
of laterally-continuous, horizontal strata dis-
playing the concept of superposition. More-
over, about a century after Steno, Giovanni
Arduino recognized, again using superposi-
tion, three basically different rocks suites in
the Italian part of the Alpine belt. A crystal-
line basement of older rocks, deformed during
the Late Paleozoic Variscan orogeny, was
overlain unconformably by mainly Mesozoic
limestones deformed later during the Alpine
orogeny; these in turn were overlain uncon-
formably by poorly consolidated clastic rocks,
mainly conglomerates. These three units con-
stituted his primary, secondary and tertiary
systems; the last term has been retained and
formalized for the period of geological time
24 INTRODUCTION TO PALEOBIOLOGY AND THE FOSSIL RECORD
succeeding the Cretaceous (Fig. 2.1b). These
three divisions were used widely to describe
rock successions elsewhere in Europe showing
the same patterns, but these three systems
were not necessarily the time correlatives of
the type succession in the Apennines.
There is now a range of different types of
stratigraphies based on, for example, lithol-
ogy (lithostratigraphy), fossils (biostratigra-
phy), tectonic units, such as thrust sheets
(tectonostratigraphy), magnetic polarity
(magnetostratigraphy), chemical composi-
(a)
Top
Base
Tertiary
Secondary
Primary
poorly consolidated
Tertiary molasse
eroded from rising
Alpine belt
cover sequence
of Mesozoic
sediment folded
during Alpine orogeny
Variscan
basement of
granites and
metamorphics(b)
L
o
n
d
o
n
C
l
a
y
(c)
CoalMountains
tract
Marl
vales
Stonebrash
hills
Clay
vales
Chalk
hills
Plains
Snowdon
Arenig
Aran Vawddry
Snowdon
Range
Caernarvon
Dolgelle
Berwyn Range
Llanfair
Killas Slate and
other strata
Red Rhab
and Dunstone
Coal
Red
Marl
Lias
Blue
Marl
Oolites
Clunch
Clay
Chalk
London
Brickearth
Longmont
Brown Clee
Abberley Hills
Worcester
Broadway
Witney
Wendover
Beaconsfield
S
a
n
d
s
a
n
d
Measures
Figure 2.1 (a) Steno’s series of diagrams illustrating the deposition of strata, their erosion and
subsequent collapse (25, 24 and 23) followed by deposition of further successions (22, 21 and 20).
These diagrams demonstrate not only superposition but also the concept of unconformity. (b) Giovanni
Arduino’s primary, secondary and tertiary systems, fi rst described from the Apennines of northern Italy
in 1760. These divisions were built on the basis of Steno’s Law of Superposition of Strata. (c) Idealized
sketch of William Smith’s geological traverse from London to Wales; this traverse formed the template
for the fi rst geological map of England and Wales. Data assembled during this horse-back survey were
instrumental in the formulation of the Law of Correlation by Fossils. (a, from Steno 1669; c, based on
Sheppard, T. 1917. Proc. Yorks. Geol. Soc. 19.)

FOSSILS IN TIME AND SPACE 25
tions (chemostratigraphy), discontinuities
(allostratigraphy), seismic data (seismic stra-
tigraphy) and depositional trends (cyclo- and
sequence stratigraphies). The fi rst two have
most application in paleontological studies,
although sequence and cyclostratigraphic
frameworks are now providing greater insights
into the climatic and environmental settings
of fossil assemblages. Here, however, we
concentrate on lithostratigraphy (rock frame-
work), biostratigraphy (ranges of fossils) and
chronostratigraphy (time dimension).
ON THE GROUND: LITHOSTRATIGRAPHY
All aspects of stratigraphy start from the rocks
themselves. Their order and succession, or
lithostratigraphy, are the building blocks for
any study of biological and geological change
through time. Basic stratigraphic data are fi rst
assembled and mapped through the defi nition
of a lithostratigraphic scheme at a local and
regional level. Lithostratigraphic units are
recognized on the basis of rock type. The
formation, a rock unit that can be mapped
and recognized across country, irrespective of
thickness, is the basic lithostratigraphic cate-
gory. A formation may comprise one or several
related lithologies, different from units above
and below, and usually given a local geo-
graphic term. A member is a more local litho-
logic development, usually part of a formation,
whereas a succession of contiguous forma-
tions, with some common characteristics is
often defi ned as a group; groups themselves
may comprise a supergroup. All stratigraphic
units must be defi ned at a reference or type
section in a specifi ed area. Unfortunately, the
entire thickness of many lithostratigraphic
units is rarely exposed; instead of defi ning the
whole formation, the bases of units are defi ned
routinely in basal stratotype sections at a type
locality and the entire succession is then pieced
together later. These sections, like yardsticks
or the holotypes of fossils (see p. 118), act as
the defi nitive section for the respective strati-
graphic units. These are defi ned within a rock
succession at a specifi c horizon, where there
is a lithologic boundary between the two
units; the precise boundary is marked on a
stratigraphic log. Since the base of the suc-
ceeding unit defi nes the top of the underlying
unit, only basal stratotypes need ever be
defi ned.
A stratigraphy, illustrated on a map and in
measured sections, is required to monitor bio-
logical and geological changes through time
and thus underpins the whole basis of Earth
history. It is a simple but effective procedure.
Successions of rock are often divided by gaps
or unconformities. These surfaces separate an
older part of the succession that may have
been folded and uplifted before the younger
part was deposited. Commonly there is a
marked difference between the attitudes of
the older and younger parts of the succession;
but sometimes both parts appear conformable
and only after investigation of their fossil
content, is it clear that the surface represents
a large gap in time.
Early geologists thought the Earth was very
young, but the Scottish scientist James Hutton
(1726–1797) noted the great cyclic process of
mountain uplift, followed by erosion, sedi-
ment transport by rivers, deposition in the
sea, and then uplift again, and argued that
such processes had been going on all through
Earth’s history. He wrote in his Theory of the
Earth (1795) that his understanding of geo-
logical time gave “no vestige of a beginning,
– no prospect of an end”. An example of
Hutton’s evidence is the spectacular uncon-
formity at Siccar Point, Berwickshire, south-
ern Scotland, where near-horizontal Old Red
Sandstone (Devonian) strata overlie steeply-
dipping Silurian greywackes. Beneath the
unconformity, Hutton recognized the “ruins
of an earlier world”, establishing the immen-
sity of geological time. This paved the way for
our present concept of the Earth as a dynamic
and changing system, a forerunner to the
current Gaia hypothesis, which describes the
Earth as a living organism in equilibrium with
its biosphere. Although the Earth is not
actually a living organism, this concept now
forms the basis for Earth system science.
USE OF FOSSILS: DISCOVERY OF
BIOSTRATIGRAPHY
Our understanding of the role of fossils in
stratigraphy can be traced back to the work
of William Smith in Britain and Georges
Cuvier and Alexandre Brongniart in France.
William Smith (1769–1839), in the course of
his work as a canal engineer in England, real-
ized that different rocks units were character-

26 INTRODUCTION TO PALEOBIOLOGY AND THE FOSSIL RECORD
ized by distinctive groups or assemblages of
fossils. In a traverse from Wales to London,
Smith encountered successively younger
groups of rocks, and he documented the
change from the trilobite-dominated assem-
blages of the Lower Paleozoic of Wales
through Upper Paleozoic sequences with
corals and thick Mesozoic successions with
ammonites; fi nally he reached the molluskan
faunas of the Tertiary strata of the London
Basin (Fig. 2.1c). In France, a little later, the
noted anatomist Georges Cuvier (see p. 12)
together with Alexandre Brongniart (1770–
1849), a leading mollusk expert of the time,
ordered and correlated Tertiary strata in the
Paris Basin using series of mainly terrestrial
vertebrate faunas, occurring in sequences sep-
arated by supposed biological catastrophes.
These early studies set the scene for bio-
stratigraphic correlation. In very broad terms,
the marine Paleozoic is dominated by bra-
chiopods, trilobites and graptolites, whereas
the Mesozoic assemblages have ammonites,
belemnites, marine reptiles and dinosaurs as
important components, and the Cenozoic is
dominated by mammals and molluskan
groups, such as the bivalves and the gastro-
pods. This concept was later expanded by
John Phillips (1800–1874), who formally
defi ned the three great eras, Paleozoic (“ancient
life”), Mesozoic (“middle life”) and Cenozoic
(“recent life”), based on their contrasting
fossils, each apparently separated by an
extinction event. Many more precise biotic
changes can, however, be tracked at the species
and subspecies levels through morphological
changes along phylogenetic lineages. Very
accurate correlation is now possible using a
wide variety of fossil organisms (see below).
Biostratigraphy: the means of correlation
Biostratigraphy is the establishment of fossil-
based successions and their use in stratigraphic
correlation. Measurements of the stratigraphic
ranges of fossils, or assemblages of fossils,
form the basis for the defi nition of biozones,
the main operational units of a biostratigra-
phy. But the use of such zone fossils is not
without problems. Critics have argued that
there can be diffi culties with the identifi ca-
tions of some organisms fl agged as zone
fossils; and, moreover, it may be impossible
to determine the entire global range of a fossil
or a fossil assemblage, so long as fossils can
be reworked into younger strata by erosion
and redeposition, but this is relatively rare.
Nonetheless, to date, the use of fossils in bio-
stratigraphy is still the best and usually the
most accurate routine means of correlating
and establishing the relative ages of strata. In
order to correlate strata, fossils are normally
organized into assemblage or range zones.
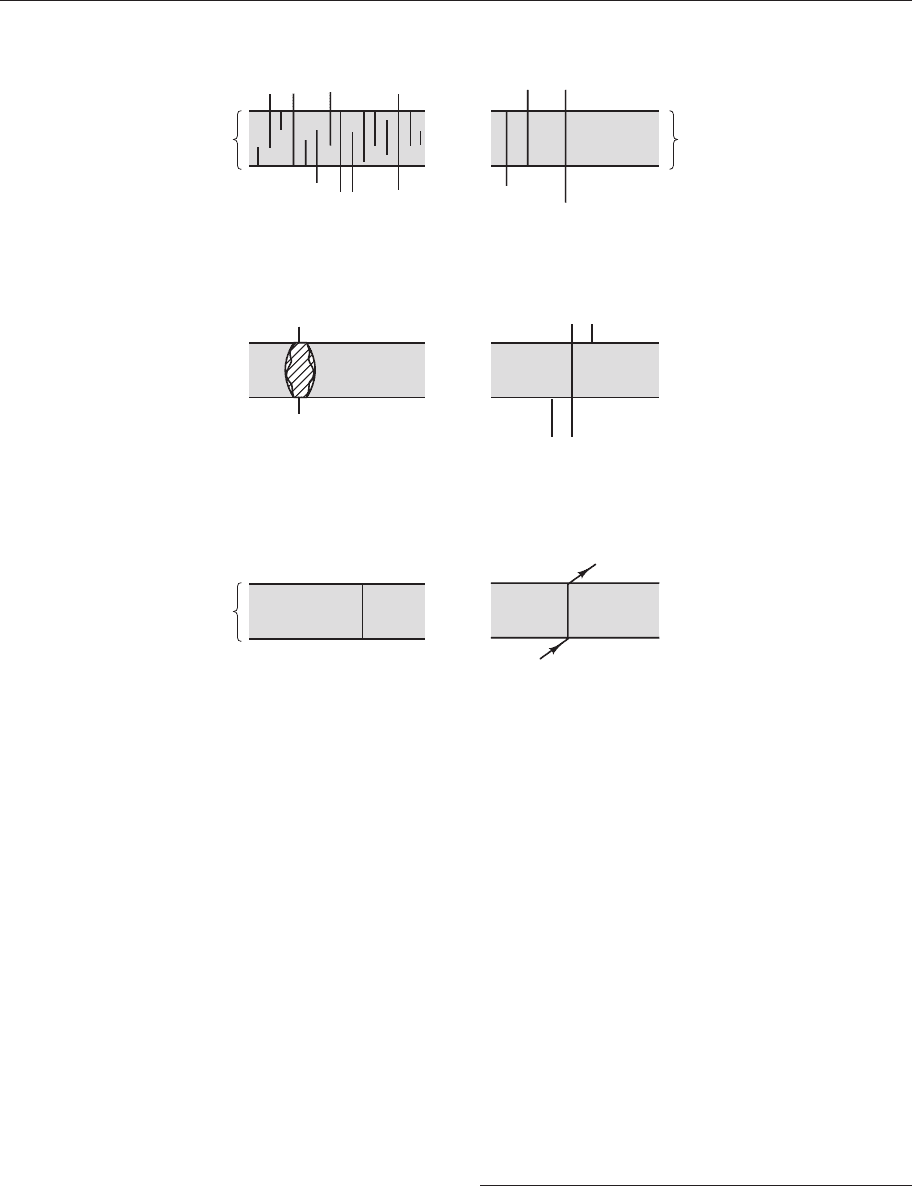
There are several types of range zone (Fig.
2.2); some are used more often than others.
The concept of the range zone is based on the
work of Albert Oppel (1831–1865). Oppel
characterized successive lithologic units by
unique associations of species; his zones were
based on the consistent and exclusive occur-
rence of mainly ammonite species through
Jurassic sections across Europe, where he rec-
ognized 33 zones in comparison with the 60
or so known today. His zonal scheme could
be meshed with Alcide d’Orbigny’s (1802–
1857) stage classifi cation of the system, based
on local sections with geographic terms,
further developed by Friedrich Quenstedt
(1809–1889). Although William Smith had
recognized the signifi cance of fossils almost
50 years previously, Oppel established a
modern and rigorous methodology that now
underpins much of modern biostratigraphy.
The known range of a zone fossil (Box 2.1)
is the time between its fi rst and last appear-
ances in a specifi c rock section, or fi rst appear-
ance datum (FAD) and last appearance datum
(LAD). Clearly, it is unlikely that the entire
global vertical range of the zone fossil is rep-
resented in any one section; nevertheless it is,
in most cases, a workable approximation.
This range, measured against the lithostratig-
raphy, is termed a biozone. It is the basic
biostratigraphic unit, analogous to the
lithostratigraphic formation. It too can be
defi ned with reference to precise occurrences
in the rock, and is defi ned again on the basis
of a stratotype or basal stratotype section in
a type area. Once biozones have been estab-
lished, quantitative techniques may be used to
understand the relationships between rock
thickness and time, and to make links from
locality to locality (Box 2.2).
This is all very well, of course, but the fossil
record is rarely complete; only a small per-
centage of potential fossils are ever preserved.
Stratigraphic ranges can also be infl uenced by
the Signor–Lipps effect (Signor & Lipps 1982),
FOSSILS IN TIME AND SPACE 27
the observation that stratigraphic ranges are
always shorter than the true range of a species,
i.e. you never fi nd the last fossil of a species.
So, incomplete sampling means that the dis-
appearances of taxa may be “smeared” back
in time from the actual point of disappear-
ance. The Signor–Lipps effect is particularly
relevant to mass extinctions, when this
backsmearing can make relatively sudden
extinction events appear gradual. This can be
corrected to some extent by the use of statisti-
cal techniques to establish confi dence inter-
vals that are modeled on known sampling
quality (see p. 165).
Many different animal and plant groups
are used in biostratigraphic correlation (Fig.
2.5). Graptolites and ammonites are the best
known and most reliable zone macrofossils
with their respective biozones as short as
1 myr and 25 kyr, respectively. The most
unusual zone fossils are perhaps those of pigs,
which have been used to subdivide time zones
in the Quaternary rocks of East Africa where
hominid remains occur. Microfossil groups
such as conodonts, dinofl agellates, foraminif-
erans and plant spores are now widely used
(see pp. 209–32, 493–7), particularly in petro-
leum exploration. Microfossils approach the
ideal zone fossils since they are usually
common in small samples, such as drill cores
and chippings, of many sedimentary litholo-
gies and many groups are widespread and
rapidly evolving. The only drawback is that
some techniques used to extract them from
rocks and sediments are specialized, involving
acid digestion and thin sections.
Dividing up geological time: chronostratigraphy
Geological time was divided up by the efforts
of British, French and German geologists
between 1790 and 1840 (Table 2.1). The divi-
sions were made fi rst for practical reasons –
one of the fi rst systems to be named was the
Assemblage biozone
Concurrent-range
biozone
2
3
47
1
5
6
11 14
15
89
10 12
13
A
B
C
B
B
C
C
Ranges of 15 taxa are shown
Biozone defined by complete
assemblage
strata of
biozone in
question
Ranges of 3 taxa, A–C, are shown
Biozone defined by overlap of these
Acme biozone
Biozone defined by exceptional
abundance of one taxon
Partial-range biozone
A
A
strata of
biozone in
question
strata of
biozone in
question
Biozone defined by total or local
range of one taxon
Biozone defined as within the range of
fossil group B, above the last appearance
of fossil group A and below the first
appearance of fossil group C
Total-range biozone
(or local-range
biozone)
Consecutive-range
biozone
Biozone defined by the range of one
taxon, B of lineage A → B → C
Figure 2.2 The main types of biozone, the operational units of a biostratigraphy. (Based on Holland
1986.)