Odekon M. Encyclopedia of paleoclimatology and ancient environments
Подождите немного. Документ загружается.

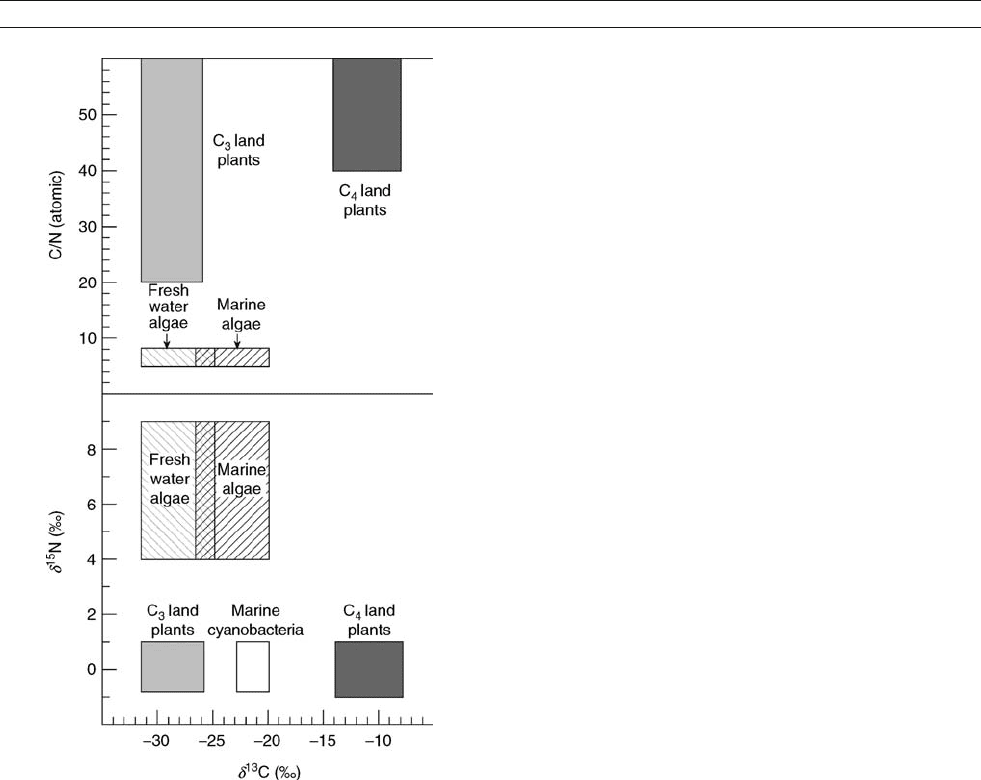
is preserved in the d
15
N values of organic matter from algae and
from land plants (Figure O28, bottom) because the limited avail-
ability of nitrogen discourages much isotopic discrimination.
Denitrification occurs during dysoxic conditions in lakes and
oceans and strongly discriminates against
15
N to yield N
2
enriched in
14
N and residual nitrate enriched in
15
N. These condi-
tions accompany the strong stratification that seasonally happens
in many lakes and is a permanent feature of some oceanic areas.
Subsequent uptake of the residual nitrate by algae leads to
organic matter with high (12–14%) d
15
N values in the sediment
records of these settings. Conversely, good ventilation of the
water column is recorded as relatively low (3–4%) d
15
Nvalues.
Nitrogen-fixing organisms such as cyanobacteria are auto-
trophs that convert dissolved N
2
to bioavailable NH
4
+
when
anoxia exists within the photic zone of lakes and oceans. Their
activity leads to lower d
15
N values in sedimented organic mat-
ter. Evidence of this special environmental condition exists in
sediments deposited between 10 to 3.3 ka in Lake Bosumtwi,
Ghana, where it records a period of strong stratification in this
lake (Talbot and Johannessen, 1992). Suitable conditions for
cyanobacterial activity also commonly occur in eutrophic lakes.
Sediments deposited in central Florida lakes display shifts to
low (1to3%) d
15
N values as eutrophication of these lakes
increased (Brenner et al., 1999). The remarkably low (4to
0%) d
15
N values of Cretaceous black shales (Rau et al.,
1987) and of Pliocene-Pleistocene sapropels in the Mediterra-
nean Sea (Struck et al., 2001) imply strong salinity stratifica-
tion of these marine settings and consequent enhancement of
cyanobacterial primary production. As such, these sediment
records indicate periods of wetter paleoclimatic conditions that
were maintained for multi-thousand-year periods.
Sediment d
15
N values are also sometimes useful in recon-
structing algal paleoproductivity histories, especially in lakes
where phosphorus is often more biolimiting than nitrogen. In
nitrogen-replete environments, biota discriminate in favor of
14
N, and sediment d
15
N values are relatively low. During times
of amplified phosphorus delivery, either from human activities
or from climate-enhanced weathering of regolith, increased algal
utilization of bioavailable nitrogen and consequent decreased
discrimination in favor of
14
N appear as shifts to larger d
15
N
values in sediments (Meyers and Lallier-Vergès, 1999).
Biomarker molecules
Biomarker molecules are compounds that characterize biotic
sources and that retain their source information after burial
in sediments (Table O2). Biomarkers in sediments and rocks
are usually diagenetically stabilized derivatives of specific
precursor compounds, yet the carbon skeletons of the derived
molecules preserve the source-distinctive structures of their
precursor compounds. Although biomarkers typically con-
stitute less than 1% of sediment organic matter, they yield
important paleoenvironmental information that bulk organic
geochemical proxies cannot provide.
Hydrocarbons
Because of their low susceptibility to microbial degradation
relative to other types of organic matter, saturated hydrocar-
bons are generally robust recorders of the origins of organic
matter in sediments. The hydrocarbon compositions of many
aquatic algae and photosynthetic bacteria are dominated by
the C
17
n-alkane (Cranwell et al., 1987). Vascular land plants
contain large proportions of C
27
,C
29
,C
31
, and C
33
n-alkanes
in their waxy coatings, and the biomarker constitutions are
indicative of different kinds of plants. The n-alkane distribu-
tions of grasses and herbs are dominated by C
31
, birches by
C
27
, and most trees by C
29
(Schwark et al., 2002).
Fatty acids
Fatty acids in lake sediments typically originate from multiple
sources. The n-C
16
and n-C
18
alkanoic and alkenoic acids
are ubiquitous components of biota, whereas the even-chain
C
24
–C
30
n-alkanoic acids originate principally from the waxy
coatings of land plants (Cranwell et al., 1987). The long-chain
n-acid components of organic matter in sediments therefore
represent transport of land-derived debris to the depositional
setting. Unsaturated n-C
16
and n-C
18
acids are major constitu-
ents of the lipids of algae, yet they are rapidly degraded by
microbes during and after sedimentation. The microbes contri-
bute their own distinctive biomarker acids to the sediments. For
example, normal and anteiso C
15
alkanoic acids have been
used as indicators of microbial biomass in lake sediments
(Goossens et al., 1989), and they represent in situ production
of secondary lipids at the expense of primary organic matter.
Figure O28 Generalized elemental and carbon isotopic (top) and
nitrogen and carbon isotopic (bottom) compositions of organic matter
from marine and freshwater algae, C
3
and C
4
land plants, and
cyanobacteria.
660 ORGANIC GEOCHEMICAL PROXIES

Unlike the shorter chain-length fatty acids, the longer chain-
length ones survive in sediments and provide evidence of their
plant origins.
Fatty alcohols
Fatty alcohols have source patterns roughly similar to those of
fatty acids. Epicuticular waxes of land plants contain even-
chain n-alkanols from C
22
to C
30
(Rieley et al., 1991). How-
ever, unlike fatty acids, n-alkanol distributions dominated by
the C
16
to C
22
components generally indicate organic matter
derived from aquatic algae and bacteria and not biota in general
(Volkman et al., 1999). Individual species of plants can have
distinctive chain-length patterns that deviate from these gener-
alized distributions.
Alkenones
Long-chain (n-C
37
, n-C
38
, n-C
39
) methyl and ethyl alkenones
are distinctive biomarkers of marine coccolithophores. Their
abundance in sedimentary records can be used as a paleopro-
ductivity proxy for these algae, but their special usefulness is
as paleotemperature proxies. Alkenone distributions usually
consist of a series of homologs having a mixture of 1, 2, 3,
and 4 double bonds that is temperature-sensitive. The algae
produce proportionally more polyunsaturated homologs as
water temperatures decreases in order to maintain cell-wall
fluidity. Alkenones also appear in lake sediment records (Zink
et al., 2001), but their utility as a paleoenvironmental proxy
is not yet established because their origin is unknown.
Sterols and their derivatives
Sterol compositions distinguish aquatic contributions of
organic matter in sediments from land-derived material. Cho-
lesterol (C
27
) is the dominant algal sterol, whereas b-sitosterol
(C
29
) is the major sterol in emergent water plants (Nishimura
and Koyama, 1977). Land-plant leaf waxes contain C
28
and
C
29
but not C
27
sterols (Rieley et al., 1991). Ternary plots of
the C
27
,C
28
, and C
29
sterol compositions of algae, vascular
plants, soils, and sediments identify ecological patterns (Huang
and Meinschein, 1979) that can be applied to paleoenviron-
mental interpretations. Specific sterols are particularly useful
to paleoenvironmental reconstructions. Stronger wind-induced
upwelling is indicated from increased amounts of dinosterol
and C
27
+C
28
4-methylstanols, which are dinoflagellate bio-
markers, in glacial-age sediments in the equatorial Pacific
(Prahl et al., 1989).
Pigments
Plants have evolved a variety of photosynthetic pigments that
can serve as biomarkers. Chlorophyll a, the most common pig-
ment, is non-specific, but aquatic plants and microbes produce
distinctive secondary pigments (Table O2) that facilitate photo-
synthesis in water. Human-induced eutrophication of Esthwaite
Water in the English Lake District is recorded as progressive
increases in the concentrations of myxoxanthophyll and oscil-
laxanthin, two carotenoids indicative of cyanophytes and hence
conditions of elevated lake productivity (Griffiths, 1978). Climate-
induced strong surface stratification of the Mediterranean
Sea during times of Pliocene sapropel formation is implied by
Table O2 Examples of important biomarker molecules and their significance to organic geochemical reconstructions of paleoenvironmental and
paleoclimatic histories
Biomarker molecules Source/ significance
Hydrocarbons
C
27
,C
29
,C
31
and C
33
n-alkanes Vascular land plant waxes; indicates land-plant organic matter contributions and community compositions
C
15
and C
17
n-alkanes Aquatic algae; indicates aquatic organic matter
7- and 8-methyl-branched C
17
alkanes Cyanobacteria, indicates aquatic nitrogen fixation
Pristane Phytol sidechain of chlorophyll; indicates zooplankton-based food chains
Phytane Phytol sidechain of chlorophyll; methanogens; halophilic bacteria; indicates anoxic conditions
Botryococcane Mat-forming freshwater alga Botryococcus braunii
Fichtelite Conifer resins; indicates reducing depositional conditions
Retene Conifer resins; indicates oxidizing depositional conditions
Fatty acids
C
24
,C
26
, and C
28
n-alkanoic acids Vascular land plant waxes
Iso and anteiso branched acids Bacteria; evidence of microbial organic matter
Fatty alcohols
C
24
,C
26
, and C
28
n-alkanols Vascular land plant waxes
Alkenones
C
37
,C
38
, and C
39
alkenones Marine haptophyte algae; proxies for sea surface paleotemperatures
Sterols and steroids
Dinosterol and derivatives Dinoflagellate-derived organic matter
4-methyl sterols Dinoflagellate-derived organic matter
Brassicasterol Diatom-derived organic matter
C
29
sterols Land-plant organic matter
C
27
sterols Marine or freshwater algal organic matter
Pigments
Fucoxanthin, diadinoxanthin Diatom-derived organic matter
Peridinin Dinoflagellate-derived organic matter
Myxoxanthophyll, oscillaxanthin Cyanobacteria; indicates aquatic nitrogen fixation
Isorenieratene derivatives Green sulfur bacteria; anoxic conditions in photic zone
Lignin-derived phenols
Cinnamyl/ vanillyl ratio Proportions of woody and non-woody land-plant tissues
Syringyl/ vanillyl ratio Proportions of gymnosperm and angiosperm land-plant organic matter
ORGANIC GEOCHEMICAL PROXIES 661

isorenieratene derivatives in the sapropel layers but not outside
them (Passier et al., 1999). Isorenieratene is a carotenoid that is
diagnostic for green sulfur bacteria, which are photosynthetic
obligate anaerobes, and their presence means that a near-
surface anoxic zone intruded into the photic zone.
Lignin derivatives
Lignins are phenolic polymers that are synthesized by higher
plants to construct parts of their vascular and structural sys-
tems. Nearly all vascular plants grow on land, and therefore
the lignin fraction of sediment organic matter largely records
the contribution and preservation of land-plant materials. Gym-
nosperms and angiosperms synthesize distinctive types of lig-
nin components. Past changes in continental vegetation can
therefore be inferred from the kinds of lignin found in sediment
records. Molecular analysis of lignin typically begins with
an aggressive oxidation step to break down the biopolymer
into various phenolic monomeric fragments. The oxidation
fragments of lignin have been grouped by Hedges and
Mann (1979) as a guide to identifying their plant sources
(Table O2). A measure of the relative contributions of woody
and non-woody land-plant tissues is given by the C/V (cinnamyl/
vanillyl) ratio, which is the sum of p-coumaric acid plus ferulic
acid concentrations divided by the sum of the three vanillyl
phenols. Gymnosperm sources of land-plant residues are distin-
guished from angiosperm sources by the S/V (syringyl/vanillyl)
ratio, which is the sum of the amounts of the three syringyl phenols
divided by the sum of the vanillyl phenols.
Compound-specific isotopic compositions
Isotopic analysis of individual biomarker compounds provides a
powerful source of paleoclimatic and paleoenvironmental infor-
mation. Compound-specific stable isotope analysis improves
identification of the biological origin of specific components
within the complex mixture of materials that constitutes sedi-
ment organic matter.
Compound-specific d
13
C values
Studies of the d
13
C values of biomarkers have been especially
important in tracing the evolution of C
4
plants. A shift to
less negative d
13
C values of individual plant-wax C
27
to C
33
n-alkanes and C
24
to C
30
n-alkanols enabled Freeman and Colar-
usso (2001) to identify a dramatic expansion of C
4
vegetation on
the Indian subcontinent between 8 and 6 Ma that has persisted to
modern times. This paleoecological change reflects uplift of the
T ibeta n Plateau and associated growth of regional aridity.
The distinctive isotopic compositions of biomarkers from C
3
and C
4
plants also trace Holocene environmental changes.
Sediments of Lake Baikal, Siberia, exhibit a 3% shift to more
negative bulk organic matter d
13
C values that might record a
shift from glacial-age C
4
to post-glacial C
3
plants. Brincat et al.
(2000) show from d
13
C values of individual C
27
to C
31
n-alkanes in the sediment record that C
3
plants have remained
the dominant vegetation throughout this period. Instead of
recording a floral transition, the bulk organic carbon isotopic
shift indicates increased fluvial delivery of isotopically light
soil-derived carbon to the lake in response to a change to
locally wetter post-glacial climate.
Applications of multiple biomarker compound-specific car-
bon isotopic measurements are especially fruitful for detailed
paleoenvironmental reconstructions. Filley et al. (2001) utilize
the d
13
C values of biomarkers from vascular plants (lignin
derivatives, C
29
and C
31
n-alkanes, C
26
n-alkanol), cyanobac-
teria (7- and 8-methylheptadecanes), and phytoplankton (C
17
n-alkane) to reconstruct the delivery of sedimentary organic
matter in Mud Lake, Florida, over the past five millennia.
Originally a land-plant dominated marsh, the lake evolved into
a cyanobacteria-dominated system as the regional climate
became more humid and the local water table rose.
Compound-specific dD values
Because the hydrogen in most organic matter originates from
water, the D/H ratio in aquatic biomarker compounds is a sen-
sitive paleohydrologic proxy. The dD of lake water can be
reconstructed within 10% from the D/H ratios of sterols
derived from phytoplankton (Sauer et al., 2001). The D/H
ratios of the C
23
n-alkane that is a biomarker for Sphagnum
were used by Xie et al. (2000) to reconstruct a multi-century
paleoclimatic record for Bolton Fell Moss, Cumbria, England.
The dD values increase during times of warmer climate and
decrease when climate was cooler as rates of evaporative distil-
lation of bog waters increased or decreased. Similarly, Huang
et al. (2002) employed dD variations in n-hexadecanoic acid,
which is produced principally by lake algae, to reconstruct
the post-glacial temperature record preserved in the sediments
of Crooked Pond, Massachusetts.
Summary
Sediment organic matter provides a variety of elemental, iso-
topic, and molecular proxies to reconstruct past climates
and ancient environments. C/N ratios identify the general
origin of organic matter from aquatic or land plants. Bulk
and compound-specific d
13
C values distinguish proportions
of C
3
and C
4
plant organic matter and thereby identify
periods of dry climate. Nitrogen cycling is sensitive to various
environmental conditions that leave distinctive impacts on d
15
N
values. Biomarker molecules identify organic matter contribu-
tions from specific sources, which record different environmen-
tal settings, and they provide information about depositional
conditions from their alterations from precursor compounds.
Philip A. Meyers
Bibliography
Brenner, M., Whitmore, T.J., Curtis, J.H., Hodell, D.A., and Schelske, C.L.,
1999. Stable isotopes (
13
C&
15
N) signatures of sedimented organic
matter as indicators of historic lake trophic state. J. Paleolimnol., 22,
205–221.
Brincat, D., Yamada, K., Ishiwatari, R., Uemura, H., and Naraoka, H.,
2000. Molecular-isotopic stratigraphy of long-chain n-alkanes in Lake
Baikal Holocene and glacial age sediments. Org. Geochem., 31,
287–294.
Cranwell, P.A., Eglinton, G., and Robinson, N., 1987. Lipids of aquatic
organisms as potential contributors to lacustrine sediments – II. Org.
Geochem., 11, 513–527.
Filley, T.R., Freeman, K.H., Bianchi, T.S., Baskaran, M., Colarusso, L.A.,
and Hatcher, P.G., 2001. An isotopic biogeochemical assessment of
shifts in organic matter input to Holocene sediments from Mud Lake,
Florida. Org. Geochem., 32, 1153–1167.
Freeman, K.H., and Colarusso, L.A., 2001. Molecular and isotopic records
of C
4
grassland expansion in the late Miocene. Geochim. Cosmochim.
Acta, 65, 1439–1454.
Goossens, H., Duren, R.R., de Leeuw, J.W., and Schenck, P.A., 1989.
Lipids and their mode of occurrence in bacteria and sediments-II. Lipids
in the sediment of a stratified, freshwater lake. Org. Geochem., 14,
27–41.
Griffiths, M., 1978. Specific blue-green algal carotenoids in sediments of
Esthwaite water. Limnol. Oceanogr., 23, 777–784.
662 ORGANIC GEOCHEMICAL PROXIES

Hedges, J.I., and Mann, D.C., 1979. The characterization of plant tissues by
their lignin oxidation products. Geochim. Cosmochim. Acta, 43,
1803–1807.
Huang, W.-Y., and Meinschein, W.G., 1979. Sterols as ecological indica-
tors. Geochim. Cosmochim. Acta, 43, 739–745.
Huang, Y., Shuman, B., Wang, Y., and Webb, T., 2002. Hydrogen isotope
ratios of palmitic acid in lacustrine sediments record late Quaternary cli-
mate variations. Geology, 30, 1103–1106.
Meyers, P.A., and Lallier-Vergès, E., 1999. Lacustrine sedimentary organic
matter records of Late Quaternary paleoclimates. J. Paleolimnol., 21,
345–372.
Meyers, P.A., Leenheer, M.J., and Bourbonniere, R.A., 1995. Diagenesis of
vascular plant organic matter components during burial in lake sedi-
ments. Aquat. Geochem., 1,35–52.
Nishimura, M., and Koyama, T., 1977. The occurrence of stanols in various
living organisms and the behavior of sterols in contemporary sediments.
Geochim. Cosmochim. Acta, 41, 379–385.
Passier, H.F., Bosch, H.J., Nijenhuis, I.A., Lourens, L.J., Böttcher, M.E.,
Leenders, A., Sinninghe Damsté, J.S., de Lange, G.J., and de
Leeuw, J.W., 1999. Sulphidic Mediterranean surface waters during Plio-
cene sapropel formation. Nature, 397, 146–149.
Prahl, F.G., Muehlhasusen, L.A., and Lyle, M.A., 1989. An organic geo-
chemical assessment of oceanographic conditions at MANOP Site C
over the past 26,000 years. Paleoceanography, 4, 495–510.
Rau, G.H., Arthur, M.A., and Dean, W.E., 1987.
15
N/
14
N variations in
Cretaceous Atlantic sedimentary sequences; implication for past
changes in marine nitrogen biogeochemistry. Earth Planet. Sci. Lett.,
82, 269–279.
Rieley, G., Collier, R.J., Jones, D.M., and Eglinton, G., 1991. The biogeo-
chemistry of Ellesmere Lake, U.K. – I: source correlation of leaf wax
inputs to the sedimentary record. Org. Geochem., 17, 901–912.
Sauer, P.E., Eglinton, T.I., Hayes, J.M., Schimmelmann, A., and Sessions, A.L.,
2001. Compound-specific D/H ratios of lipid biomarkers from sediments as
a proxy for environmental and climatic conditions. Geochim. Cosmochim.
Acta, 65,213–222.
Schwark, L., Zink, K., and Lechterbeck, J., 2002. Reconstruction of post-
glacial to early Holocene vegetation history in terrestrial Central Europe
via cuticular lipid biomarkers and pollen records from lake sediments.
Geology, 30, 463–466.
Struck, U., Emeis, K.C., Voss, M., Krom, M.D., and Rau, G.H., 2001. Bio-
logical productivity during sapropel S5 formation in the Eastern Medi-
terranean Sea: Evidence from stable isotopes on nitrogen and carbon.
Geochim. Cosmochim. Acta, 65, 3249–3266.
Talbot, M.R., and Johannessen, T., 1992. A high resolution palaeoclimatic
record for the last 27,500 years in tropical West Africa from the carbon
and nitrogen isotopic composition of lacustrine organic matter. Earth
Planet. Sci. Lett., 110,23–37.
Twichell, S.C., Meyers, P.A., and Diester-Haass, L., 2002. Significance of
high C/ N ratios in organic-carbon-rich Neogene sediments under the
Benguela Current upwelling system. Org. Geochem., 33, 715–722.
Volkman, J.K., Barrett, S.M., and Blackburn, S.I., 1999. Eustigmatophyte
microalgae are potential sources of C
29
sterols, n-C
23
– n-C
28
n-alka-
nols and C
28
–C
32
n-alkyl diols in freshwater environments. Org. Geo-
chem., 30, 307–318.
Xie, S., Nott, C.J., Avsejs, L.A., Volders, F., Maddy, D., Chambers, F.M.,
Gledhill, A., Carter, J.F., and Evershed, R.P., 2000. Palaeoclimate
records in compound-specific dD values of a lipid biomarker in ombro-
trophic peat. Org. Geochem., 31, 1053–1057.
Zink, K.-G., Leythaueser, D., Melkonian, M., and Schwark, L., 2001. Tem-
perature dependency of long-chain alkenone distributions in Recent to
fossil limnic sediments and in lake waters. Geochim. Cosmochim. Acta,
65, 253–265.
Cross-references
Alkenones
Carbon Isotopes, Stable
Deuterium, Deuterium Excess
Geochemical Proxies (Non-Isotopic)
Nitrogen Isotopes
Paleoceanography
Paleoclimate Proxies, an Introduction
Paleolimnology
Sapropels
OSTRACODES*
Introduction
Ostracodes (also ostracods, Ostracoda) are a class of bivalved,
aquatic Crustacea that secrete a small (0.1 to > 2-mm long) cal-
citic shell (the carapace) that is commonly fossilized. Ostracodes
are used widely in paleoclimatology and the reconstruction of
marine and non-marine paleoenvironments because of their
small size, ecological sensitivity, well-known biology and shell
chemistry, long stratigraphic range (Ordovician-present), and
occurrence in sediments from lakes, estuaries, bays, and oceans.
There are an estimated 33,000 described living and extinct
ostracode species divided into two subclasses, the Myodocopa
and Podocopa, and 10 orders distinguished from one another on
the basis of appendages and carapace features such as size, shape,
muscle scar pattern, pore patterns, and hinge structure. Many
Myodocopids are pelagic in habitat; some are non-calcitic and
are rarely fossilized. The Podocopa includesthe order Podocopida,
(Figures O29 and O30), which are the most commonly fossilized
ostracode group and the most applicable to paleoclimatology.
Morphology and ecology
The carapace consists of two valves articulated along the dorsal
margin, encompasses the living animal, and forms part of the
organism’s cuticle, which is secreted by the epidermis. Ostra-
codes grow by molting (ecdysis), during which podocopids pro-
duce 8–9 molts or instars. Both adult and juvenile carapaces and
valves are commonly fossilized. Most ostracode genera and spe-
cies can be identified on the basis of external and internal cara-
pace features, although examination of soft parts is necessary
to identify species in some groups and recent genetic studies
have added considerably to understanding ostracode taxonomy.
Most species are free-living (either benthonic or pelagic),
move by crawling or swimming, reproduce sexually (a few are
parthenogenic), and disperse passively (there is no planktonic
larval stage). They inhabit a full range of aquatic environments
from small ephemeral pools of water, groundwater, to all types
of lakes, estuaries, bays, rivers, and oceanic environments. Spe-
cies are adapted to specific microhabitat conditions and physical
and chemical factors controlling species distribution include
temperature, salinity, light availability, solute chemistry, alkali-
nity, oxygen and carbon dioxide levels, and food resources.
Application to paleoclimatology and
paleoenvironments
The ecological sensitivity of living species has made ostracodes
useful for paleoenvironmental reconstruction since the nine-
teenth Century, especially in studies of Quaternary sediments
where living species are often found as fossils. Quantitative
analyses of faunal assemblages and indicator species have
been used extensively to reconstruct water solute composition
and regional atmospheric conditions in lakes, salinity and sub-
merged aquatic vegetation in estuaries, ocean temperatures on
continental shelves, dissolved oxygen on continental slopes,
and deep water masses in abyssal environments.
During the past decade, the application of ostracode shell
chemistry to contemporary issues surrounding climate varia-
bility has accelerated rapidly, especially in paleolimnology
*All rights reserved
OSTRACODES 663
because ostracodes are usually the only calcitic microfossil
group. With greatly improved understanding of the factors that
control the trace element (magnesium/calcium, strontium/
calcium ratios) and stable isotopic (oxygen and carbon) com-
position of the carapace, ostracode researchers have been able
to quantify temporal changes in lake and ocean temperature,
atmospheric temperature and precipitation, estuarine and bay
salinity, and other climate-related parameters (see review
papers in Holmes and Chivas, 2002). For example, in semi-arid
to subhumid, hydrologically-closed lakes, precipitation
influences the salinity, solute chemistry and
18
O/
16
O ratios of
the water. Chivas et al. (1993) applied these relationships to
infer salinity and evaporation changes in Lake Keilambete,
Australia during the late Quaternary using a d
18
O isotopic
record and Sr/Ca ratios for several ostracode species. In deep
lakes, ostracode shell chemistry can provide a record of atmo-
spheric temperature. Von Grafenstein et al. (1999) recon-
structed decadal scale changes in atmospheric temperature
from the d
18
O record of Candona from the Ammersee,
Germany for the important glacial, deglacial and Holocene
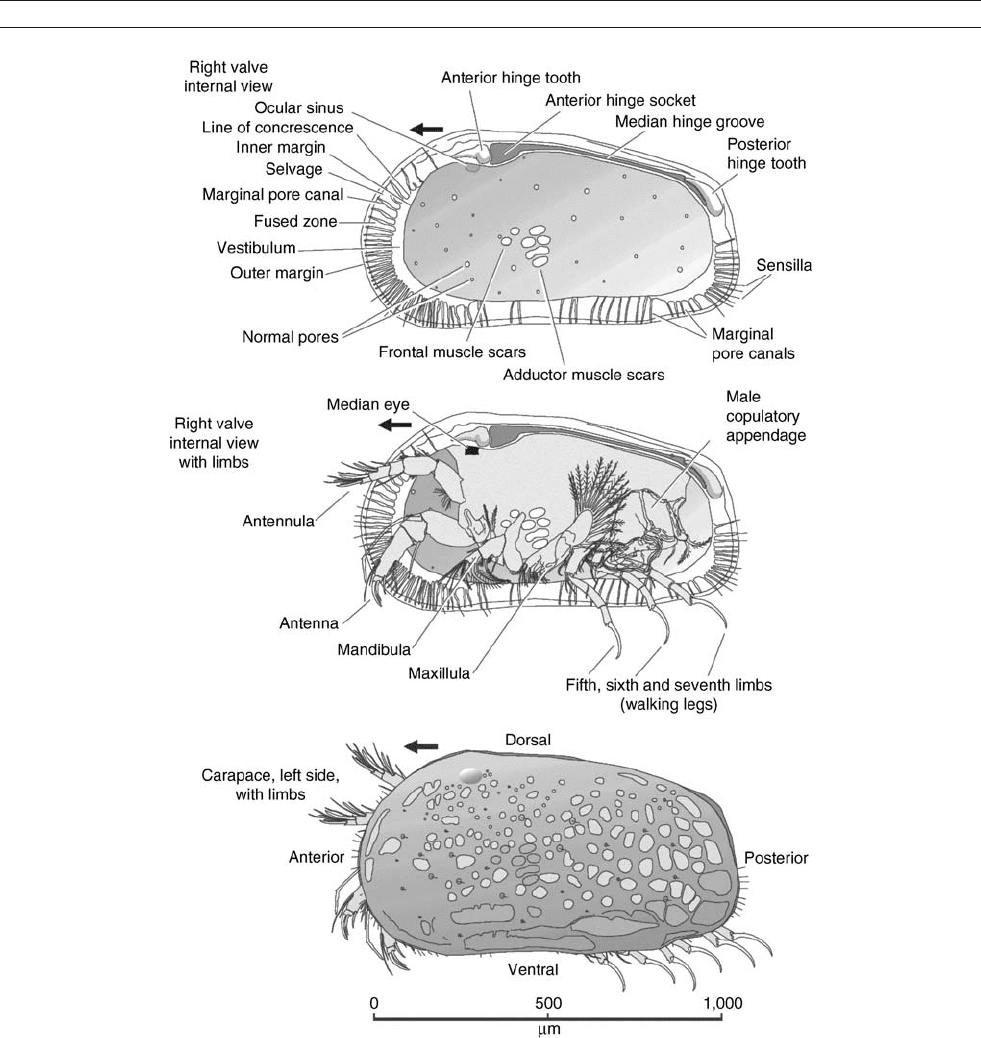
Figure O29 Morphology of Podocopid ostracode Hemicythere villosa (courtesy of David J. Horne).
664 OSTRACODES

interval, and showed a remarkable correspondence to the paleo-
temperature record from Greenland ice cores. Wansard (1996)
also reconstructed summer temperatures from Lake Banyoles,
Spain for the interval 30,000–6,000 years ago using Mg/Ca
ratios in Cyprideis.
Ostracode shell chemistry has also been applied to shallow
water paleoclimate records. In the Gulf of Carpentaria, northern
Australia, DeDeckker et al. (1988) reconstructed the late Qua-
ternary salinity changes caused by changing sea level from
Mg/Ca, Sr/Ca and
87
Sr/
86
Sr ratios of Cyprideis. Dwyer et al.
(2001) showed that salinity influences Mg/Ca ratios in Florida
Bay, which allowed them to reconstruct paleosalinity history
controlled by climate processes such as the El Nino-Southern
Oscillation. In Chesapeake Bay, eastern North America, water
temperature controls the Mg/Ca ratios in Loxoconcha and
Cronin et al. (2003) reconstructed a 2,200-year bay temperature
record. In the deep sea, studies by Dwyer et al. (1995, 2000),
Cronin et al. (2000), and Didié and Bauch (2002) provide
evidence from ostraocde shell chemistry for Pliocene and
Quaternary variability in deep-sea temperatures, associating
them with orbital and suborbital control of deep-water formation.
Thomas M. Cronin
Bibliography
Chivas, A.R., De Deckker, P., Cali, J.A., Chapman, A., Kiss, E., and
Shelley, J.M.G., 1993. Coupled stable isotope and trace-element mea-
surements of lacustrine carbonates as paleoclimatic indicators. In Swart,
P.K., Lohmann, K.C., McKenzie, J., and Savin S. (eds.), Climate
Change in Continental Isotopic Records, Washington, USA: American
Geophysical Union, pp. 113–121.
Cronin, T.M., Dwyer, G.S., Baker, P.A., Rodriguez-Lazaro, J., and
DeMartino, D.M., 2000. Orbital and suborbital variability in North
Atlantic bottom water temperature obtained from deep-sea ostracode
Mg/ Ca ratios. Palaeogeogr. Palaeoecol. Palaeoclimatol., 162,45–57.
Cronin, T.M., Dwyer, G.S., Kamiya, T., Schwede, S., and Willard, D.A.,
2003. Medieval warm period, Little Ice Age and 20th century temp-
erature variability from Chesapeake Bay. Glob. Planet. Change, 36
(1–2), 17–29.
DeDeckker, P., Chivas, A.R., Shelley, J.M.G., and Torgersen, T., 1988.
Ostracod shell chemistry: A new palaeoenvironmental indicator applied
to a regressive/ transgressive record from the Gulf of Carpentaria,
Australia. Palaeogeogr. Palaeoclimatol. Palaeoecol., 66, 231–241.
Didié, C., and Bauch, H.A., 2002. Implications of Upper Quaternary stable
isotope records of marine ostracodes and benthic foraminifers for
paleoecological and paleoceanographical investigations. In Holmes, J.A.,
and Chivas, A.R. (eds.), The Ostracoda: Applications in Quaternary
Research. Washington, USA: American Geophysical Union, pp. 279–299.
Dwyer, G.S., and Cronin, T.M., 2001. Ostracode shell chemistry as a paleo-
salinity proxy in Florida Bay. In Wardlaw, B. (ed.), Bulletins of
American Paleontology No. 361, pp. 249–276.
Dwyer, G.S., Cronin, T.M., Baker, P.A., Raymo, M.E., Buzas, J., and
Corrège, T., 1995. Late Pliocene and Quaternary bottom water
temperature change in the deep North Atlantic. Science, 270,
1347–1351.
Dwyer, G.S., Cronin, T.M., Baker, P.A., and Rodriguez-Lazaro, J., 2000.
Changes in North Atlantic deep-sea temperature during climatic fluctua-
tions of the last 25,000 years based on ostracode Mg/Ca ratios.
Geochem. Geophys. Geosyst., 1(12): (doi:10.1029/ 2000GC000046).
Holmes, J.A., and Chivas, A.R. (eds.), 2002. The Ostracoda: Applications
in Quaternary Research. Washington, USA: American Geophysical
Union, 313pp.
Von Grafenstein, U., Erlenkeuser, H., Brauer, A., Jouzel, J., and Johnsen. S.J.,
1999. A mid-European decadal isotope-climate record from 15,500 to
5,000 years
BP. Science, 284,1654–1657.
Wansard, G., 1996. Quantificaton of paleotemperature changes during
isotopic stage 2 in the La Draga continental sequence (NE Spain)
based on the Mg /Ca ratio of freshwater ostracods. Quaternary Sci.
Rev., 15, 237–245.
Cross-references
Geochemical Proxies (non-isotopic)
Oxygen Isotopes
Paleolimnology
Paleotemperatures and Proxy Reconstructions
Strontium Isotopes
OUTWASH PLAINS
Outwash plains are extensive areas of glaciofluvial sediments
deposited in the proglacial zone. They occur in topographically
unconfined locations, such as the margins of ice sheets or pied-
mont glaciers, distinct from the valley trains or valley bottom
deposits that occur in front of valley glaciers in confined settings.
The Icelandic term “Sandur” (plural: Sandar) is often used.
Sediment is deposited primarily by braided meltwater streams
that migrate across the surface of the outwash plain. The whole
plain is rarely covered all at onceexcept during extremejökulhlaup
floods. During floods, previously deposited sediments may be
reworked and redeposited. Outwash deposits can retain sedimen-
tological signatures of long periods of discharge from the glacier,
making it possible to judge the relative significance of ablation
dominated discharge and discharge related to extraordinary events
such as subglacial volcanic eruptions as in Iceland. Outwash
plains that are periodically inundated by floods exhibit distinctive
sediment structures.
Outwash plains can aggrade to thicknesses of several hundred
meters and bury the snout of the glacier from which the meltwater
and sediment are derived. Melting of buried ice can then lead to
the formation of kettles and pitted outwash surfaces, and rapid
retreat of the ice margin can leave a distinctive ice-contact
slope at the head of the outwash plain. Fluctuations in meltwater
discharge or supply of sediment can cause changes in the rate
of aggradation, and incision can occur creating terraces in the
outwash surface.
Close to the glacier, sediments are often coarse grained and
poorly sorted, with sub-angular clasts. Farther from the glacier
sediments are characteristically finer grained and better sorted
with more rounded clasts.
Figure O30 Scanning electron photo of deep sea ostracode Bradleya.
Left valve, 1.0 mm long.
OUTWASH PLAINS 665
Sedimentary structures within the outwash can reveal details
of flow regimes and sedimentary processes,can help to reconstruct
paleodischarges, and can help to unravel the dynamic history of
the glacier from which they are derived Benn and Erans (1998)
provide a useful review of this topic.
Peter G. Knight
Bibliography
Benn, D.I., and Evans, D.J.A., 1998. Glaciers and Glaciation. London,
UK: Arnold, 734pp.
Cross-references
Glacial Geomorphology
Glacial Sediments
Kettles
OXYGEN ISOTOPES
Natural occurrence
Oxygen has three stable isotopes with atomic mass numbers
of 16, 17, and 18 (
16
O,
17
O and
18
O), which occur naturally in
relative proportions of 99.76%, 0.04%, and 0.2%, respectively.
Because
17
O and
18
O fractionate (to a first order) proportionally
with respect to
16
O, and because of the very small abundance of
17
O, the isotopic ratio
18
O/
16
O(1/500) is most commonly
measured in water (e.g., groundwater, ice), carbonate sediments
(CaCO
3
), oxygen gas (O
2
), carbon dioxide (CO
2
), and organic
matter, in which the
18
O/
16
O ratio varies by more than 100%.
The
18
O/
16
O ratio of a sample is measured by mass spec-
trometry on CO
2
or O
2
. The isotopic composition is usually
expressed relative to a standard (which depends on the mea-
sured material) and expressed with the d notation:
d
18
O ¼
ð
18
O=
16
OÞ
Sample
ð
18
O=
16
OÞ
Standard
1
"#
ð1;000; per milÞð1Þ
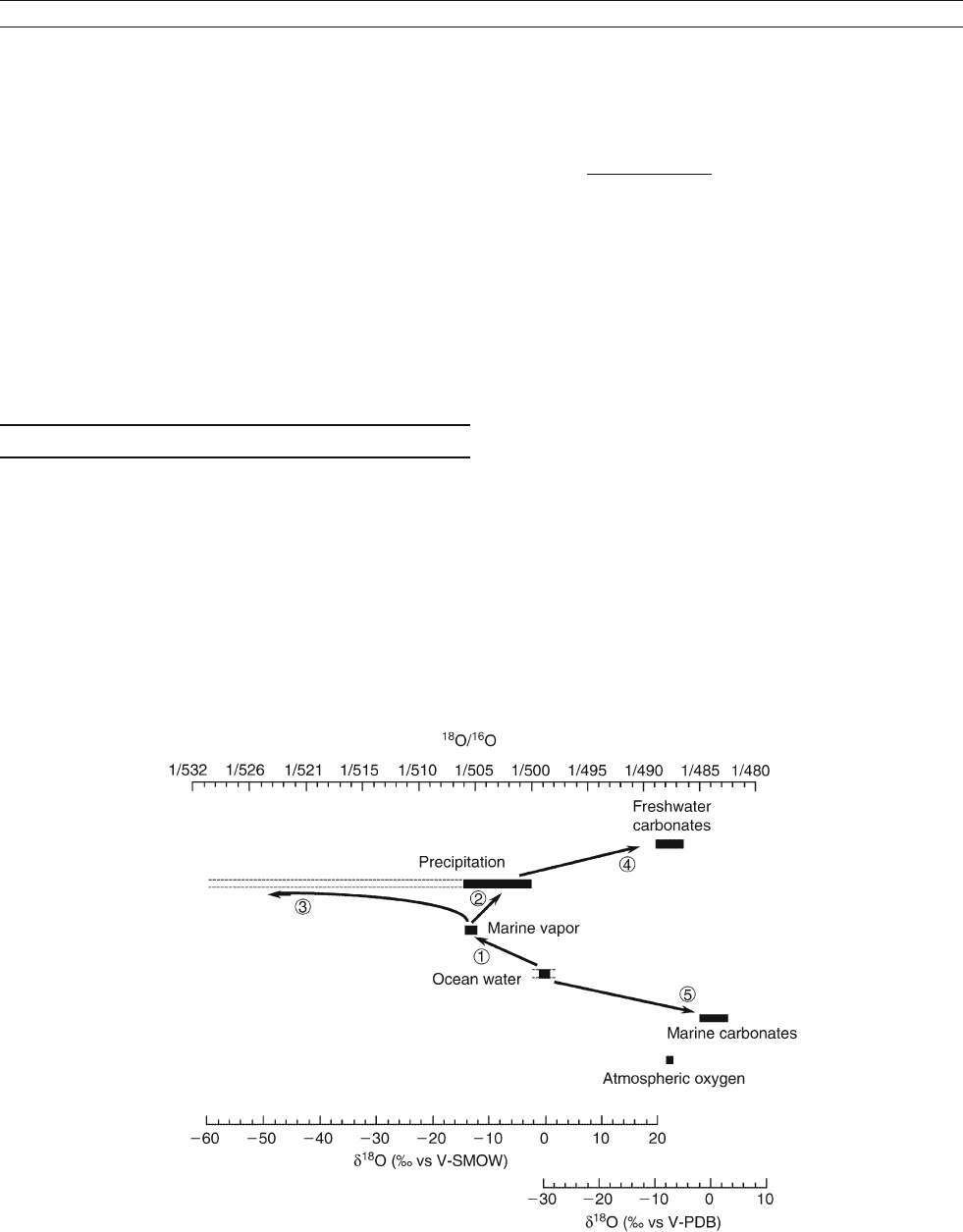
Atmospheric fractionation and local temperature
control of th e isotopic composition
Isotopic fractionation occurs during water evaporation and con-
densation (processes 1, 2 and 3 in Figure O31). At equilibrium,
the
18
O/
16
O ratio in the condensed phase (liquid or solid) is
higher by approximately 10 per mil as compared to the vapor
phase. In the hydrological cycle, water is mostly fractionated
in the atmosphere during the net transport of vapor between
low to high latitudes, ocean to continents, or low to high eleva-
tions, as a result of successive condensation–evaporation stages
(Dansgaard, 1964). At each stage, the isotopic enrichment of
the condensate depletes the remaining vapor in
18
O as com-
pared to
16
O(Figure O32, and process 3 in Figure O31), and
this results in more depleted precipitation at the next con-
densation stage. In this process of distillation, the isotopic
depletion of the vapor is primarily related to its exhaustion
(quantity of remaining vapor compared to the initial quantity).
Multivariate analyses of the
18
O/
16
O ratio in modern precipita-
tion have shown that the main parameters controlling this
depletion are the local temperature, the quantity of precipita-
tion, the distance to the coast, and the elevation (Figure O33).
The corresponding effects have been coined “temperature
effect,”“amount effect,”“continental effect,” and “altitude
Figure O31 Range of the isotopic composition of oxygen at the surface, and relationships between the different compositions.
666 OXYGEN ISOTOPES
effect.” This isotopic depletion in the atmosphere determines
the composition of the water reservoirs on the Earth, from
the ocean to the cryosphere. See Mook ( 2001) for a thorough
exposition of the environmental isotopes.
When minerals like carbonate, phosphate, and silica, which
constitute sediments, form in water, their
18
O/
16
O ratios are
fractionated with respect to the water ratio, a fractionation
mainly controlled by temperature (Epstein et al., 1953). Hence,
fossil water, mainly ice and groundwater, as well as sedi-
ments, represent potential archives of information regarding
past climates and environments.
Precipitation and the isotopic thermometer
The first primary standard for water was the Standard Mean
Ocean Water (SMOW; Craig, 1961a,b). Because of problems
with its definition, the International Atomic Energy Agency
(IAEA) defined the Vienna Standard Mean Ocean Water
(V-SMOW), with an isotopic composition very close to
SMOW, and prepared in a large amount such that it is still
distributed to laboratories today. Its absolute
18
O/
16
O atomic
ratio is 2,005.20 ( 0.45) 10
6
(Baertschi, 1976). This com-
position has been chosen to be very close to the oceanic
composition. As such, most precipitation has a much lower
18
O/
16
O ratio (Figure O31 and O33), and secondary, more
depleted, standards are used for their measurements. IAEA
now recommends that isotopic measurements on water be
expressed with respect to V-SMOW, and normalized on a
two-standard scale, V-SMOW and SLAP (Standard Ligh t
Antarctic Precipitation), assuming a value d
18
O ¼55.5%
for S LAP ( Coplen, 1995).
The usual measurement technique is to equilibrate a small
amount of CO
2
gas with each water sample, and to determine
the isotopic composition of this CO
2
by mass spectrometry.
This technique requires only a few milliliters of water. The
Figure O32 Isotopic evolution of a vapor mass, modeled as a
Rayleigh-type distillation: an initial vapor mass is exhausted by
successive condensation stages, for instance during an advection to
higher latitude. At each stage, the
18
O/
16
O ratio of the condensate is
enriched by 10% compared to the vapor ratio, which depletes the
remaining vapor. F is the fraction of remaining vapor.
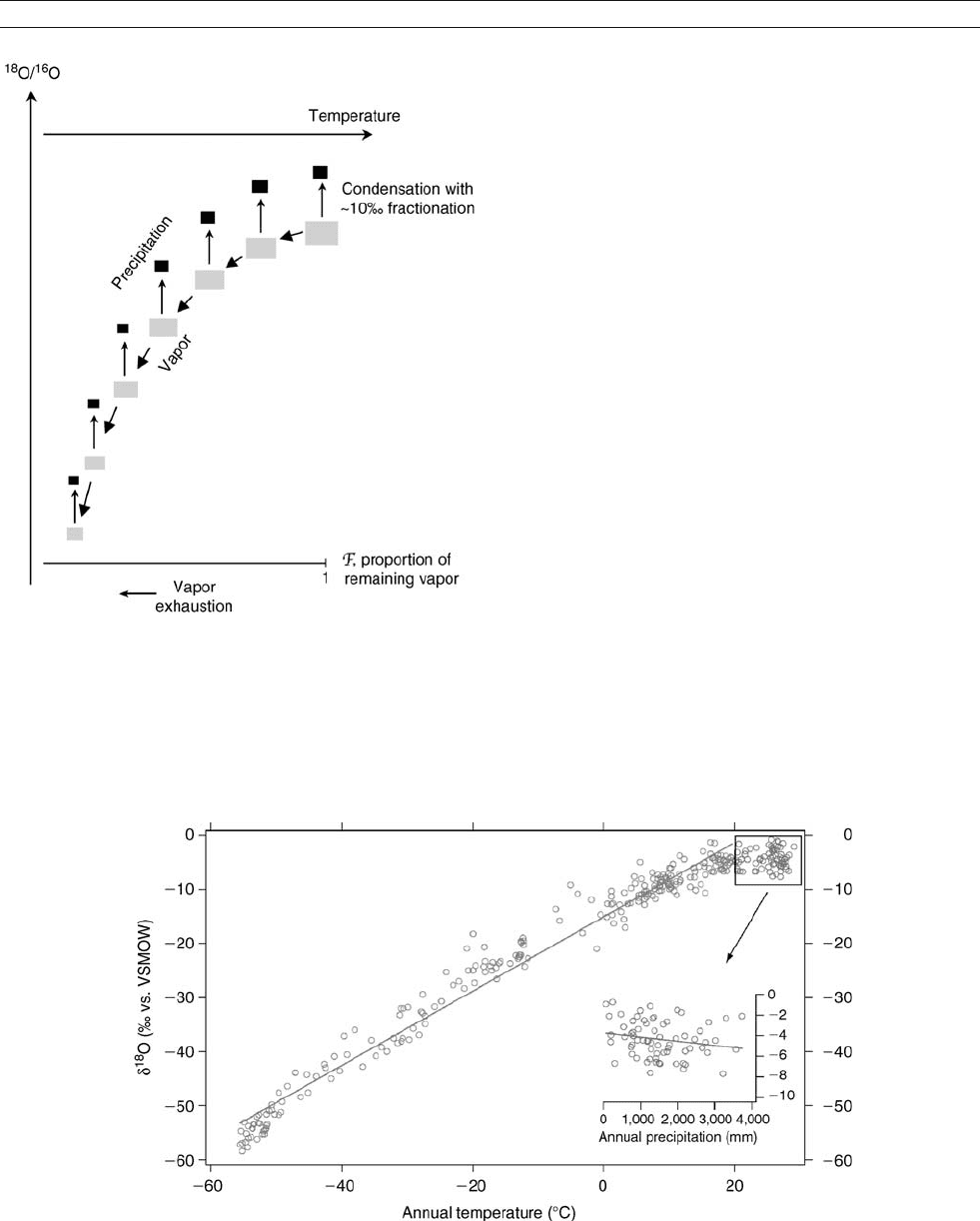
Figure O33 d
18
O – surface temperature relationship for modern precipitation (annual averages) (data from the GNIP network Rozanski et al., 1993),
augmented by some polar data (cf. Figure O34). The linear correlation is calculated for points with an annual temperature <20
C(N = 243 points,
r = 0.987). The insert shows the annual d
18
O – precipitation relationship (“amount effect”) for stations with annual temperature >20
C.
OXYGEN ISOTOPES 667
sample composition is measured with respect to a reference sam-
ple, which has been previously calibrated against V-SMOW.
Both samples are measured under the same conditions, so that
a correction for the gas–water fractionation does not have to
be applied. This technique usually achieves a precision better
than 0.1%, but with lower accuracy, as shown from interlabora-
tory comparisons organized by IAEA. Other techniques have
been developed, like water hydrolysis and laser spectrometry.
The isotopic composition of precipitation is imprinted, more
or less directly, in many continental records like glacier ice,
snow, groundwaters, tree rings, freshwater carbonates, spe-
leothems, etc. The discussion below focuses on ice, but may
be extended in a similar way to the other archives (see below).
Isotopic measurements on precipitation have been shown
to correlate strongly with surface temperature at mid-to-high
latitudes, and with the amount of precipitation at low latitudes,
the so-called “amount effect” (Figure O33 ; Dansgaard, 1964;
Rozanski et al., 1993). The isotope-temperature relationship
can be described with the vapor exhaustion, as shown by a sim-
ple distillation model (batch distillation, also called “Rayleigh
distillation”: Figure O32; Dansgaard, 1964). The modeling of
this relationship can be improved by considering other second-
ary parameters, like the proportion of different water phases in
the clouds (vapor, liquid, and solid water), evaporation of
the falling droplets (especially for the “amount effect”); and
kinetic fractionation during the formation of snow (Jouzel
and Merlivat, 1984).
The observed correlation between d
18
O and temperature
(Figure O33) is strongest for annual means, and in polar
regions (Figure O34). It is based on different stations, and thus
a spatial gradient Dd
18
O/DT is defined, where D is a variation
over space. This gradient is traditionally used to interpret isoto-
pic variations (e.g., in the ice) in terms of past temperature, and
has been called the “isotopic paleothermometer” (Figure O34).
Yet, a temporal gradient Dd
18
O/DT must be used to interpret
past isotopic variations, where D is a variation, at one location,
over the timescale of the considered climatic variations. In recent
years, estimates of temperature variations in Central Greenland
have been achieved independently of d
18
O. They have shown
that, over the last glacial period, the isotopic paleothermometer
underestimates past temperature variations by a factor of two;
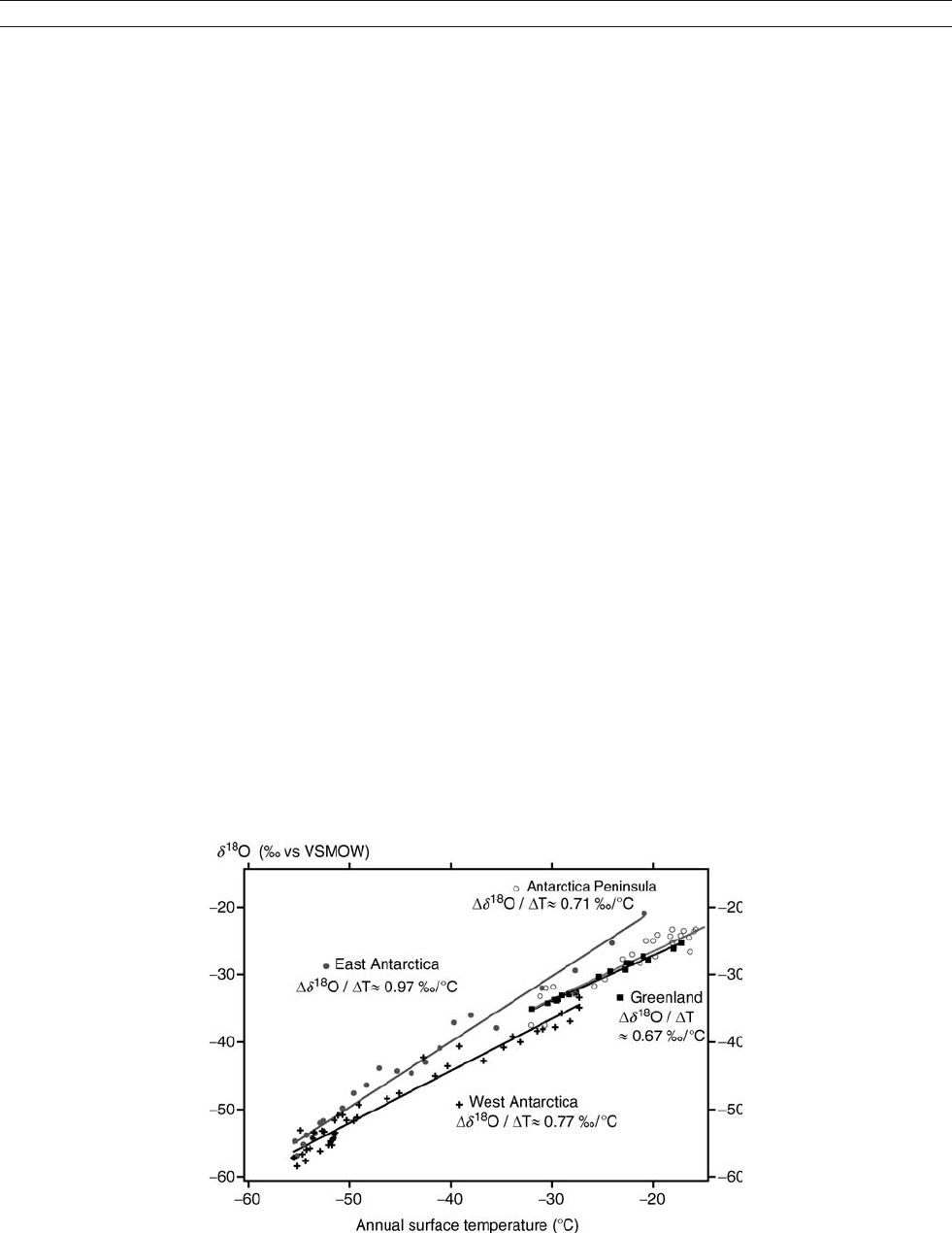
that is, the temporal gradient is about half the spatial one (Cuffey
et al., 1995; Jouzel, 1999). So far, there is no evidence for such
a strong bias in East Antarctica records (Jouzel et al., 2003).
The difficulty in interpreting past d
18
O variations thus consists
in determining the relevant temporal d
18
O–temperature gradient,
because a number of climatic parameters can cause this gradient
to differ from the spatial gradient. Some of these parameters
are briefly described below; a thorough review can be found in
Jouzel et al. (1997).
The origin of the temperature-isotope relationship in preci-
pitation has to be determined for each case. The d
18
Oof
the condensate is determined by both the vapor d
18
O and the
local temperature at the condensation level, T
c
. Over the year,
temperatures at the surface (T
s
) and at the condensation level
(T
c
) may well correlate during precipitation events, parti-
cularly in polar regions. For Antarctica, a linear relationship
T
c
¼ 0.67 T
s
– 1.2 has been obse rved (J ouz el and Merl ivat,
1984). This justifies the use of the surface temperature T
s
,
rather than T
c
, in t he paleothermometer, and alternatively
may help explain some bias of this thermometer if the correla-
tion between Ts and Tc has not held through climatic changes.
The d
18
O of the vapor, before condensation, mainly depends
on the fraction of remaining vapor (F in Figure O32). To a
first order approximation, as shown by simple modeling,
this fraction is somehow determined by the difference between
the temperature of the remote vapor source and the tempera-
ture at the location of precipitation. This is so because the
temperature governs the quantity of vapor the air can hold
(specific humidity). Thus, interpreting a d
18
O record in terms
of local temperature changes only requires that the remote
(source) temperature has remained constant in time. For
instance, Aristarain et al. (1986) analyzed snow from the
Antarctic Peninsula and showed that, given that the precipita-
tion originates in the Southern Ocean, a covariation of the local
and oceanic temperatures can explain a temporal gradient
Figure O34 Isotopic paleothermometer for polar stations: data from Dahe et al. (1994) for Antarctica, and Johnsen et al. (1989) for Greenland.
668 OXYGEN ISOTOPES

Dd
18
O/DT lower by a factor of two than that of the observed
spatial slope. A way to check the temperature stability of the
precipitation source is provided by the deuterium-excess, a
second-order isotopic parameter measured in precipitation
(Jouzel and Merlivat, 1984). To a first order, the isotopic
composition of hydrogen in the water (dD, see Deuterium,
deuterium excess) is linearly related to the composition of
oxygen, d
18
O, because both vary similarly with temperature;
thus, combining d
18
O and dD gives access to other climatic
parameters.
It is also important to consider that each isotopic measure
from an archive results from the accumulation of many pre-
cipitation events, and is thus a weighted average, usually
over several months or years. If the accumulation is not evenly
distributed over the year, the temperature record may be
biased towards one particular season. For Central Greenland,
modeling studies have shown that the contribution of the warm
season to the yearly accumulation may have been more impor-
tant during glacial periods than today (Krinner et al., 1997).
This would bias the glacial d
18
O signal towards warmer
temperature, and may explain why the temporal gradient
Dd
18
O/DT is lower than the spatial gradient (cf. supra).
At low latitudes, the d
18
O depends less on temperature
and more on the quantity of precipitation (Figure O33), which
is called the “amount effect.” The isotopic signal may be
interpreted in terms of hydrologic changes, especially in the
monsoon area.
The isotopic record of carbonate sediments
Carbonate isotopic fractionation and the
temperature scale
Carbonate is a widely distributed sediment, usually of biogenic
origin, and for this reason it is also the most frequently used to
retrieve a climatic signal routinely from the
18
O composition.
The climatic interpretation of the carbonate d
18
O signal is based
on the pioneering work done by Harold C. Urey, Samuel
Epstein, Cesare Emiliani, and their colleagues at the University
of Chicago. They developed the concept of paleothermometry,
and defined the first isotopic reference, a belemnite from the
Peedee formation in South Carolina, called PDB. The isotopic
ratio is measured with a mass spectrometer directly on the
CO
2
degassed by reaction with pure H
3
PO
4
(McCrea, 1950).
For PDB, this isotopic ratio is 2,067.2 10
6
(Figure O31 ).
From measurements of the isotopic fractionation between water
and carbonate, they demonstrated the potential of fossil carbo-
nate to record environmental parameters (Urey, 1947 ; McCrea,
1950), especially temperature: newly formed carbonate records
the isotopic composition of the water, accounting for an isoto-
pic fractionation that is temperature-dependent. Epstein et al.
(1953) calibrated a temperature equation on living mollusks,
based on the isotopic composition of the carbonate (d
c
) and
of the water (d
w
):
t ¼ 16:5 4:3 ðd
c
d
w
Þþ0:14 ðd
c
d
w
Þ
2
ð2Þ
with t in
C and the d in per mil with respect to VPDB. Apply-
ing this equation to a long marine record, Emiliani (1955 )
pioneered the use of d
18
O in paleoceanography. Due to a short-
age in PDB, IAEA defined the Vienna Peedee Belemnite
(V-PDB) reference with respect to the standard of the National
Bureau of Standard NBS-19 as: d
18
O
NBS19 / VPDB
¼2.2%.
The reference material currently distributed to laboratories is
NBS-19.
Isotopic records and stratigraphy for the last
million years
The paleotemperature equation (Equation 2) is applied to carbo-
nate shells secreted by planktonic or benthic micro-organisms
like foraminifera, coccolithophoridae, which are found in mar-
ine sediments retrieved by deep sea coring. It is also applied to
other organic carbonate found in marine as well as freshwater
environments, like corals, mollusks, etc. (Figure O31, pro-
cesses 4 and 5), and to inorganic carbonates like speleothems.
Comparison between inorganically and organically precipitated
calcite has shown a fair agreement for the isotopic fractiona-
tion of oxygen, suggesting that, to a first order, organic carbo-
nate precipitates close to equilibrium with the surrounding
water. Yet, the temperature dependence of this fractionation
(Equation 2) has been recalibrated several times on different
species to account for small departures from equilibrium
(Wefer and Berger, 1991; Bemis et al., 1998). This temperature
dependence is approximately 0.25 per mil of d
c
per
C, or 4
C
per mil of d
c
. Equation 2 also shows that, for a constant tempera-
ture, a 1 per mil variation of d
w
is equivalent to a 4
C change.
Over the last million years, which have been dominated by
glacial cycles, both temperature and sea water composition
have changed, with similar effects on the isotopic record.
The marine glacial cooling of a few degrees has increased
fractionation by around 1 per mil, while stocking up water
in wide ice sheets has also enriched d
w
(and thus d
c
)by
1 per mil. This amplification of global climatic changes
explains the similarity of isotopic records retrieved from dif-
ferent oceanic regions, which underlies the so-called oxygen
isotope stratigraphy: periodic d
18
O up-and-downs are matched
between records and numbered, with odd and even num-
bers corresponding to warm and cold periods, respectively
(Figure O35). The SPECMAP group (Imbrie et al., 1984)
compiled a global d
18
O signal by retaining the most signifi-
cant excursions of different records. The final objective of
SPECMAP was to produce a common temporal framework
in order to date other records. The absolute dating of this fra-
mework has been a difficult task. Magnetic reversals detected
in the sediments are widely used, the youngest one being the
Brunhes-Matuyama about 780,000 years ago. Sea level mar-
kers like coral terraces are also used, since sea level variations
are imprinted in the marine isotopic signal. Spectral analysis
of the isotopic records have proved their origin in the varia-
tions of the Earth orbit (Hays et al., 1976), confirming the
astronomical theory of glacial cycles. A dating technique
derived from this confirmation, called “orbital tuning,” con-
sists in tuning the age of a record to the orbital configura-
tions, assuming some relationships between them.
The marine d
18
O signal: temperature and ice volume
The marine carbonates record both variations of water tem-
perature and of water d
18
O. The latter is conceptually divided
into a global and a local component. The global component
affects the quantity of
18
O of the ocean as a whole, on a
timescale longer than the ocean mixing time so that it is
expected to be homogeneous. This is due to fluctuations of
the ice sheet volume, which extract or release isotopically
depleted water from or to the oceans (Figure O31). On time-
scales longer than hundred million years, an imbalance in the
18
O oceanic budget is also possible (Lécuyer and Allemand,
1999). The local component affects the spatial distribution
of
18
O. For the surface, this is mainly due to hydrological
OXYGEN ISOTOPES 669