Тіманюк В.О., Животова О.М. Біофізика
Подождите немного. Документ загружается.

121
б)
в)
г)
д)
5.4. Електродифузійне рівняння Нернста — Планка має вигляд:
a)
б)
в)
г)
д)
5.5. Рівняння Гольдмана має вигляд:
а)
б)
в)
г)
д)
5.6. Рівняння Нернста має вигляд:
122
а)
б)
в)
г)
д)
5.7. Зміна вільної енергії при проникненні одного моля іонів із
розчину з діелектричною проникністю ε
в
у мембрану з діелектричною
проникністю ε
м
складає:
а)
б)
в)
г)
д)
5.8. У порівнянні з простою дифузією полегшена дифузія:
а) відбувається з меншою швидкістю;
б) відбувається з більшою швидкістю;
в) швидкість транспорту змінюється не за модулем, а за напрям-
ком;
г) швидкість транспорту не змінюється.
5.9. Na
+
,K
+
Fнасос транспортує:
а) у клітину 2Na
+
, із клітини 3К
+
;
б) у клітину 3Na
+
, із клітини 3К
+
;
в) у клітину 3K
+
, із клітини 2Na
+
;
г) у клітину 3Na
+
, із клітини 2К
+
;
д) у клітину 2K
+
, із клітини 3Na
+
.

123
5.10. Молекула валіноміцину транспортує через мембрану:
а) К
+
і Na
+
;
б) переважно Na
+
;
в) переважно K
+
;
г) К
+
і Са
2+
;
д) Н
+
і Cl
–
.
5.11. Напівпроникна мембрана розділяє два розчини одновалент них
іонів концентрацією 50 ммоль/мл і 500 ммоль/мл. Знайти різницю по-
тенціалів, що встановлюється на мембрані, при темпера турі 37 °С:
а) 30 мВ;
б) 97,8 мВ;
в) 200 мВ;
г) 50 В;
д) 61,5 мВ.

124
125
Розділ 6. БІОФІЗИКА НЕРВОВОГО ІМПУЛЬСУ
Нормальне функціонування організму неможливе без обміну інфор-
мацією між клітинами, одним із способів якого є можливість генерації і
сприйняття клітинами нервового імпульсу. У організмі існують так звані
збудливі клітини, до яких відносяться м’язові, нер вові і секреторні. Ці
клітини здатні реагувати якимFнебудь способом на їхнє збудження. Так,
м’язові клітини скорочуються, секреторні виділяють речовини, а нервові
генерують електричні коливання (нервовий імпульс).
§ 31. Потенціал спокою
Звичайно в клітині існує трансмембранна різниця потенціалів, тобто
електричний потенціал всередині клітини відрізняється від зовніш-
нього. Ця різниця потенціалів називається потенціалом спокою. Якщо
зовнішній потенціал прийняти рівним нулю, то внутрішній складе від
–50 до –90 мВ, у залежності від виду клітини.
Причиною існування потенціалу спокою є різна концентрація іонів
K
+
, Na
+
і Cl
–
з обох боків клітинної мембрани (табл. 6.1). Допустимо, що
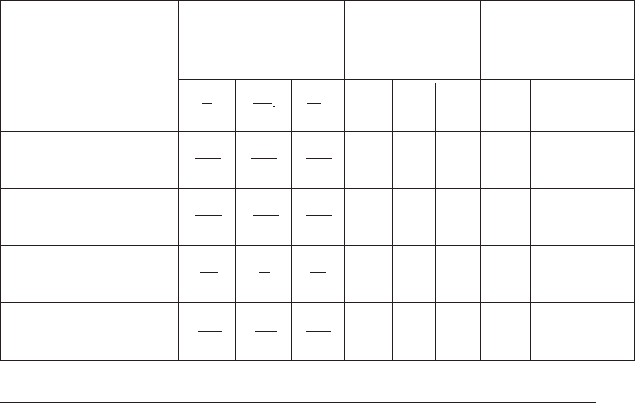
Табл. 6.1
Концентрація іонів K
+
, Na
+
і Cl
–
всередині і зовні клітини, значення по-
тенціалів спокою і потенціалів дії деяких клітин
Гігантський аксон 340 49 114 –88 +57 –42 –60 +50
каракатиці 10,4 463 592
Аксон 360 69 157 –90 +46 –29 –60 +35
кальмара 10,0 425 496
М’язове волокно 48 1 1 –98 +49
–
105
–88 +34
жаби 1 7 64
Моторний нейрон 150 15 9 –90 +60 –70 –90 +30
Клітини
Значення концентра-
цій іонів всередині (i) і
зовні (o) клітини, мМ
Рівноважний
потенціал для
різних іонів, мВ
Вимірені потенціа-
ли, мВ
на максимумі
потенціалу дії
К
i
+
К
о
+
Na
i
+
Na
о
+
Cl
i
+
Cl
о
+
спо-
кою
К
+
Na
+
Cl
+
126
мембрана проникна тільки для іонів калію і непроникна для інших, тоді
потенціал спокою буде визначатися рівноважним калієвим потенціалом
і описуватися рівнянням Нернста:
де R — універсальна газова стала; T — абсолютна температура;
F — число Фарадея; [K
+
]
o
— зовнішня концентрація іонів калію;
[K
+
]
i
— внутрішня. Якщо концентрація іонів калію всередині клітини
в 50 разів перевищує зовнішню, то потенціал спокою при температурі
27 °С складе —100,7 мВ.
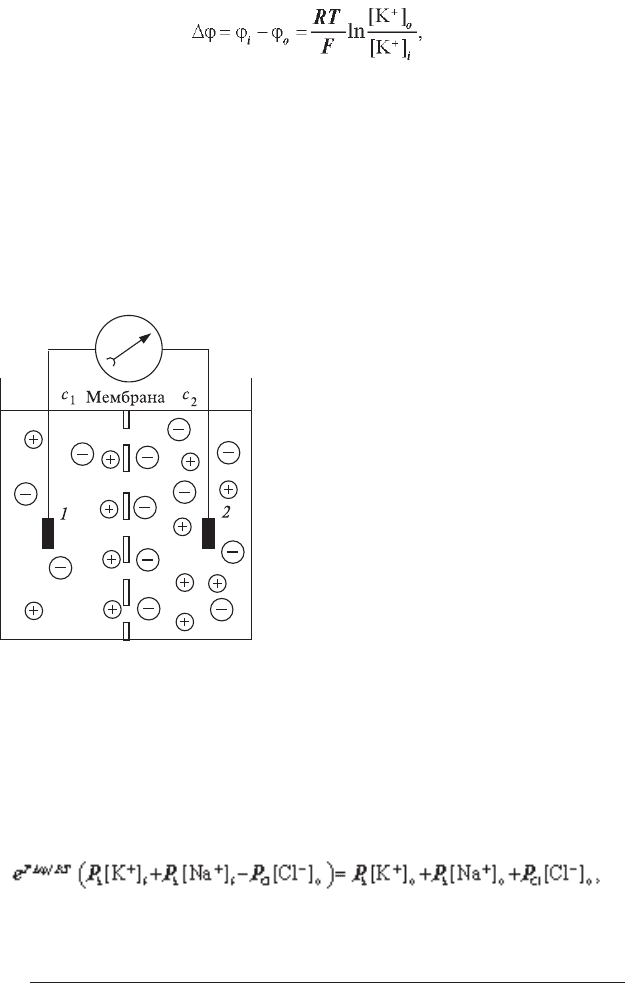
Як виникає потенціал спокою, легко зрозуміти з наступного експе-
рименту. Посудина розділена негативно зарядженою мембраною, з обох
боків якої знаходиться розчин тієї самої речовини, наприклад, К
2
SO
4
,
але різної концентрації (рис. 6.1). Позитивно заряджені іони калію по-
чинають дифундувати через мембрану
в розчин із меншою концентрацією,
приносячи туди свій позитивний за-
ряд. Негативні сульфатFіони, нездатні
пройти через мембрану, накопичуються
біля неї і електростатично утримують
іони калію, що знаходяться по інший
бік мем брани. У результаті виникає
трансмембранна різниця потенціалів.
Величина потенціалу спокою за-
вжди декілька нижча рівноважного
калієвого потенціалу, що пояснюється
просочуванням усередину клітини іонів
натрію, які приносять туди свій пози-
тивний заряд. Дифузія негативно за-
ряджених іонів хлору, навпаки, сприяє
збільшенню ∆ϕ. З цього випливає, що
на величину потенціалу спокою впли-
ває проникність клітинної мембрани
для різних іонів.
Так, якщо мембрана в спокої про-
никна не тільки для іонів калію, але і для інших, то внесок у формування
потенціалу спокою вносять потоки всіх цих іонів. У подальших розра-
хунках будемо враховувати транспорт тільки іонів K
+
, Na
+
і Cl
–
.
або
Рис. 6.1. Виникнення різниці по-
тенціалів на мембрані, що розділяє
розчини К
2
SO
4
концентрацій С
1
і С
2
.
1,2 — електроди; 3 — електровимірю-
вальний прилад.

127
У стаціонарному стані сума потоків іонів через мембрану дорівнює
нулю:
(6.1)
хоча потоки окремих іонів можуть бути і відмінні від нуля. Так як іони
хлору заряджені негативно, то перед потоком іонів хлору J
Cl
F
ставиться
знак «–».
Висловимо з рівняння Гольдмана (5.6) потоки для іонів K
+
, Na
+
та
Cl
–
з урахуванням їхнього заряду z:
і підставимо їх у рівняння (6.1). Скоротивши на загальний множник
запишемо
Розділимо чисельник і знаменник останнього доданку на множник
після чого одержимо

128
звідки остаточно одержуємо
Даний вираз називається рівнянням Гольдмана — Ходжкіна — Кат-
ца.
У стані спокою проникності для іонів K
+
, Na
+
і Cl
–
відносяться один
до одного як
P
K
: P
Na
: P
Cl
= 1 : 0,04 : 0,045.
Так як в стані спокою проникність клітини для іонів калію набагато
більша її проникності для інших іонів, то потенціал спокою визначається
переважно різницею концентрацій іонів калію.
Підтримка різниці концентрацій іонів здійснюється за допомогою
роботи іонних насосів, що використовують енергію АТФ. K
+
,Na
+
Fнасос,
крім того, сприяє збільшенню трансмембранного потенціалу, тому що
виводить три іони натрію і закачує у клітину тільки два іони калію, що
збільшує позитивний заряд міжклітинного середовища, а, отже, збіль-
шує потенціал спокою.
Збільшення трансмембранної різниці потенціалів називається
гіперполяризацією, зменшення — деполяризацією. У останньому
випадку може навіть змінитися знак внутрішньоклітинного потен-
ціалу.
Порушення роботи АТФази, яку можна викликати, наприклад, дією
серцевого глікозиду урбаніну, зменшує потенціал спокою. До такого ж
ефекту приводить дія різних отрут (наприклад, батрахотоксину), які
збільшують проникність клітинної мембрани для іонів натрію. Депо-
ляризацію викликає також гальмування процесів, які забезпечують у
клітині синтез АТФ.
§ 32. Потенціал дії
При впливі на клітину якогось подразника її трансмембранний
потенціал змінюється, виникає так званий потенціал дії, або спайк.
Причиною такого коливання потенціалу спокою є зміна проникності
мембрани для натрію, що, у свою чергу, викликано відкриттям натрієвих
іонних каналів. При збудженні мембранні проникності для іонів калію
і натрію відносяться як
P
K
: P
Na
= 1 : 20.
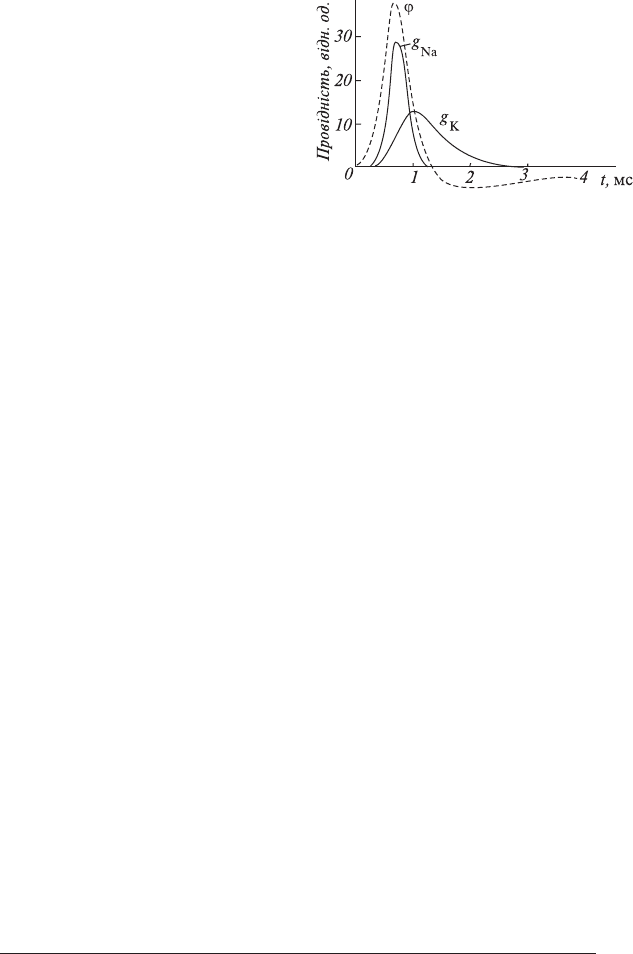
129
У результаті потік іонів нат рію в клітину починає перевищувати
потік іонів калію з клітини (рис. 6.2). Якщо раніше потенціал на мем-
брані був близький до рівноважного калієвого потенціалу, то тепер він
прагне до рівноважного натріє-
вого, який складає, наприклад,
для гігантського аксона кальмара
+46 мВ. І дійсно, мембранний
потенціал наближається до цієї
величини, але не досягає її тільки
внаслідок того, що проникності
для іонів калію і хлору не рівні
нулю. У клітині відбувається депо-
ляризація мембрани: негативний
потенціал клітини наближається
до нуля, а потім і зовсім змінює
знак на протилежний (рис. 6.3).
Останній процес називається ре-
версією мембранного потенціалу.
Максимальне значення потенціалу дії складає звичайно +30÷+40 мВ.
Збільшення натрієвої проникності триває частки мілісекунд. Далі вона
починає знижуватися, а калієва — зростати, у результаті чого в клітині
відновлюється потенціал спокою. Цей процес називається реполяри-
зацією клітинної мембрани.
Для відновлення потенціалу спокою клітині потрібен якийсь час.
У процесі реполяризації спочатку відбувається швидке наближення
до нормального значення потенціалу спокою, потім швидкість зміни
потенціалу зменшується, і клітина якийсь час перебуває в деполяри-
зованому стані. Це називається слідовою деполяризацією. У деяких
клітинах, навпаки, виникає слідова гіперполяризація, тобто в процесі
реполяризації різниця потенціалів починає перевищувати звичайну.
Обидва ці відхилення від нормального значення потенціалу спокою
називаються слідовими потенціалами (рис. 6.3).
Тривалість потенціалу дії відрізняється для різних клітин і коли-
вається в межах від 0,5 до 3 мс. Значний вплив на цю величину надає
температура. При її зменшенні на 10 °С час існування потенціалу дії
збільшується в три рази. Тривалість реполяризації звичайно перевищує
тривалість деполяризації.
Проникність мембрани для тих або інших іонів залежить від того,
відкритий або закритий канал для цього іона. Мембранний канал може
знаходитися тільки в одному із цих двох станів. Загальна провідність
мембрани залежить від співвідношення відкритих і закритих каналів.
Вони мають виборчу проникність і здатність активуватися або дезак-
тивуватися під дією змін трансмембранного потенціалу.
Рис. 6.2. Залежність провідності мембрани
для іонів натрію і калію (g
Na
і g
К
) від часу для
гігантського аксона кальмара під час генера-
ції потенціалу дії ϕ.

130
Дія подразника звичайно приводить до локальної деполяризації
мембрани. Це викликає відкриття натрієвих каналів, чутливих до зміни
потенціалу, а, отже, — збільшує натрієву провідність, що приводить до
ще більшої деполяризації. Цей процес сприяє відкриттю нових натрієвих
каналів. Існування такого зворотного зв’язку забезпечує регенеративну
або самообновлюючу деполяризацію клітинної мембрани.
Після виникнення потенціалу дії дана ділянка мембрани нервової
клітини якийсь час знаходиться в незбудливому (рефрактерному) стані,
тобто дія подразника не викликає генерацію потенціалу дії, тому що
натрієві канали якийсь час після активації знаходяться в закритому
стані і не здатні відкритися у відповідь на зміну трансмембранного
потенціалу.
При виникненні потенціалів дії в клітині зменшується концентрація
іонів калію і збільшується концентрація іонів натрію. Тому в цих умовах
необхідна активація мембранних насосів, що здійснюють активний
транспорт іонів. Так як для цього потрібні молекули АТФ, то в клітині
активуються процеси подиху, збільшується споживання кисню і теп-
лообмін.
§ 33. Подразнення мембрани електричним струмом
У експерименті збудження мембран частіше усього викликають
електричним струмом. Це дозволяє одержати багато кількісних харак-
теристик цього процесу. Одним із найбільш поширених методів є той,
при якому в досліджувану клітину поміщають анод, а катод залишають
у позаклітинному середовищі. Пропускання електричного струму
викликає деполяризацію мембрани, а, отже, — виникнення потенціалу
дії. При зворотному розташуванні електродів, коли анод залишають у
поверхні клітини, а катод поміщають всередину, потенціал дії не може
виникнути, тому що відбувається гіперполяризація мембрани.
Рис. 6.3. Фази потенціалу дії: 1 — фаза деполяризації; 2 — фаза реполяризації;
3 — слідовий потенціал: а) негативний слідовий потенціал; б) позитивний слідовий потен-
ціал.
б
а