Berg J.M., Tymoczko J.L., Stryer L. Biochemistry
Подождите немного. Документ загружается.


Note that 30 molecules of ATP are consumed per hexose molecule formed when the C
4
pathway delivers CO
2
to the
Calvin cycle, in contrast with 18 molecules of ATP per hexose molecule in the absence of the C
4
pathway. The high
concentration of CO
2
in the bundle-sheath cells of C
4
plants, which is due to the expenditure of the additional 12
molecules of ATP, is critical for their rapid photosynthetic rate, because CO
2
is limiting when light is abundant. A high
CO
2
concentration also minimizes the energy loss caused by photorespiration.
Tropical plants with a C
4
pathway do little photorespiration because the high concentration of CO
2
in their bundle-
sheath cells accelerates the carboxylase reaction relative to the oxygenase reaction. This effect is especially important at
higher temperatures. The geographic distribution of plants having this pathway (C
4
plants) and those lacking it (C
3
plants) can now be understood in molecular terms. C
4
plants have the advantage in a hot environment and under high
illumination, which accounts for their prevalence in the tropics. C
3
plants, which consume only 18 molecules of ATP per
hexose molecule formed in the absence of photorespiration (compared with 30 molecules of ATP for C
4
plants), are
more efficient at temperatures of less than about 28°C, and so they predominate in temperate environments.
Rubisco is found in bacteria, eukaryotes, and even archaea, though other photosynthetic components have not
been found in archaea. Thus, rubisco emerged early in evolution, when the atmosphere was rich in CO
2
and
almost devoid of O
2
. The enzyme was not originally selected to operate in an environment like the present one, which is
almost devoid of CO
2
and rich in O
2
. Photorespiration became significant about 60 million years ago, when the CO
2
concentration fell to present levels. The C
4
pathway is thought to have evolved in response to this selective pressure no
more than 30 million years ago and possibly as recently as 7 million years ago. It is interesting to note that none of the
enzymes are unique to C
4
plants, suggesting that this pathway was created using existing enzymes.
20.2.4. Crassulacean Acid Metabolism Permits Growth in Arid Ecosystems
Crassulacean acid metabolism (CAM) is yet another adaptation to increase the efficiency of the Calvin cycle.
Crassulacean acid metabolism, named after the genus Crassulacea (the succulents), is a response to drought as well as
warm conditions. In CAM plants, the stomata of the leaves are closed in the heat of the day to prevent water loss (Figure
20.18). As a consequence, CO
2
cannot be absorbed during the daylight hours when it is needed for glucose synthesis.
When the stomata open at the cooler temperatures of night, CO
2
is fixed by the C
4
pathway into malate, which is stored
in vacuoles. During the day, malate is decarboxylated and the CO
2
becomes available to the Calvin cycle. In contrast
with C
4
plants, CO
2
accumulation is separated from CO
2
utilization temporally in CAM plants rather than spatially.
II. Transducing and Storing Energy 20. The Calvin Cycle and the Pentose Phosphate Pathway 20.2. The Activity of the Calvin Cycle Depends on Environmental Conditions

Figure 20.14. Light Regulation of the Calvin Cycle. The light reactions of photosynthesis transfer electrons out of the
thylakoid lumen into the stroma and they transfer protons from the stroma into the thylakoid lumen. As a consequence of
these processes, the concentrations of NADPH, reduced ferredoxin (Fd), and Mg
2+
in the stroma are higher in the light
than in the dark, whereas the concentration of H
+
is lower in the dark. Each of these concentration changes helps couple
the Calvin cycle reactions to the light reactions.
II. Transducing and Storing Energy 20. The Calvin Cycle and the Pentose Phosphate Pathway 20.2. The Activity of the Calvin Cycle Depends on Environmental Conditions
Figure 20.15. Thioredoxin. The oxidized form of thioredoxin contains a disulfide bond. When thioredoxin is reduced
by reduced ferredoxin, the disulfide bond is converted into two free sulfhydryl groups. Reduced thioredoxin can cleave
disulfide bonds in enzymes, activating certain Calvin cycle enzymes and inactivating some degradative enzymes.
II. Transducing and Storing Energy 20. The Calvin Cycle and the Pentose Phosphate Pathway 20.2. The Activity of the Calvin Cycle Depends on Environmental Conditions
Table 20.1. Enzymes regulated by thioredoxin
Enzyme Pathway
Rubisco Carbon fixation in the Calvin cycle
Fructose 1,6-bisphosphatase Gluconeogenesis
Glyceraldehyde 3-phosphate dehydrogenase Calvin cycle, gluconeogenesis, glycolysis
Sedoheptulose bisphosphatase Calvin cycle

Glucose 6-phosphate dehydrogenase Pentose phosphate pathway
Phenylalanine ammonia lyase Lignin synthesis
Ribulose 5
-phosphate kinase Calvin cycle
NADP
+
-malate dehydrogenase
C
4
pathway
II. Transducing and Storing Energy 20. The Calvin Cycle and the Pentose Phosphate Pathway 20.2. The Activity of the Calvin Cycle Depends on Environmental Conditions
Figure 20.16. Enzyme Activation by Thioredoxin. Reduced thioredoxin activates certain Calvin cycle enzymes by
cleaving regulatory disulfide bonds.
II. Transducing and Storing Energy 20. The Calvin Cycle and the Pentose Phosphate Pathway 20.2. The Activity of the Calvin Cycle Depends on Environmental Conditions

Figure 20.17. C
4
Pathway. Carbon dioxide is concentrated in bundle-sheath cells by the expenditure of ATP in
mesophyll cells.
II. Transducing and Storing Energy 20. The Calvin Cycle and the Pentose Phosphate Pathway 20.2. The Activity of the Calvin Cycle Depends on Environmental Conditions
Figure 20.18. Electron Micrograph of an Open Stoma and a Closed Stoma. [Herb Charles Ohlmeyer/Fran Heyl
Associates.]
II. Transducing and Storing Energy 20. The Calvin Cycle and the Pentose Phosphate Pathway
20.3 the Pentose Phosphate Pathway Generates NADPH and Synthesizes Five-Carbon
Sugars
The pentose phosphate pathway meets the need of all organisms for a source of NADPH to use in reductive biosynthesis
(Table 20.2). This pathway consists of two phases: the oxidative generation of NADPH and the nonoxidative
interconversion of sugars (Figure 20.19). In the oxidative phase, NADPH is generated when glucose 6-phosphate is
oxidized to ribose 5-phosphate. This five-carbon sugar and its derivatives are components of RNA and DNA, as well as
ATP, NADH, FAD, and coenzyme A.
In the nonoxidative phase, the pathway catalyzes the interconversion of three-, four-, five-, six-, and seven-carbon sugars
in a series of nonoxidative reactions that can result in the synthesis of five-carbon sugars for nucleotide biosynthesis or
the conversion of excess five-carbon sugars into intermediates of the glycolytic pathway. All these reactions take place
in the cytosol. These interconversions rely on the same reactions that lead to the regeneration of ribulose 1,5-
bisphosphate in the Calvin cycle.
20.3.1. Two Molecules of NADPH Are Generated in the Conversion of Glucose 6-
phosphate into Ribulose 5-phosphate
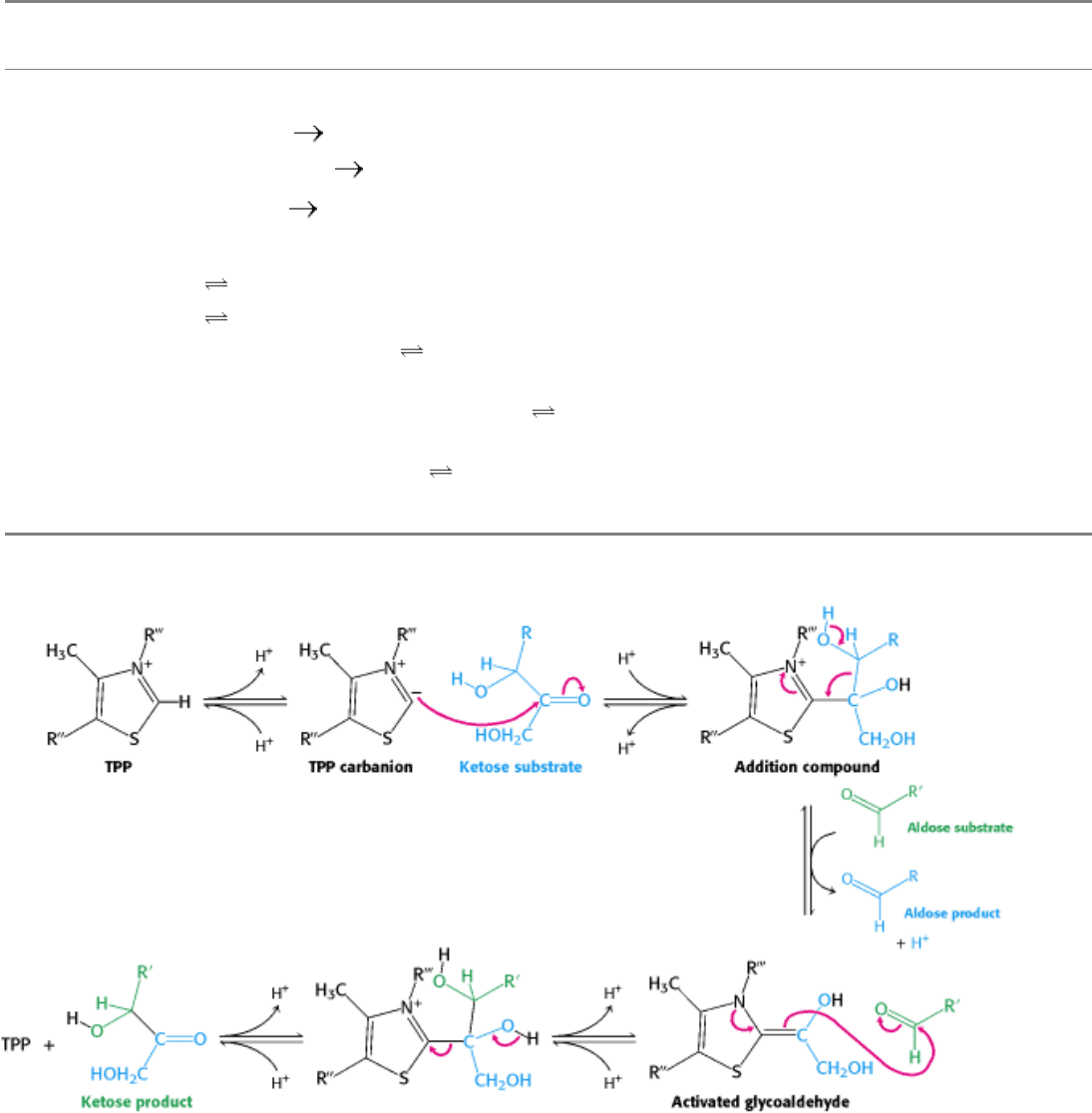
The oxidative phase of the pentose phosphate pathway starts with the dehydrogenation of glucose 6-phosphate at carbon
1, a reaction catalyzed by glucose 6-phosphate dehydrogenase (Figure 20.20). This enzyme is highly specific for NADP
+
; the K
M
for NAD
+
is about a thousand times as great as that for NADP
+
. The product is 6-phosphoglucono-δ-lactone,
which is an intramolecular ester between the C-1 carboxyl group and the C-5 hydroxyl group. The next step is the
hydrolysis of 6-phosphoglucono-δ-lactone by a specific lactonase to give 6-phosphogluconate. This six-carbon sugar is
then oxidatively decarboxylated by 6-phosphogluconate dehydrogenase to yield ribulose 5-phosphate. NADP
+
is again
the electron acceptor. The final step in the synthesis of ribose 5-phosphate is the isomerization of ribulose 5-phosphate
by phosphopentose isomerase (see Figure 20.11)
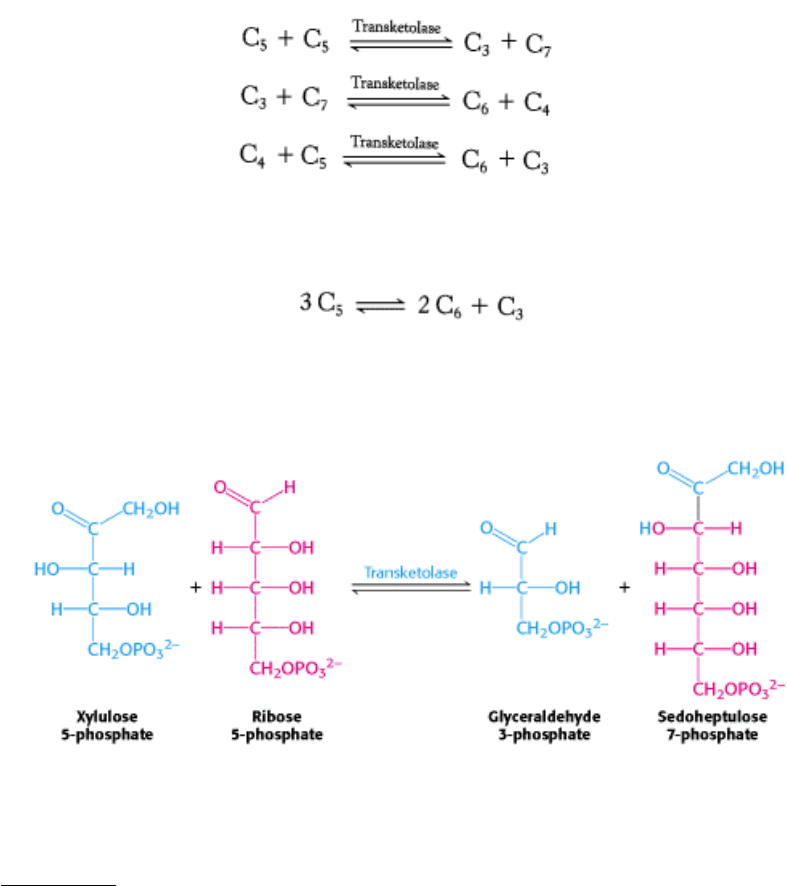
20.3.2. The Pentose Phosphate Pathway and Glycolysis Are Linked by Transketolase
and Transaldolase
The preceding reactions yield two molecules of NADPH and one molecule of ribose 5-phosphate for each molecule of
glucose 6-phosphate oxidized. However, many cells need NADPH for reductive biosyntheses much more than they need
ribose 5-phosphate for incorporation into nucleotides and nucleic acids. In these cases, ribose 5-phosphate is converted
into glyceraldehyde 3-phosphate and fructose 6-phosphate by transketolase and transaldolase. These enzymes create a
reversible link between the pentose phosphate pathway and glycolysis by catalyzing these three successive reactions.
The net result of these reactions is the formation of two hexoses and one triose from three pentoses:
The first of the three reactions linking the pentose phosphate pathway and glycolysis is the formation of glyceraldehyde
3-phosphate and sedohep-tulose 7-phosphate from two pentoses.
The donor of the two-carbon unit in this reaction is xylulose 5-phosphate, an epimer of ribulose 5-phosphate. A ketose is
a substrate of transketolase only if its hydroxyl group at C-3 has the configuration of xylulose rather than ribulose.
Ribulose 5-phosphate is converted into the appropriate epimer for the transketolase reaction by phosphopentose
epimerase (see Figure 20.11) in the reverse reaction of that which occurs in the Calvin cycle.
Glyceraldehyde 3-phosphate and sedoheptulose 7-phosphate generated by the transketolase then react to form fructose 6-
phosphate and erythrose 4-phosphate.

This synthesis of a four-carbon sugar and a six-carbon sugar is catalyzed by transaldolase.
In the third reaction, transketolase catalyzes the synthesis of fructose 6-phosphate and glyceraldehyde 3-phosphate from
erythrose 4-phosphate and xylulose 5-phosphate.
The sum of these reactions is
Xylulose 5-phosphate can be formed from ribose 5-phosphate by the sequential action of phosphopentose isomerase and
phosphopentose epimerase, and so the net reaction starting from ribose 5-phosphate is
Thus, excess ribose 5-phosphate formed by the pentose phosphate pathway can be completely converted into glycolytic
intermediates. Moreover, any ribose ingested in the diet can be processed into glycolytic intermediates by this pathway.
It is evident that the carbon skeletons of sugars can be extensively rearranged to meet physiologic needs (Table 20.3).
20.3.3. Transketolase and Transaldolase Stabilize Carbanionic Intermediates by
Different Mechanisms
The reactions catalyzed by transketolase and transaldolase are distinct yet similar in many ways. One difference is that
transketolase transfers a two-carbon unit, whereas transaldolase transfers a three-carbon unit. Each of these units is
transiently attached to the enzyme in the course of the reaction. In transketolase, the site of addition of the unit is the

thiazole ring of the required coenzyme thiamine pyrophosphate. Transketolase is homologous to the E
1
subunit of the
pyruvate dehydrogenase complex (Section 17.1.1) and the reaction mechanism is similar (Figure 20.21).
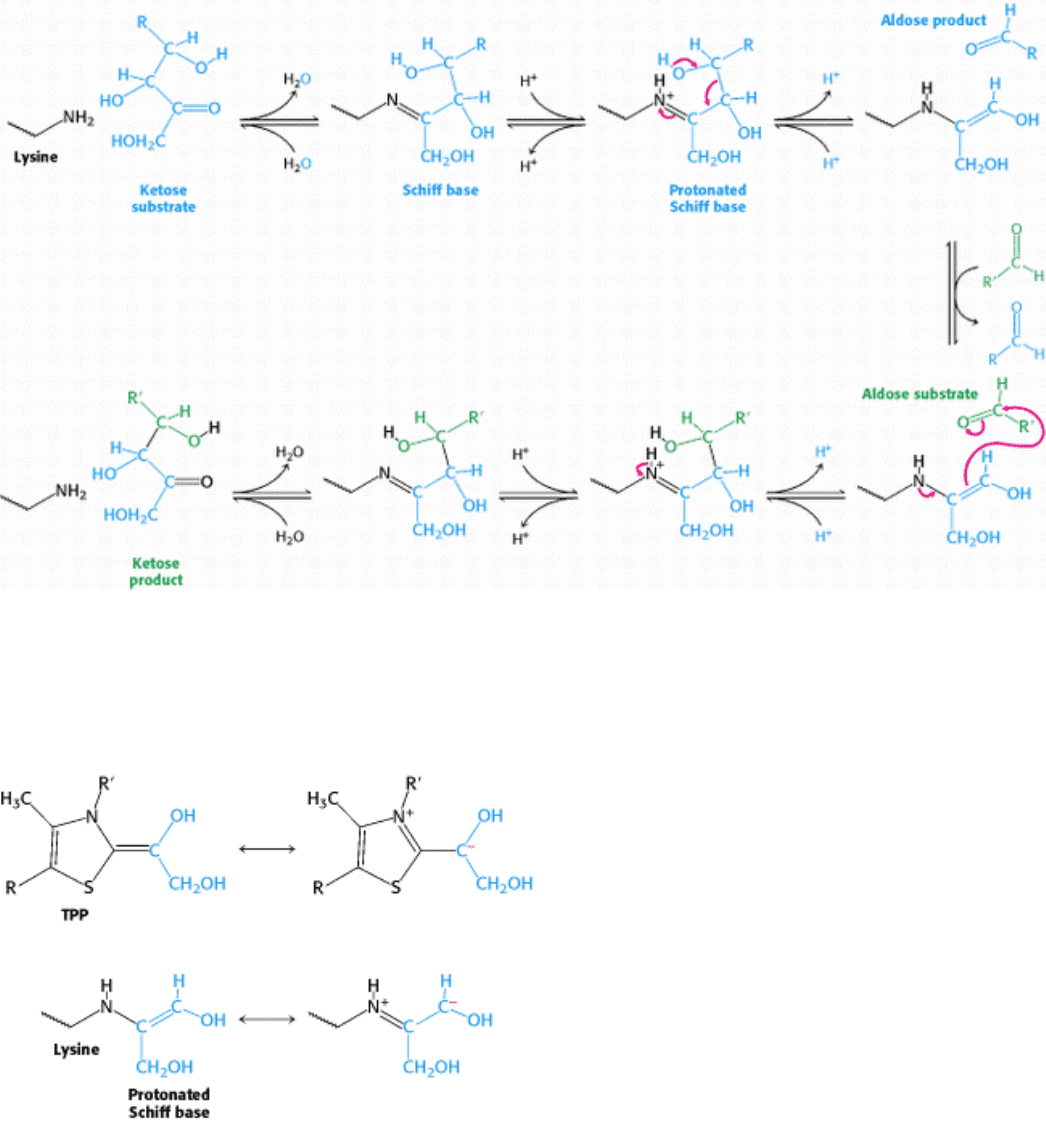
The C-2 carbon atom of bound TPP readily ionizes to give a carbanion. The negatively charged carbon atom of this
reactive intermediate attacks the carbonyl group of the ketose substrate. The resulting addition compound releases the
aldose product to yield an activated glycoaldehyde unit. The positively charged nitrogen atom in the thiazole ring acts as
an electron sink in the development of this activated intermediate. The carbonyl group of a suitable aldose acceptor then
condenses with the activated glycoaldehyde unit to form a new ketose, which is released from the enzyme.
Transaldolase transfers a three-carbon dihydroxyacetone unit from a ketose donor to an aldose acceptor. Transaldolase,
in contrast with transketolase, does not contain a prosthetic group. Rather, a Schiff base is formed between the carbonyl
group of the ketose substrate and the ε-amino group of a lysine residue at the active site of the enzyme (Figure 20.22).
This kind of covalent enzyme-substrate intermediate is like that formed in fructose 1,6-bisphosphate aldolase in the
glycolytic pathway (Section 16.1.3) and, indeed, the enzymes are homologous. The Schiff base becomes protonated, the
bond between C-3 and C-4 is split, and an aldose is released. The negative charge on the Schiff-base carbanion moiety is
stabilized by resonance. The positively charged nitrogen atom of the protonated Schiff base acts as an electron sink. The
Schiff-base adduct is stable until a suitable aldose becomes bound. The dihydroxyacetone moiety then reacts with the
carbonyl group of the aldose. The ketose product is released by hydrolysis of the Schiff base. The nitrogen atom of the
protonated Schiff base plays the same role in transaldolase as the thiazole-ring nitrogen atom does in transketolase. In
each enzyme, a group within an intermediate reacts like a carbanion in attacking a carbonyl group to form a new carbon-
carbon bond. In each case, the charge on the carbanion is stabilized by resonance (Figure 20.23).
II. Transducing and Storing Energy 20. The Calvin Cycle and the Pentose Phosphate Pathway 20.3 the Pentose Phosphate Pathway Generates NADPH and Synthesizes Five-Carbon Sugars
Table 20.2. Pathways requiring NADPH
Synthesis
Fatty acid biosynthesis
Cholesterol biosynthesis
Neurotransmitter biosynthesis
Nucleotide biosynthesis
Detoxification
Reduction of oxidized glutathione
Cytochrome P450 monooxygenases
II. Transducing and Storing Energy 20. The Calvin Cycle and the Pentose Phosphate Pathway 20.3 the Pentose Phosphate Pathway Generates NADPH and Synthesizes Five-Carbon Sugars
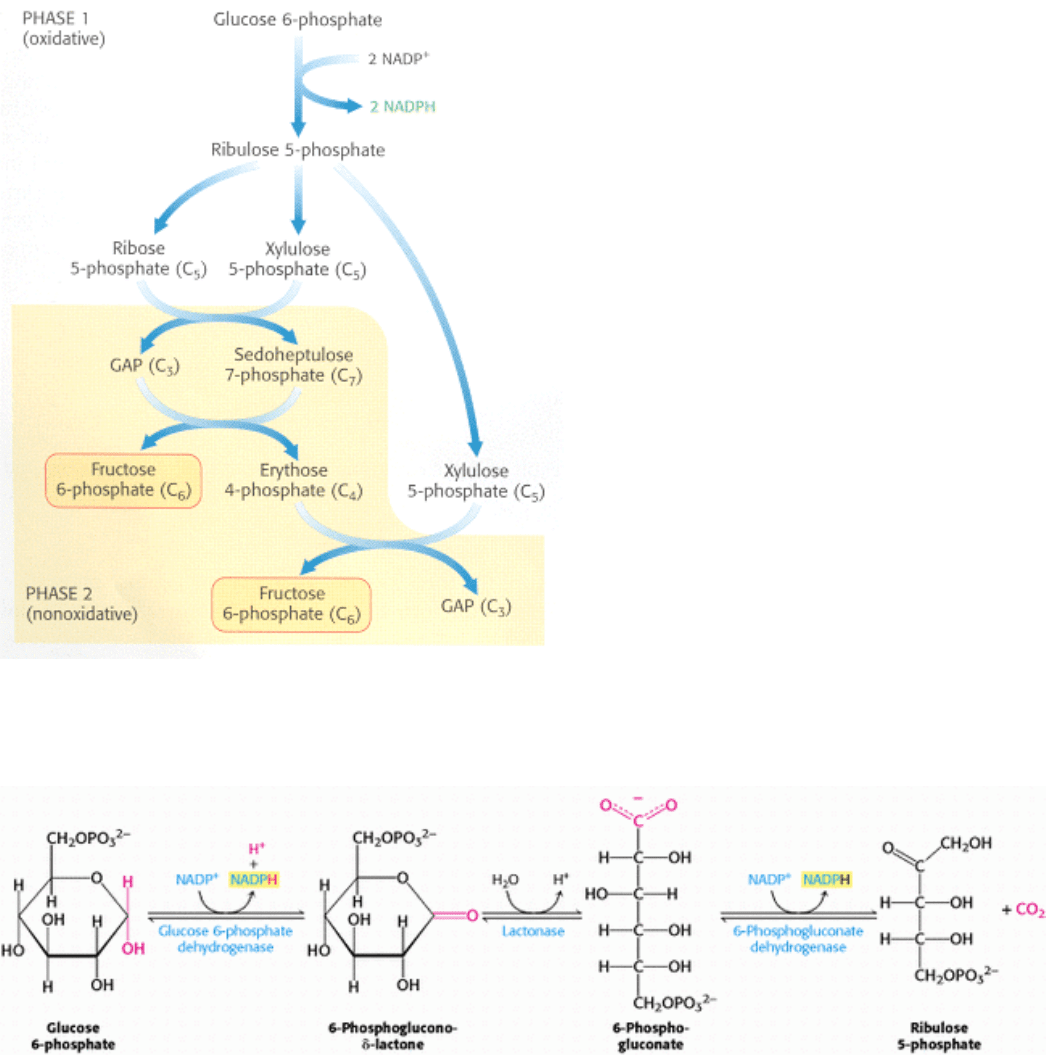
Figure 20.19. Pentose Phosphate Pathway. The pathway consists of (1) an oxidative phase that generates NADPH and
(2) a nonoxidative phase that interconverts phosphorylated sugars.
II. Transducing and Storing Energy 20. The Calvin Cycle and the Pentose Phosphate Pathway 20.3 the Pentose Phosphate Pathway Generates NADPH and Synthesizes Five-Carbon Sugars
Figure 20.20. Oxidative Phase of the Pentose Phosphate Pathway. Glucose 6-phosphate is oxidized to 6-
phosphoglucono-δ-lactone to generate one molecule of NADPH. The lactone product is hydrolyzed to 6-
phosphogluconate, which is oxidatively decarboxylated to ribulose 5-phosphate with the generation of a second molecule
of NADPH.
II. Transducing and Storing Energy 20. The Calvin Cycle and the Pentose Phosphate Pathway 20.3 the Pentose Phosphate Pathway Generates NADPH and Synthesizes Five-Carbon Sugars
Table 20.3. Pentose phosphate pathway
Reaction Enzyme
Oxidative phase
Glucose 6-phosphate + NADP
+
6-phosphoglucono-δ-lactone + NADPH + H
+
Glucose 6-phosphate dehydrogenase
6-Phosphoglucono-δ-lactone + H
2
O 6-phosphogluconate + H
+
Lactonase
6-Phosphogluconate + NADP
+
ribulose 5-phosphate + CO
2
+ NADPH
6-Phosphogluconate dehydrogenase
Nonoxidative Phase
Ribulose 5-phosphate
ribose 5-phosphate Phosphopentose isomerase
Ribulose 5-phosphate
xylulose 5-phosphate Phosphopentose epimerase
Xylulose 5-phosphate + ribose 5-phosphate
sedoheptulose 7-phosphate +
glyceraldehyde 3-phosphate
Transketolase
Sedoheptulose 7-phosphate + glyceraldehyde 3-phosphate
fructose 6-phosphate
+ erythrose 4-phosphate
Transaldolase
Xylulose 5-phosphate + erythrose 4-phosphate
fructose 6-phosphate +
glyceraldehyde 3-phosphate
Transketolase
II. Transducing and Storing Energy 20. The Calvin Cycle and the Pentose Phosphate Pathway 20.3 the Pentose Phosphate Pathway Generates NADPH and Synthesizes Five-Carbon Sugars
Figure 20.21. Transketolase Mechanism. The carbanion of thiamine pyrophosphate (TPP) attacks the ketose substrate.
Cleavage of a carbon-carbon bond frees the aldose product and leaves a two-carbon fragment joined to TPP. This
activated glycoaldehyde intermediate attacks the aldose substrate to form a new carbon-carbon bond. The ketose product
is released, freeing the TPP for the next reaction cycle.
II. Transducing and Storing Energy 20. The Calvin Cycle and the Pentose Phosphate Pathway 20.3 the Pentose Phosphate Pathway Generates NADPH and Synthesizes Five-Carbon Sugars
Figure 20.22. Transaldolase Mechanism. The reaction begins with the formation of a Schiff base between a lysine
residue in transaldolase and the ketose substrate. Protonation of the Schiff base leads to release of the aldose product,
leaving a three-carbon fragment attached to the lysine residue. This intermediate adds to the aldose substrate to form a
new carbon-carbon bond. The reaction cycle is completed by release of the ketose product from the lysine side chain.
II. Transducing and Storing Energy 20. The Calvin Cycle and the Pentose Phosphate Pathway 20.3 the Pentose Phosphate Pathway Generates NADPH and Synthesizes Five-Carbon Sugars
Figure 20.23. Carbanion Intermediates. For transketolase and transaldolase, a carbanion intermediate is stabilized by
resonance. In transketolase, TPP stabilizes this intermediate; in transaldolase, a protonated Schiff base plays this role.