Pugnaire F.I. Valladares F. Functional Plant Ecology
Подождите немного. Документ загружается.

(ABA), which mediates stomatal closure (Davies and Zhang 1991). When soil water is not
limited, and when the internal CO
2
concentrations fall below ambient levels in the atmosphere,
stomata opens in sunlight. In approximately half of the known species, stomata have the
additional capability of sensing the relative humidity (RH) of the ambient air and tend to
progressively close as RH in the air adjacent to the leaves decreases. Rapid stomatal opening is
mediated by movement of K
þ
from the epidermal cells to the guard cells (Figure 6.2c–f ). The
movement of K
þ
makes p
c
less negative in the epidermal cells and more negative in the guard
cells; consequently, water flows from the epidermal cells to the guard cell, P
t
falls in the epidermal
Guard cell
Subsidiary cell
Epidermal cell
Closed stomate
Open stomate
Vein
Cuticle
Palisade
Mesophyll
Lower
epidermis
Guard cell
Upper
epidermis
(d)(c)
(f)
(a)
(b)
(e)
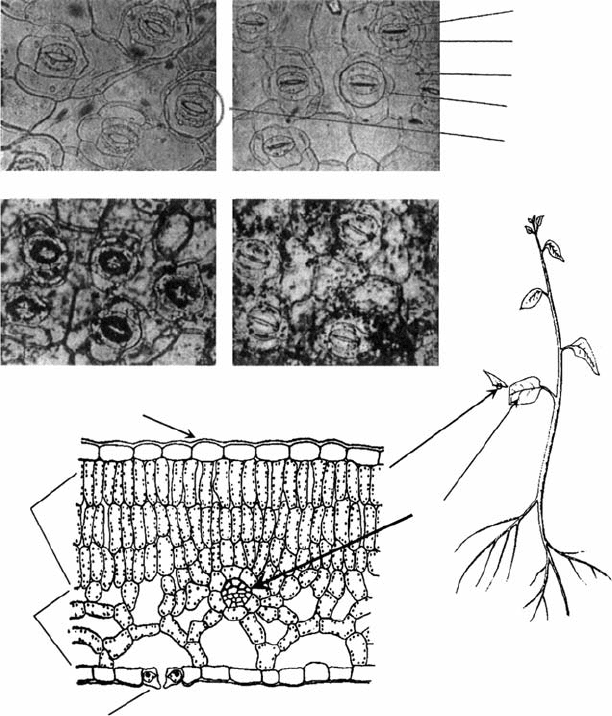
FIGURE 6.2 (a) A typical plant. (b) The lower leaf has been cut and the cross section is shown enlarged
approximately 750. (c–f ) Photographs of the lower leaf surface enlarged approximately 1000. (e) and
(f ) have been stained to show location of K
þ
. (c) and (e) illustrate the open state of stomata when the
leaf is exposed to light. Stoma is from the Greek and means mouth (plural, stomata); the two guard cells
for a structure looks much like a mouth. (d) and (f ) are similar leaves in light but also exposed to abscisic
acid (ABA), causing stomata to remain closed in light. Note that in open stomata, K
þ
is concentrated in
the guard cells (e) and the guard cells have moved apart to form an air passage for gas exchange (the
stomatal pore). Note that the K
þ
is located in the epidermal cells when the stomata are closed
(f ). (Adapted from Kramer, P.J., Water Relations of Plants, Academic Press, New York, 1983; Bidwell,
R.G.S., Plant Physiology, Macmillan, New York, 1979.)
Francisco Pugnaire/Functional Plant Ecology 7488_C006 Final Proof page 180 10.5.2007 2:47pm Compositor Name: VBalamugundan
180 Functional Plant Ecology
cells, and P
t
rises in the guard cells. The mechanical effect of this water movement and change in
P
t
causes the guard cells to swell into the epidermal region and open a stomatal pore.
The evaporative flux density (E) of water vapor from leaves is ultimately governed by
Fick’s law of diffusion of gases in air. The control exercised by the plant is to change the area
available for vapor diffusion through the opening and closing of stomatal pores. The value of
E (mmol water s
1
m
2
of leaf surface) is given by
E ¼ g
L
(X
i
X
o
), (6:8)
where g
L
is the diffusional conductance of the leaf (largely controlled by the stomatal
conductance, g
s
), X
i
is the mole fraction of water vapor at the evaporative surface of the
palisade and mesophyll cells, and X
o
is the mole fraction of water vapor in the ambient air
surrounding the leaf. The mole fraction is defined as X ¼n
w
=N, where n
w
is the number of
moles of water vapor and N is the number of moles of all gas molecules, including water
vapor, the most abundant gas molecules are N
2
and O
2
. The dependence of g
L
on some
environmental and physiological variables is illustrated in Figure 6.3.
The maximum value of X occurs when RH is 100%, that is, when the air is saturated with
water vapor; the maximum value of X increases exponentially with the Kelvin temperature of
the air. The air at the evaporating surface of leaves is at the temperature of the leaf (T
L
), and
X
i
is taken as the maximum value of X at saturation, which can be symbolized as X
i
¼X [T
L
].
(a)
0
01234−2.5 −2.0 −1.5 −1.0 −0.5 0.0
⌿
L
(MPa)V
PD
(kPa)
500
1.0
0.8
0.6
0.4
0.2
0.0
1.0
0.8
0.6
0.4
0.2
0.0
1000
PAR (µE m
−2
s
−1
)
T
L
(°C)
Relative g
L
Relative g
L
1500 2000 0 15 20 25 30 35
(b)
(d)(c)
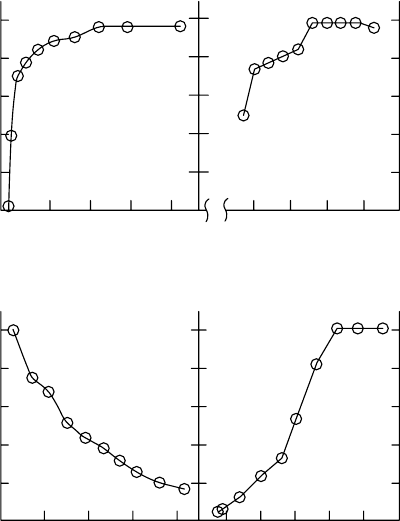
FIGURE 6.3 Relationship between change of g
L
relative to the maximum value, g
L
=g
L,max
, and various
environmental factors measured on Acer saccharum leaves. The environmental factors were: (a) photo-
synthetically active radiation (PAR), (b) leaf temperature (T
L
), (c) vapor pressure deficit (V
PD
), and
(d) leaf water potential (C
L
). (Adapted from Yang, S., Liu, X., and Tyree, M.T., J. Theor. Biol., 191,
197, 1998. With permission.)
Francisco Pugnaire/Functional Plant Ecology 7488_C006 Final Proof page 181 10.5.2007 2:47pm Compositor Name: VBalamugundan
Water Relations and Hydraulic Architecture 181
The value of X
o
depends on the microclimate near the leaf, that is, the air temperature and
RH. However, the microclimate of the leaf is strongly influenced by the behavior of the plant
community surrounding the leaf. Therefore, although the leaf has direct control over the
value of g
s
and g
L
, it has less control over E than might appear from Equation 6.8.
The qualitative aspects of how leaves influence their own microclimate is easily explained.
When the sun rises in the morning, the radiant energy load on the leaf increases. This has two
effects: T
L
rises as the sun warms the leaves, and hence, X
i
rises and g
L
increases as stomata
open. However, the increased evaporation from the leaves causes X
o
to rise as all the water
vapor is added to the ambient air. Even changes of g
L
under constant radiant energy load
causes less change in E than might be expected from Equation 6.8. When g
L
doubles, E also
doubles, but only temporarily. The increased E lowers T
L
because of increased evaporative
cooling, and hence lowers X
i
. The increased E from all of the leaves in a stand eventually
increases X
o
, hence X
i
X
o
declines, causing a decline in E. Consequently, Equation 6.8 is not
very useful in predicting the value of E at the level of plant communities. Leaf-level behavior
can be extrapolated to community-level equations if we take into account leaf-level solar
energy budgets, that is, equations that describe light absorption by leaves at all wavelengths
and the conversion of this energy to temperature and heat fluxes. Studies of solar energy
budgets have been conducted at both the leaf and stand-level (Slatyer 1967, and Chang 1968;
see also Section ‘‘Factors Controlling the Rate of Water Uptake and Movement’’).
Most of the changes in E at the leaf level can be explained in terms of net radiation
absorbed at the stand level, which is relatively easy to measure. This relationship is illustrated
in Figure 6.4, where daily values of E at the leaf level (kg m
2
day
1
) are correlated with daily
0
0.0
0.5
1.0
1.5
2.0
Bp (9)
Fm (1)
Ip (2)
Cs (4)
Cd (3)
Pa (4)
Bf (2)
Da (2)
246
E
∗
(kg m
−2
day
−1
)
Net radiation (MJ m
−2
day
−1
)
8101214
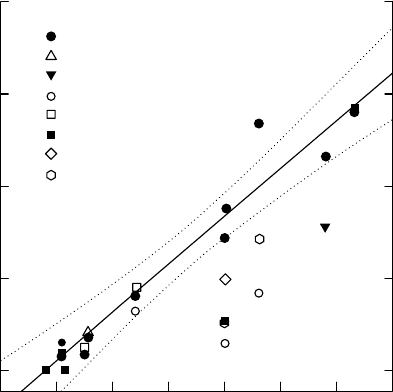
FIGURE 6.4 Correlation of daily water use of leaves (E*) and net radiation (NR). Data are from
potometer experiments with nine woody cloud forest species in Panama. The number of days per species
is given in parenthesis. The regression (with 95% confidence intervals) is for Bp: Baccharis pendunculata
(E* ¼0.20 þ1.29 NR, r
2
¼0.92). Even when the data for all species are pooled, NR proved to be a very
good predictor of daily water use (E* ¼0.2 þ1.04 NR, r
2
¼0.72). Cm, Citharexylum macradenium; Cd,
Croton draco; Fm, Ficus macbridei; Ip, Inga punctata; Pa, Parathesis amplifolia; Bf, Blakea foliacea; Cs,
Clusia stenophylla; and Da, Dendropanax arboreus. (From Zotz, G., Tyree, M.T. Patin
˜
o, S., and
Carlton, M.R., Trees, 12, 302, 1998. With permission.)
Francisco Pugnaire/Functional Plant Ecology 7488_C006 Final Proof page 182 10.5.2007 2:47pm Compositor Name: VBalamugundan
182 Functional Plant Ecology
values of net radiation (MJ m
2
of ground per day) measured with an Eppley-type net
radiometer. Equations presented elsewhere to describe stand-level E in terms of net radiation
and other factors are validated by relationships such as shown in Figure 6.4. The other factors
most commonly included account for plant control over E through leaf area index (leaf area
per unit ground area), the effect of drought on g
L
, and ambient temperature (see Section
‘‘Factors Controlling the Rate of Water Uptake and Movement’’).
TISSUE–WATER RELATIONS (PRESSURE–VOLUME CURVES)
From the preceding section, the reader might falsely conclude that plants can lose water
from leaves without any negative impact on the growth and survival. However, there is
more to water loss than is apparent from stomatal physiology and the energy interactions
between leaves and their immediate environment. Whenever leaves lose water faster than the
rate of water uptake by roots, the water potential in the xylem (C
x
) and of the leaf cells (C
c
)
must also fall (see Section ‘‘Water Relations of Plant Cells’’). Most of the decline in C
x
is
due to a drop in P
x
, and when P
x
becomes too negative, cavitations occur that prevent water
flow through vessels (see Section ‘‘Cohesion–Tension Theory and Xylem Dysfunction’’).
Most of the decline in C
c
is due to a drop in P
t
, and when P
t
becomes too small, cell growth
stops. Growing cells are surrounded by relatively ridged ‘‘wooden boxes’’ consisting of
cellulose cell walls. Cell walls must be stretched plastically to grow large, and the motive
force on plastic stretch is the force of P
t
against the cell walls. As leaves lose water, P
t
and
P
x
must fall for the plant to extract water from the soil at a rate approximately equal to the
rate of water loss. As soils become dry, values of P
t
and P
x
must fall even more for plants
to extract water from soils. From Equation 6.7 it can be seen that P
t
¼C
c
p
c
, hence
plants have some control over P
t
through ‘‘adjustments’’ in p
c
. Some species have evolved
to have lower values of p
c
than others, and some species can make p
c
more negative in
response to drought by increasing the osmolal concentration (C) of solutes in their cells
(because p
c
¼RTC).
The relationship between C and its components (P and p) in leaves can be described by a
leaf-level Ho
¨
fler diagram (Figure 6.2a). Many studies have reported the comparative physi-
ology of tissue–water relationship of leaves and have discussed how differences in these
relationships might explain ecological adaptation of plants. Readers interested in learning
more should consult the references contained in Tyree and Jarvis (1982). The following is a
brief overview of how Ho
¨
fler diagrams are measured in leaves and some ecological applica-
tions of this information.
The fastest way to derive a Ho
¨
fler diagram is to measure the water potential of leaves
(C
leaf
) versus water loss using a Scholander–Hammel pressure bomb (Scholander et al. 1965,
Tyree and Hammel 1972). This is done by obtaining a series of balance pressure (P
B
) versus
weight loss of leaves or shoots; the P
B
is an approximate measure of the C
leaf
. The pressure
bomb is a metal chamber into which is placed an excised leaf or shot (Figure 6.5a) that is at an
unknown water potential, C
leaf
. When gas pressure (P
gas
) is applied to the leaf surface, the
pressure of the fluid in the cells is increased by an equal amount so that
C
leaf
¼ C
c
¼ p
c
þP
t
þP
gas
. The xylem water potential and cell water potential are at equili-
brium, C
c
¼C
x
; from this it follows that p
x
þP
x
¼p
c
þP
t
þP
gas
or P
x
¼p
c
þP
t
þP
gas
p
x
.
When the P
gas
is at the balance pressure (P
B
), P
x
has risen to zero and xylem sap is squeezed
out of the end of the branch or petiole protruding outside the pressure bomb (Figure 6.5).
Therefore, we have P
B
¼(p
c
þP
t
) þp
x
¼C
leaf
, when the branch or leaf was outside the
bomb) þp
x
¼P
x
. P
B
is usually approximated by C
leaf
since p
x
is usually much smaller than
C
leaf
. However, P
B
really measures P
x
and the pressure bomb cannot measure pressure
gradients within a leaf because all gradients dissipate during the P
B
measurement. In some
cases these distinctions are important to remember.
Francisco Pugnaire/Functional Plant Ecology 7488_C006 Final Proof page 183 10.5.2007 2:47pm Compositor Name: VBalamugundan
Water Relations and Hydraulic Architecture 183
0
0.5 0.6 0.7 0.8 0.9 1.0
0.0
−4
−3
−2
−1
0
1
2
0.3
0.6
A
Pressure
gauge
Pressure
release valve
Connected to
compressed
air source
Pressure
bomb
(a)
B
C
(b)
1/P
B
(MPa
−1
)
Water potential or components (MPa)
P
B
(MPa)
0.9
1.2
1.5
24
⌿
6 8 10 12
∆W (g)
R
wc
P
t
14 16 18 20 22 24
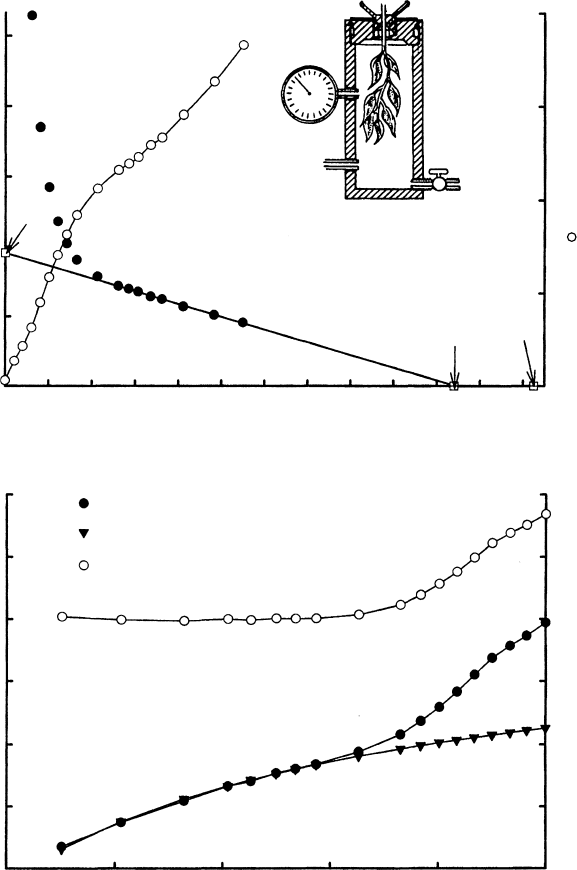
FIGURE 6.5 Scholander–Hammel pressure bomb and analysis of pressure bomb data. (a) A pressure
bomb is shown in the inset. Open circles (right axis): Original data of balance pressure (P
B
) versus weight
loss of shoot in the bomb. Weight loss is induced by removing the shoot from the bomb and allowing
water to evaporate. Closed circles (left axis): Transformed data of 1=P
B
versus weight loss. The liner
portion of the curve has been extrapolated to two points, A ¼y-intercept ¼1=p
o
, where p
o
is the
average osmotic pressure of the symplast at full hydration when C ¼0; B ¼x-intercept ¼weight of water
in the symplast when C ¼0 [this value can also be computed from (1=p
0
)=slope]; C ¼total
water content of the shoot ¼maximum weight loss when oven-dried. (b) The Ho
¨
fler diagram for the
whole shoot derived from the data in (a). The x-axis is relative water content of the shoot and the y-axis
is p ¼osmotic potential of the symplasm (solid triangles), P
t
¼turgor pressure of symplasm (open
circles), and C ¼total water potential of the symplasm (solid circles).
Francisco Pugnaire/Functional Plant Ecology 7488_C006 Final Proof page 184 10.5.2007 2:47pm Compositor Name: VBalamugundan
184 Functional Plant Ecology

A pressure–volume curve is obtained by slowly dehydrating a shoot and obtaining a series
of weights, W, versus P
B
values. If W
o
was the original weight, then the cumulative weight
loss is DW ¼W
o
W. The pressure–volume curve is a plot of 1=P
B
versus DW. Strictly
speaking, the pressure–volume curve should be called a pressure–weight curve, but if DW is
given in g, then that is the same as volume in mL since 1 mL of water weighs 1 g. The
pressure–volume curve has a curved region for small DW values and a linear region for larger
DW values (Figure 6.5b). When the linear portion of the plot is extrapolated back DW ¼0, the
y-intercept (the point marked A in Figure 6.5a) is equal to 1=p
o
, where p
o
is the solute
potential of the living cells at zero water potential. The point marked B in Figure 6.5a is the
turgor loss point (C
tlp
), that is, the value of C
leaf
when P
t
reaches zero. The x-intercept
(point C) is the volume of water contained in the symplast (W
s
¼total water in the proto-
plasm and vacuoles of all living cells), and the difference in x-values (D–C) is the amount of
water in the apoplast (W
a
¼total water in xylem and cell walls). Ho
¨
fler diagrams for shoots or
leaves are usually plots of C, p, and P
t
versus relative water content of the shoot or leaf.
Relative water content (R
WC
) is defined as (the current water content)=(the maximum water
content at full hydration), R
WC
¼(W
o
DW )=(W
o
W
d
), where W
d
is the dry weight. Values
of p at different R
WC
are calculated from p
o
W
s
=(W
o
DW ), values of C are equated to
P
B
, and values of P
t
are calculated from P
B
p
o
=R
WC
. The justification for these relation-
ships is given in a report by Tyree and Hammel (1972).
As plants dry, changes in p can be caused by changes in symplastic water content, W
s
,orin
the number of moles of solute in the symplasm, N
s
, because p ¼RTN
s
=W
s
. Considerbale
emphasis has been placed on demonstrating changes in p as a result to changes in N
s
(Turner
and Jones 1980). A change in p caused by a change in N
s
is called an osmotic adjustment.
Diurnal changes in p ranging from 0.4 to 1.6 MPa have been reported for some plants; the
amount of change resulting from diurnal changes in N
s
is in the range of 0.2–0.8 MPa. Medium-
term changes in p induced by slow soil dehydration have also been attributed to osmotic
adjustment in drought-stressed versus unstressed plants. Osmotic adjustments of 0.1–1 MPa
have been reported over periods of 3 days to 3 weeks. Long-term or seasonal changes in p range
from 0.2 to 1.8 MPa; some of the largest changes are recorded during the onset of winter in
temperate plants and appear to be correlated to changes in frost tolerance. The degree of
diurnal, medium-term, and long-term osmotic adjustment varies widely between species.
There are some species that have shown little or no adjustment (Tyree and Jarvis 1982).
Low values of p in plants should enhance the ability of plants to take up soil water under
dry or saline conditions (Tyree 1976). This advantage is probably marginal in sandy soils
because the available water reserves at soil water potentials less than 0.4 MPa are very
small; therefore, the plant’s ability to grow deep roots (Section ‘‘Cohesion–Tension Theory
and Xylem Dysfunction’’) is probably of greater advantage. In clay soils, however, there are
considerable water reserves at water potentials less than 0.4 MPa, so that low leaf and root
values of p may be as important as root growth in assisting water uptake. Low values of p in
leaves also enable P
t
to remain above zero at lower values of C than otherwise would be
possible as C falls. This allows the maintenance of open stomata with larger apertures and
high stomatal conductances and higher net rates of photosynthesis down to lower values of C
than would be the case if p were higher (less negative). Osmotic adjustments and=or lower p
values also enable maintenance of turgor pressure for growth, since it has been shown that the
rate of volume growth (r ¼dV=dt) of a cell is given by m(P
t
Y ), where m is the growth rate
constant of a cell and Y is the yield point of the cell (Green et al. 1971).
Some attention in the past has been focused on the slope of the P
t
line in the Ho
¨
fler
diagram in which the x-axis is R
WC
. The bulk modulus of elasticity of a tissue is defined as
« ¼
DP
t
DR
WC
R
WC
: (6:9)
Francisco Pugnaire/Functional Plant Ecology 7488_C006 Final Proof page 185 10.5.2007 2:47pm Compositor Name: VBalamugundan
Water Relations and Hydraulic Architecture 185
The higher value of « represents higher value of the slope of P
t
versus R
wc
. A large value of « is
seen as an advantage for water conservation. For a plant to extract water from the soil, it must
first lose some water so that its C
leaf
falls below the soil water potential. Because most of the
change in C
leaf
is due to the change in P
t
, a large value of « means that a plant would have to
lose less water to lower C
leaf
than would a plant with a small value of «.
Although values of « have been reported to range from 0.5 to 20 MPa, the adaptive
advantages of large versus small « have never been clearly established (Tyree and Jarvis 1982)
because the ecological advantage of a large « may be rather marginal. All plants lose water
during the day and regain most or all of the lost water during the night. Therefore, the
advantage of less water loss means more constant concentrations of biochemical substrates
during the day. Leaves can lose anywhere from 1% to 20% of their water content during the
day, and this water loss would cause a corresponding 1%–20% diurnal variation in substrates.
Because the concentration of reactants and products would normally change during the day,
even without the influence of leaf dehydration, it is not clear how much is gained by keeping
relatively constant cell volume because of large «.
WATER ABSORPTION BY PLANT ROOTS
The primary factor affecting the pattern of water extraction by plants from soils is the rooting
depth. Rooting depths can be extremely variable depending on soil conditions and species of
plant producing the roots (Figure 6.6a). Many of the early studies of rooting depth and
branching pattern of roots were performed in the 1920s and 1930s in deep, well-aerated
prairie soils, where roots penetrate to great depths. At the extreme, roots have been traced to
depths of 10–25 m, for example, alfalfa (10 m), longleaf pine, (17 m) (Kramer 1983), and
drought-evading species in the California chaparral (25 m) (S. Davis, April 1991, personal
communication). The situation is very different for plants growing in heavy soils, where 90%
of the roots can be found in the upper 0.5–1.0 m.
In seasonally dry regions, for example, central Panama, the majority of the roots may be
located in the upper 0.5 m, but it is far from clear if the majority of water absorption occurs in
the upper 0.5 m during the dry season. Water use by many evergreen trees is higher in the dry
season than in the wet season, although the upper 1 m of the soils is much drier than the leaves
of the trees (C
soil
< C
leaf
) and excavations have shown that liana roots are less than 5 m in
depth and hence extract water from quite wet soil during the dry season (M.T. Tyree,
February 1989, personal observation). Hence, the role of shallow versus deep roots of
woody species deserves more study.
In addition, deeply rooted species may contribute to the water supply of shallow-rooted
species through a process called hydraulic lift (Richards and Caldwell 1987). In one study, it
was found that shallow-rooted species growing within 1–5 m of the base of maple trees were in
a better water balance than the same species growing more than 5 m away (Dawson 1993).
Each night, the C
soil
at a depth of 0.5 m increased underneath maple trees, but C
soil
did not
increase at distances greater than 5 m from the trees. This indicated that the deep maple roots
were in contact with moist soil and were capable of transporting water from deep maple
roots to shallow maple roots overnight. Since the water potential of the shallow maple roots
exceeded the water potential of the adjacent soil, water flow from the shallow maple roots to
the adjacent soil contributed to the overnight rehydration of shallow soils.
Roots absorb both water and mineral solutes found in the soil, and the flow of solutes and
water interact with each other. The mechanism and pathway of water absorption by roots is
more complex than in the case of a single cell (Section ‘‘Water Relations of Plant Cells’’).
Water must travel first radially from the epidermis to cortex, endodermis, and pericycle
before it finally reaches the xylem vessels, from which point water flow is axial along
Francisco Pugnaire/Functional Plant Ecology 7488_C006 Final Proof page 186 10.5.2007 2:47pm Compositor Name: VBalamugundan
186 Functional Plant Ecology
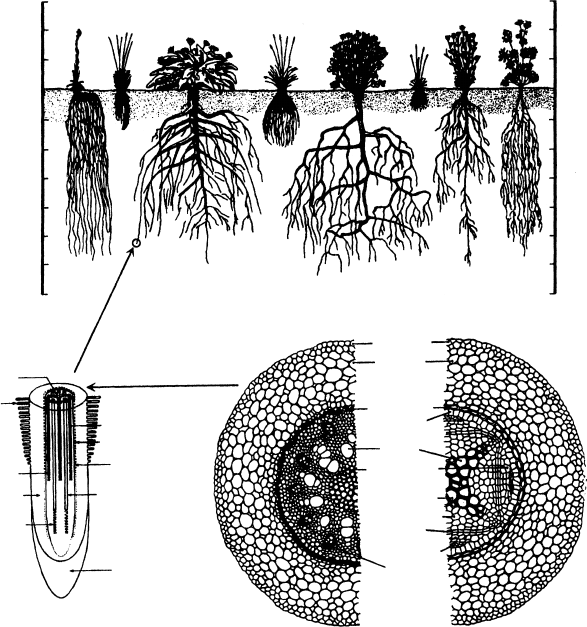
the root (Figures 6.6b and 6.6c). The radial pathway (typically 0.3 mm long in young roots) is
usually much less conductive than the axial pathway (>1 m in many cases); therefore,
whole-root conductance is generally proportional to the root surface area. The radial
pathway can be viewed as a composite membrane separating the soil solution from the
solution in the xylem fluid. The composite membrane consists of serial and parallel pathways
made up of plasmalemma membranes, cell wall ‘‘membranes,’’ and plasmodesmata
(pores <0.5 mm diameter) that connect adjacent cells. The composite membrane is rather
leaky to solutes; therefore, differences in osmotic potential between the soil (p
s
) and the xylem
(p
x
) have less influence on the movement of water. At any given point along the axis, the
water flux density across the root radius (J
r
) is given by
J
r
¼ L
r
[(P
s
P
x
) þ s(p
s
p
x
)], (6:10)
m
h
k
b
fg
p
ho
po
(a)
(c)
(b)
0.6
0.0
0.6
1.2
1.8
Immature
metaxylem
Immature
protoxylem
Immature
protophloem
Root hair
Root cap
Protophloem
Pericycle
Protoxylem
Epidermis
Cortex
Endodermis
Pericycle
Xylem
Phloem
Monocot Dicot
Parenchyma
Primary xylem
Secondary
xylem
Cortex
Endodermis
FIGURE 6.6 (a) Differences in root morphology and depth of root systems of various species of prairie
plants growing in a deep, well-aerated soil. Species shown are: h, Hieracium scouleri;k,Loeleria cristata;
b, Balsamina sgittata;f,Festuca ovina ingrata;g,Geranium viscosissimum;p,Poa sandbergii; ho,
Hoorebekia racemosa; and po, Potentilla blaschkeana. (b) Enlargement of a dicot root tip enlarged
approximately 50. (c) Cross section of monocot and dicot roots enlarged approximately 400.
(Adapted from Kramer, P.J., Water Relations of Plants, Academic Press, New York, 1983; Steward,
F.C., Plants at Work, Addison-Wesley, Reading, MA, 1964; Bidwell, R.G.S., Plant Physiology,
Macmillan, New York, 1979.)
Francisco Pugnaire/Functional Plant Ecology 7488_C006 Final Proof page 187 10.5.2007 2:47pm Compositor Name: VBalamugundan
Water Relations and Hydraulic Architecture 187
where L
r
is the radial root conductance to water and s is the solute reflection coefficient. For
an ideal membrane in which water but not solutes may pass, s ¼1. For the composite
membrane of roots, s is usually between 0.1 and 0.8. The system of equations that describes
water transport in roots is complex when all of the factors are taken into account, for
example, axial and radial conductances, the fact that each solute has a different s, and that
the rate of water flow is influenced by the solute loading rate (J
s
). Water and solute flow in
roots can be described by a standing gradient osmotic flow model (readers interested in the
details may consult Tyree et al. (1994b) and Steudle (1992).
Fortunately, the equations describing water flow become simple when the rate of water
flow is high. The concentration of solutes in the xylem fluid is equal to the ratio of solute flux
to water flux (J
s
:J
w
) during steady-state flux. Solute flux tend to be more or less constant with
time, but water flux increases with increasing transpiration. When water flow is high, the
concentration of solutes in the xylem fluid becomes small and approaches values comparable
to that in the soil solution, and pressure differences become quite large, hence (P
s
P
x
) >>
(p
s
p
x
). Only at night or during rainy periods can values of (P
s
P
x
) approach those of
s(p
s
p
x
). Therefore, water flow (J
w
,kgs
1
) through a whole-root system during the day can
be approximated by
J
w
ffi k
r
(P
s
P
x,b
)(6:11)
where P
x,b
is the xylem pressure at the base of the plant and k
r
is the total root conductance
(combined radial and axial conductances).
HYDRAULIC ARCHITECTURE AND PATHWAY OF WATER
MOVEMENT IN PLANTS
Van den Honert (1948) quantified water flow in plants in a classical paper in which he viewed
the flow of water in the plant as a catenary process, where each catena element is viewed as a
hydraulic conductance (analogous to an electrical conductance) across which water (analogous
to electric current) flows. Thus, van den Honert proposed an Ohm’s law analog for water flow
in plants. The Ohm’s analog leads to the following predictions; (1) the driving force of sap
ascent is a continuous decrease in P
x
in the direction of sap flow; and (2) evaporative flux
density from leaves (E) is proportional to negative of the pressure gradient (dP
x
=dx) at any
given point (cross section) along the transpiration gradient (dP
x
=dx) at any given point
(cross section) along the transpiration stream. Thus, at any given point of a root, stem, or leaf
vein, we have
dP
x
=dx ¼ AE=K
h
þ rg dh=dx, (6:12)
where A is the leaf-area supplied water by a stem segment with hydraulic conductivity K
h
and rg dh=dx is the gravitational potential gradient, where r is density of water, g is acceler-
ation due to gravity, and dh=dx is height gained, dh, per unit distance, dx, traveled by water in
the stem segment.
In the context of stem segments of length (L) with finite pressure drops across ends of the
segment, we have
DP
x
¼ LAE=K
h
þ rgDh: (6:13)
Figure 6.7 illustrates water flow through a plant represented by a linear catena of conductance
elements near the center and a branched catena of conductance elements on the left. The
number and arrangement of catena elements are dictated primarily by the spatial precision
Francisco Pugnaire/Functional Plant Ecology 7488_C006 Final Proof page 188 10.5.2007 2:47pm Compositor Name: VBalamugundan
188 Functional Plant Ecology
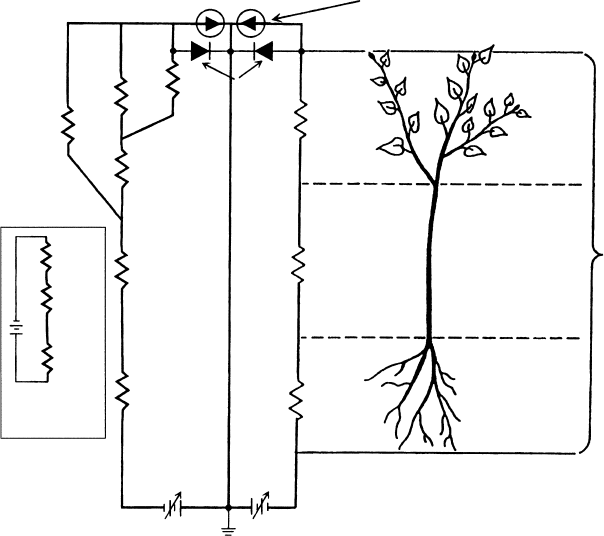
desired in the representation of water flow through a plant; a plant can be represented by
anywhere from one to thousands of conductance elements. Reviews of the hydraulic archi-
tecture of woody plants can be found in a report by Tyree and Ewers (1991 and 1996).
The usual way of representing the Ohm’s law circuit is incomplete. The usual but
incomplete representation of the Ohm’s law circuit is shown in the boxed inset in the lower
left side of Figure 6.7. The battery represents the water potential drop from root to leaf, but it
is an incomplete representation of reality because no ground point is shown in the circuit. The
ground point represents zero water potential. Water potential is always measured relative to
pure water at the same temperature and pressure as the plant. The incomplete representation
gives the correct drop in water potential across each conductance element but not the correct
water potential relative to ground (the zero reference). The complete representation has a
variable battery that gives the soil water potential (C
soil
) and has some analog components to
represent the evaporation from the leaves. In Figure 6.8, the evaporation is shown by a constant
current source and a rectifier in parallel. The amount of current, E, in the constant current
source can be approximated by Equation 6.8. When E ¼0 we expect leaf water potential to
equal soil water potential and that there should be no water current flowing through the
circuit; this condition is achieved in the circuit by the introduction of the rectifier that
prevents backward current flow when E ¼0andC
soil
< 0. In some experimental conditions,
C
soil
can be raised to positive values by placing a potted root system in a pressure chamber with
Branched model
Usual
incomplete
representation
−−++
K
leaf 2
K
leaf1
K
leaf
K
stem 2
K
stem1
K
stem
K
liquid
K
stem
K
root
K
root
⌿
soil
⌿
soil
=
⌿
root
=
⌿
leaf
=
⌿
soil
K
root
K
leaf 3
K
leaf
Rectifier
−0.7 (MPa)
−0.2 (MPa)
−0.05 (MPa)
Linear
model
(constant current source)
E
= g
L
∆X
FIGURE 6.7 The Ohm’s law analogy. The total conductance is seen as resultant conductance (K ) of the
root, stem, and leaf in series and parallel. Water flow is driven by evaporation of water from leaves,
which creates a difference in water potential between the soil, C
soil
, and the water potential at the
evaporating surface, C
evap
. On the right is the simplest Ohm’s law analogy with conductances in series.
On the left is a more complex conductance catena in which some conductance elements are in series and
some in parallel.
Francisco Pugnaire/Functional Plant Ecology 7488_C006 Final Proof page 189 10.5.2007 2:47pm Compositor Name: VBalamugundan
Water Relations and Hydraulic Architecture 189