Хант Р.В.Г. Цветовоспроизведение
Подождите немного. Документ загружается.


Цветовое восприятие предметных стимулов скорее относительно, чем абсолютно, к
примеру, если на некую серую поверхность сначала посмотреть при ярком солнечном
свете (освещенность порядка 50000 лк), а затем при лампе искусственного дневного
света (освещенность порядка 50 лк), то абсолютная фотометрическая яркость этой по
верхности уменьшится в 1000 раз, и, хотя поверхность субъективно окажется гораздо
менее яркой, то же коснется и прочих объектов поля зрения. В результате предмет так
и останется серым, то есть величина визуальной «серости» в целом будет неизменной.
Сие свойство восприятия необходимо человеческому зрению для оценки значимых
стимулов относительно стимулов, исходящих от прочих объектов поля зрения. Одна
ко, поскольку в большинстве обычных просмотровых ситуаций поверхностные стиму
лы лишь весьма малого диапазона яркостей воспринимаются как белые (стимулы
бо<льших яркостей выглядят самосветящимися, меньших — светлосерыми), можно
сделать упрощение, предположив, что цветовая оценка стимулов осуществляется зре
нием относительно белого (что и предполагается колориметрическим цветовоспроиз
ведением) (Evans, 1959).
В середине XX века было обнаружено (Hunt, Pitt и Ward, 1969), что, оценивая ре
зультат воспроизведения серого именно относительно белого, можно уйти от многих
сложностей, возникающих при необходимости воспроизведения по восприятию (см.
гл. 6). К примеру, светлосерый объект на солнечном свету дает стимул такой субъек
тивной яркости, что уравнять его на отпечатке, предназначенном к рассматриванию
при искусственном освещении, совершенно невозможно. Однако если на отпечатке
211
ГЛАВА 11 УРОВНИ ЦВЕТОВОСПРОИЗВЕДЕНИЯ
ТАБЛИЦА 11.1 Типичные уровни освещенности
ЕСТЕСТВЕННОЕ ОСВЕЩЕНИЕ
Яркое солнце от 50 000 до 100 000 лк
Солнце в дымке от 25 000 до 50 000 лк
Высокая облачность от 10 000 до 25 000 лк
Средняя облачность от 2000 до 10 000 лк
Низкая облачность от 100 до 2000 лк
Закат от 1 до 100 лк
Полная луна от 0.01 до 0.1 лк
Свет звезд от 0.0001 до 0.001 лк
ИСКУССТВЕННОЕ ОСВЕЩЕНИЕ
Операционная от 5000 до 10 000 лк
Витрины магазинов от 1000 до 5000 лк
Чертежные бюро от 300 до 500 лк
Офисы от 200 до 300 лк
Жилые помещения от 50 до 200 лк
Коридоры от 50 до 100 лк
Хорошее уличное освещение 20 лк
Плохое уличное освещение 0.1 лк

объект будет воспроизведен как светлосерый, то такая тоновая коррекция даст впол
не удовлетворительный результат.
Та же проблема возникает и в отношении полноты цвета изображений объектов:
к примеру, если при солнечном свете на зрительную систему воздействуют некие сти
мулы высокой колориметрической чистоты, то вызванные ими ощущения будут обла
дать большей полнотой цвета, чем даже ощущения от монохроматических стимулов,
но предъявленных при типичных уровнях искусственного освещения (Hunt, 1953).
Казалось бы, по этой причине выполнить удовлетворительную цветовую репродукцию
солнечных сцен, предназначенную к просмотру при искусственном освещении, — не
возможно, — однако, как мы знаем, опыт свидетельствует об обратном.
Действительно, существуют все основания строить цветовоспроизведение, оттал
киваясь от белого, и, следовательно, фотометрия яркостей относительно ярко осве
щенного белого эталона вполне оправданна. Итак: равенство относительных ярко
стей и равенство относительных цветностей мы и называем колориметрическим цве
товоспроизведением.
Таким образом, достижение уровня колориметрического цветовоспроизведения —
это хороший критерий качества отражающих отпечатков (при условии, что их окру
жение сходно с окружением оригинала), рассматриваемых при свете той же цветно
сти, но обычно меньшей интенсивности, чем в оригинальной сцене. Однако нельзя за
бывать, что, поскольку субъективная яркость и полнота цвета меняются при смене
уровня освещенности, всегда можно «поймать» тот ее уровень, при котором репродук
ция будет максимально близка к оригиналу по восприятию.
11.6 ТОЧНОЕ ЦВЕТОВОСПРОИЗВЕДЕНИЕ
Когда в дополнение к равенству по цветностям и фотометрическим яркостям рав
ными оказываются и абсолютные фотометрические яркости объектов оригинальной
сцены и элементов ее репродукции, мы говорим о ситуации, при которой исключено
отличие в освещенности обеих (или, в случае телевидения, — в экранной яркости). Сие
представляет собой т.н. точное цветовоспроизведение. Следовательно:
ð
Точное цветовоспроизведение — это процесс воспроизведения цветовых стиму
лов с теми же цветностями, относительными и абсолютными фотометрическими
яркостями, что и у стимулов оригинальной сцены.
Результатом точного цветовоспроизведения является равенство трихроматиче
ских воздействий стимулов оригинала и его копии на зрительную систему наблюдате
ля, а также равенство ощущений, вызванных этими воздействиями. Последнее обес
печено равенством адаптаций зрительной системы в условиях просмотра оригиналь
ной сцены и в условиях просмотра ее изображения (факторами, оказывающими основ
ное влияние на адаптацию зрения, являются: фотометрическая яркость и цветность
окружения стимула, угловой размер области стимуляции и паразитная подсветка;
если все перечисленные условия одинаковы — адаптация будет одна и та же).
Таким образом, если воспроизведение данного зрительного стимула является точ
ным, то, при одновременном соблюдении всех наиболее важных условий просмотра,
результат цветового восприятия этого стимула наблюдателем будет ровно таким же,
как результат восприятия соответствующего стимула оригинальной сцены.
Отличия же в восприятии возникнут в случаях, когда:
— условия просмотра оригинала и его репродукции различны;
212
ГЛАВА 11 УРОВНИ ЦВЕТОВОСПРОИЗВЕДЕНИЯ

— фактический наблюдатель ощутимо отличен от Стандартного колориметриче
ского двухградусного наблюдателя CIE;
— спектральное распределение энергии осветителей несколько отличается от того,
что было использовано при расчете цветностей и относительных фотометрических яр
костей стимулов (то есть, когда имели место колориметрические погрешности).
ð
Точное цветовоспроизведение — это прерогатива и цель абстрактных цветовос
производящих систем и цветопробных процессов (см. раздел 25.9).
1
11.7 ЭКВИВАЛЕНТНОЕ ЦВЕТОВОСПРОИЗВЕДЕНИЕ
Существует множество ситуаций, при которых достижение уровня колориметри
ческого или точного цветовоспроизведения было бы ошибкой. К примеру, когда сцена,
освещенная лампами накаливания, воспроизводится в расчете на просмотр при днев
ном освещении, то и колориметрическое, и точное цветовоспроизведения приведут
к выраженному сдвигу общего хроматического баланса репродукции в желтую сторо
ну. Такая ситуация обычно возникает в телевидении: если студийная сцена, освещен
ная лампами накаливания, воспроизводится экранами телеприемников на колори
метрическом или точном уровнях, то при дневном просмотре она выглядит желтой.
Так происходит изза того, что зрительная система адаптирована в основном к дневно
му свету, поскольку он занимает бо<льшую часть поля зрения, тогда как в оригиналь
ной сцене зрение адаптировано к свету ламп накаливания, и, следовательно, по отно
шению к средневолновой («зеленой») чувствительности его коротковолновая («си
няя») чувствительность повышена, а длинноволновая («красная») — понижена. (Оп
тимальным хроматическим балансом для телевизионных передач является баланс по
смешанному освещению, о чем говорится в разделе 21.13).
Поскольку на цветовое восприятие световых потоков влияют параметры просмот
ра, в частности описанные выше, необходимо позиционировать четвертый уро
вень цветовоспроизведения — уровень эквивалентный.
ð
Эквивалентное цветовоспроизведение — это процесс воспроизведения цветовых
стимулов, при котором их цветности, относительные и абсолютные фотометри
ческие яркости таковы, что в репродукционных условиях просмотра результат
цветового восприятия этих стимулов идентичен результату цветового воспри
ятия стимулов оригинальной сцены.
Существует как минимум три варианта обстоятельств, которые в данной связи
имеют прикладное значение:
— отличия в цветностях оригинального и репродукционного осветителей;
— отличия в интенсивностях оригинального и репродукционного осветителей;
— отличия в окружениях оригинальной сцены и ее изображения.
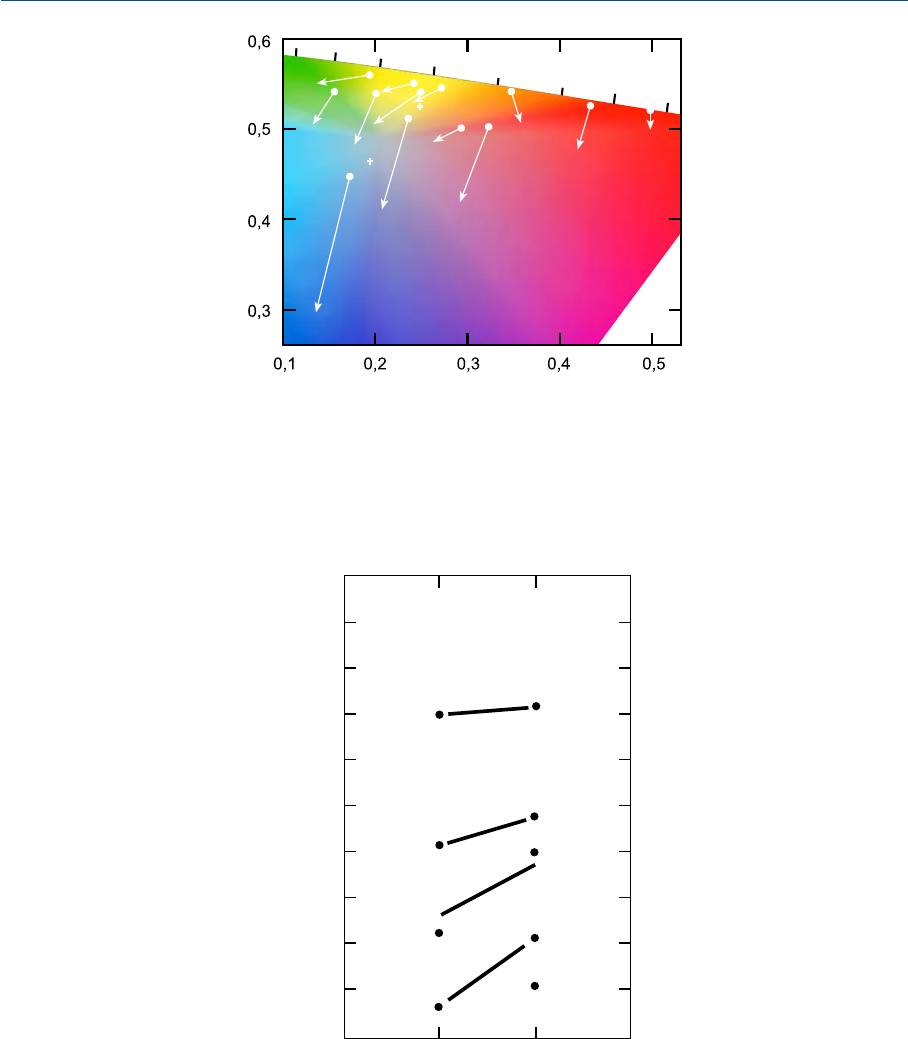
Проиллюстрируем сказанное на примере результатов экспериментов по т.н. гапло
скопическому уравниванию (см. раздел 8.10).
На рис. 11.3 даны цветности нескольких пар физически разных стимулов, вызы
вающих одинаковые цветовые ощущения в той ситуации, когда стимулы (цветности
213
ГЛАВА 11 УРОВНИ ЦВЕТОВОСПРОИЗВЕДЕНИЯ
1
Под «абстрактными цветовоспроизводящими системами» здесь подразумеваются системы
оцифровки физических оригиналов; результатом работы этих систем является файл, содержа
щий данные о цветностях и фотометрических яркостях стимулов сцены. — Прим. пер.

которых отмечены точками) предъявлены в условиях адаптации по свету ламп нака
ливания (правый глаз), а эквивалентные им стимулы (на цветности которых указыва
ют острия стрелок) — при адаптации по дневному свету (левый глаз). Мы видим, что
стимул, который в условиях дневной адаптации вызовет точно такое же цветовое ощу
щение как некий другой стимул, предъявленный в условиях адаптации к свету ламп
накаливания, должен содержать большее количество коротковолновой энергии (то
есть быть синее при сравнении обоих стимулов в полной темноте). Такие стимулы име
нуются эквивалентными стимулами. Отметим, что в свое время были предложены
уравнения расчета трехстимульных значений эквивалентных цветовых стимулов
(Burnham, Evans и Newhall, 1957; Nayatani, Takahama и Sobagaki, 1981; Hunt, 1998;
см. гл. 34).
На рис. 8.20 даны цветности нескольких эквивалентных стимулов при вариантах
адаптации по освещениям одинаковой цветности, но разной интенсивности (Hunt,
1952 и 1953): мы видим, что по мере снижения интенсивности освещения и, следова
тельно, по мере снижения интенсивности тестовых стимулов снижается полнота цвета
последних.
На рис. 13.7 показаны фотометрические яркости эквивалентных серых стимулов
(от белого до черного), предъявленных в определенном диапазоне величин освещен
ности сцены (Hunt, 1965a): мы видим, что по мере снижения освещенности и вызван
ного им снижения интенсивности исследуемых предметных стимулов, понижается
субъективная яркость последних, а также слегка падает визуальный контраст меж
ду стимулами (последнее иногда относят на счет нелинейности шкалы ординат, но
ряд исследователей считает, что снижение контраста все же имеет место [Bartleson и
Breneman, 1967 a]).
На рис. 11.4 дан пример влияния окружения: эквивалентные стимулы подобраны
по серой шкале, предъявленной наблюдателю сначала на сером, а затем на черном фо
нах. Мы видим, что (как отмечалось в гл. 6) темное окружение ведет к повышению ви
зуальной гамма (Hunt, 1965 b). Напомним при этом, что оно же может понизить полно
ту цвета элементов изображения (Hunt, 1950; Rowe, 1972; Hunt, 1973; Pitt и Winter,
1974; Breneman, 1977). Любопытно, что коррекция изображения, необходимая для
компенсации падения визуального контраста и полноты цвета при тусклом или тем
ном окружениях, в восприятии самого цветокорректора ведет к усилению тоновых и
хроматических отличий репродукции от оригинала.
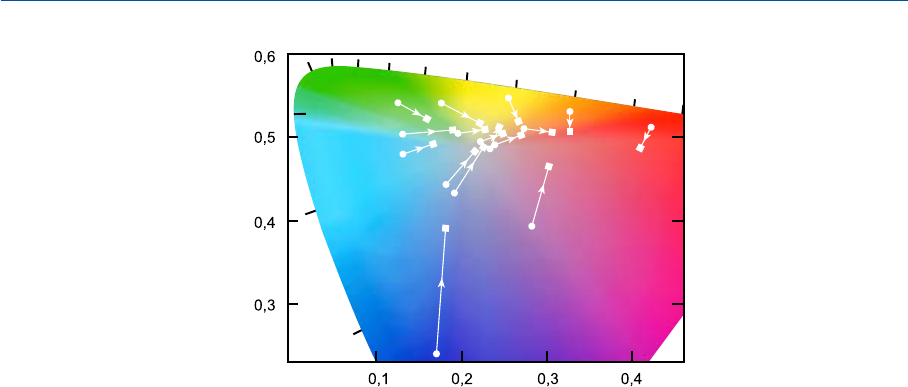
На рис. 11.5 дан пример влияния изменений в цветности и интенсивности освеще
ния и в окружающих стимулах: эквивалентные стимулы были выявлены с помощью
специальной тесткарты при ярком солнечном свете и при свете проектора с лампой
накаливания; последнее имитировало проекционный просмотр изображения в тем
ном окружении (Hunt, 1965 b). Понятно, что цветовое восприятие стимулов в обоих
случаях было весьма различным, а ослабление света проектора вело к снижению пол
ноты цвета образцов карты с одновременным хроматическим сдвигом в желто
оранжевую сторону. (Однако результаты данного эксперимента зависели в том числе
и от специфического спектрального коэффициента отражения образцов: метамерные
патчи карты, то есть образцы, имевшие одинаковые цветности на солнечном свету, но
при этом различные кривые спектрального коэффициента отражения, продемонстри
ровали различную степень хроматического сдвига при переходе к свету проектора и,
следовательно, имели иные эквивалентные стимулы. Вместе с тем общее направление
изменений было таким же.)
Забегая вперед, скажем, что описанные выше эффекты можно спрогнозировать с по
214
ГЛАВА 11 УРОВНИ ЦВЕТОВОСПРОИЗВЕДЕНИЯ
215
ГЛАВА 11 УРОВНИ ЦВЕТОВОСПРОИЗВЕДЕНИЯ
600
550
560
570
580
590
610
620
v'
u'
S
A
S
C
Рис. 11.3 Цветности стимулов, воспринимаемых одинаково при равных фотометрических яр
костях адаптирующих полей (8.1 кд/м
2
). При этом цветность правого адаптирующего поля
(правый глаз) равна цветности Стандартного Аосветителя (цветности стимулов отмечены точ
ками); цветность левого адаптирующего поля (левый глаз) равна цветности Стандартного осве
тителя С (цветности стимулов отмечены остриями стрелок).
160 ëê
ïðè ñåðîì
îêðóæåíèè
òåñòîâûõ
ñòèìóëîâ
0
20
40
60
80
100
L*
4
5
4
3
2
1
3
2
1
160 ëê
ïðè òåìíîì
îêðóæåíèè
òåñòîâûõ
ñòèìóëîâ
Рис. 11.4 Фотометрические яркости серых стимулов (отложены по CIE L
*
шкале), подобран
ные для правого глаза, при его адаптации по полю с яркостью 3600 кд/м
2
и коррелированной
цветовой температурой 4000 К, с целью достижения визуальной эквивалентности нескольким
серым стимулам в среднем (левая колонка) и темном (правая колонка) окружениях, последова
тельно предъявленным левому глазу, адаптированному к свету лампы накаливания при осве
щенности сцены 160 лк; благодаря темному окружению темносерые образцы воспринимают
ся светлее, что снижает общий визуальный контраст серии.
216
ГЛАВА 11 УРОВНИ ЦВЕТОВОСПРОИЗВЕДЕНИЯ
v'
u'
600
550
500
560
570
580
590
Ïóðïóðíûé
Îðàíæåâûé
Êðàñíûé
Æåëòûé
Æåëòî-çåëåíûé
Çåëåíûé
Ñâåòëî-çåëåíûé
Ãîëóáîé
Íåáåñíî-ñèíèé
Êîæà
Ïåðñèêîâûé
Áëåäíî-ñèíèé
Ñèíèé
Ëèñòâà
Рис. 11.5 ¡ — цветности стимулов, подобранных для правого глаза при его адаптации по полю с
фотометрической яркостью 3600 кд/м
2
и коррелированной цветовой температурой 4000Ксце
лью достижения визуальной эквивалентности 16ти разным стимулам, последовательно
предъявленным левому глазу при ярком солнечном свете с освещенностью сцены в 43000 лк;
£ — цветности стимулов, подобранных для правого глаза (при тех же условиях его адаптации)
с целью достижения визуальной эквивалентности 16ти разным стимулам, последовательно
предъявленным левому глазу при его адаптации по свету лампы накаливания (проектор) с ос
вещенностью сцены 160 лк и темном окружении; стрелки указывают на снижение полноты
цвета стимулов, предъявляемых левому глазу при переходе к тусклому свету проектора, и од
новременное смещение их цветового тона в желтооранжевую сторону.
Сопутствующие изменения в светлоте стимулов:
Ощущение светлоты
Цветовой стимул Яркое солнце Свет проектора
Белый 87 64
Желтый 79 75
Телесный 72 53
Песочный 68 43
Оранжевый 61 36
Светлозеленый 61 32
Желтозеленый 63 36
Персиковый 56 42
Небесносиний 56 32
Бледносиний 58 35
Цвета листвы 53 28
Красный 44 22
Пурпурный 40 19
Зеленый 41 13
Голубой 30 9
Синий 28 10

мощью т.н. моделей цветового восприятия, в частности CIECAM97s и CIECAM02 (см. гл.
35 и Приложение 6).
ð
Эквивалентное цветовоспроизведение — это прерогатива и цель передачи изо
бражений по интернет.
11.8 СОГЛАСОВАННОЕ ЦВЕТОВОСПРОИЗВЕДЕНИЕ
Дабы справиться с трудностями, возникающими на пути к колориметрическому и
эквивалентному уровням (то есть трудностями, связанными с изменчивостью субъек
тивной яркости и полноты цвета), нам следует обратиться к концепции т.н. согласо
ванного цветовоспроизведения:
ð
Согласованное цветовоспроизведение — это процесс воспроизведения цветовых
стимулов, при котором их цветности и относительные фотометрические яркости
таковы, что результат цветового восприятия этих стимулов оказывается тем же,
что и у стимулов оригинальных, но при условии равенства освещенностей и/или
интегральных яркостей оригинала и репродукции.
Учтя отличия в средних уровнях абсолютной фотометрической яркости оригинала
и репродукции, как, скажем, в случае колориметрического цветовоспроизведения,
мы уйдем от парадоксального вывода о невозможности воспроизведения изображений
ярко освещенных сцен при низких уровнях освещенности отражающих репродукций
(низких уровнях яркости телеизображений). Однако требование равенства по воспри
ятию требует учета влияния на это равенство отличий в окружении и в цветности осве
щения. Пути учета влияния среднего, тусклого и темного окружений на восприятие
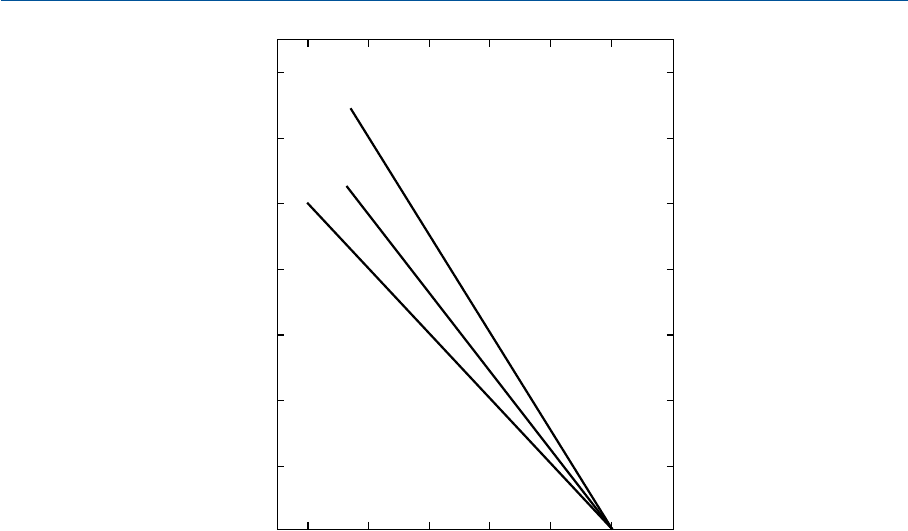
белого, серого и черного стимулов показаны на рис. 11.6, взятом из гл. 6: оптические
плотности элементов репродукции, необходимые для достижения согласованного цве
товоспроизведения, отложены по оси ординат, а логарифмы относительных фотомет
рических яркостей объектов оригинальной сцены (Bartleson и Breneman, 1967 b) — по
оси абсцисс (Hunt, 1969).
Как уже было сказано в гл. 6, среднее окружение — это типичное окружение отра
жающих отпечатков; тусклое окружение — типичное окружение телевизионного про
смотра или просмотра листовых слайдов в просмотровых кабинах; темное окруже
ние — окружение проекционного показа слайдов в затемненном помещении. В послед
них двух случаях величина гамма должна быть около 1.25 и 1.5 соответственно, по
скольку тусклое и темное окружения понижают визуальную контрастность изображе
ний. Внешнее освещение при телевизионном просмотре, как правило, изменчиво, но
замечено, что когда дисплей телеприемника имеет гамма показа 1.5, то паразитная
подсветка, исходящая от внешнего освещения обычного уровня, понижает гамма до
1.25, что и требуется (Novick, 1969) (см. раздел 19.13).
Напомним, что по мере роста гамма очень кстати растет и колориметрическая
чистота элементов репродукции, поскольку высокая гамма усиливает разброс между
их трехстимульными значениями (MacAdam, 1938); несмотря на то что темное или
тусклое окружения слегка понижают эту чистоту, имеет место ее общий рост (см.
раздел 7.9).
И снова забегая вперед, скажем, что с целью учета влияния на цветовое восприятие
изменений в цветности освещения можно использовать т.н. расчет смены хроматиче
217
ГЛАВА 11 УРОВНИ ЦВЕТОВОСПРОИЗВЕДЕНИЯ
ской адаптации (Chromatic Adaptation Transform — CAT), позволяющий вычислять
трехстимульные значения т.н. согласованных стимулов (см. гл. 34).
ð
Согласованные цветовые стимулы — это физически разные световые потоки, с
разными трехстимульными значениями, которые при адаптации зрения по ос
вещениям разной цветности, но равной интенсивности воспринимаются одина
ково.
Расчет смены хроматической адаптации, в частности, учитывает тот факт, что
цветность освещения далеко не всегда соответствует цветности белой точки, к приме
ру: когда в затемненном помещении с помощью проектора с лампой накаливания де
монстрируются слайды, свет от экрана воспринимается слегка желтоватым, причем
даже тогда, когда наблюдатель, казалось бы, адаптирован к нему. Следовательно, для
создания ощущения белого нужный участок слайда должен быть слегка «синеватым»
(Hunt, 1965b; Раздел 5.7). Однако напомним, что адаптация бывает полной по
среднесерому и избыточной по темносерому адаптирующим стимулам (Helson, 1938;
Judd, 1940). Далее об этом см. в гл. 34, 35 и 36.
ð
Концепция согласованного цветовоспроизведения, пожалуй, более всего подхо
дит решению общих цветорепродукционных задач.
Отметим также, что уровень согласованного цветовоспроизведения настолько же
выше уровня эквивалентного цветовоспроизведения, насколько уровень цветовоспро
218
ГЛАВА 11 УРОВНИ ЦВЕТОВОСПРОИЗВЕДЕНИЯ
012
0
1
2
3
Ñðåäíåå
îêðóæåíèå
g=1.0
Îïòè÷åñêàÿ ïëîòíîñòü ýëåìåíòîâ ðåïðîäóêöèè
Òåìíîå
îêðóæåíèå
g=1.5
Òóñêëîå
îêðóæåíèå
g=1.25
Ëîãàðèôì îòíîñèòåëüíîé
ôîòîìåòðè÷åñêîé ÿðêîñòè îáúåêòîâ ñöåíû
Рис. 11.6 Оптические плотности, необходимые для достижения уровня согласованного воспро
изведения белых, серых и черных стимулов при трех вариантах окружения изображений:
среднем (в основном отпечатки), тусклом (телепередача или листовые слайды в просмотровой
кабине) и темном (проекционный показ слайдов в затемненном помещении).

изведения колориметрического выше уровня точного цветовоспроизведения, по
скольку, относя стимулы оригинала и репродукции к белому эталону, мы учитываем
склонность зрения к неизолированному восприятию этих стимулов, то есть воспри
ятию с опорой на некий зрительный эталон, задаваемый условиями просмотра.
Таким образом, когда, к примеру, изображение солнечной сцены воспроизводится
на эквивалентном уровне, скажем, путем проекции слайда светом лампы накаливания
в темном окружении, колориметрическая чистота эквивалентных стимулов должна
быть очень высокой, дабы стимулы эти вызывали ощущения высокой полноты цвета
объектов этой сцены. Однако мы знаем, что в затемненной обстановке проекционного
показа полнота всех цветовых ощущений снижается и изображение выглядит менее ес
тественным.
Численное описание отмеченных выше эффектов требует углубленного исследова
ние, и нельзя забывать, что (как и в случае колориметрического воспроизведения) изо
бражение будет тем более похожим на оригинал, чем ближе освещенность репродук
ции к освещенности оригинальной сцены. Таким образом, качество экранного изобра
жения солнечной сцены можно повысить за счет повышения экранной яркости стиму
лов, полученных от слайда (Bartleson, 1965). Отпечатки солнечных сцен тоже смот
рятся лучше при ярком свете.
Аналогичные эффекты возникают и в телевидении: с внедрением зеленых люмино
форов на основе сульфида цинка (см. раздел 21.12) ощутимо снизилась колориметриче
ская чистота зеленого кардинального стимула, что привело к снижению качества коло
риметрического цветовоспроизведения. Однако, поскольку зеленый сульфидный лю
минофор позволяет воспроизводить изображения с большей абсолютной фотометриче
ской яркостью — снижение колориметрической чистоты, если угодно, автоматически
компенсируется. В итоге, общее качество телеизображений выросло (Matthews, 1963).
Отметим, что в данном случае согласованное цветовоспроизведение уступает позиции
воспроизведению эквивалентному.
11.9 ВЫДЕЛЕННОЕ ЦВЕТОВОСПРОИЗВЕДЕНИЕ
К рассуждениям об уровнях цветовоспроизведения, очевидно, необходимо доба
вить тот факт, что кожа европеоида на репродукции выглядит реалистичнее, когда пе
редана как загорелая (MacAdam, 1951; Bartleson и Bray, 1962). Существует множество
цветов, к которым применимы аналогичные поправки, к примеру, синие небо и вод
ная гладь, радующие глаз на отпечатках или слайдах, в реальных сценах могут выгля
деть почти серыми: дело в том, что фотопленки обладают некоторой чувствительно
стью к ультрафиолетовому излучению и, следовательно, повышают синеву неба и во
ды относительно чистоты цвета прочих элементов репродукции. Однако, когда такое
повышение не чрезмерно, оно предпочтительно для большинства изображений неба
и воды. Более того, и некоторые другие цветоискажения могут оказаться желательны
ми при создании определенного настроения и атмосферы снимков.
Сказанное имеет очень большое прикладное значение, но вместе тем означает, что
концепции спектрального, колориметрического, точного, эквивалентного и согласо
ванного цветовоспроизведений — это базис, на котором строится разговор о намерен
ных цветовых искажениях в репродукции. То есть, мы говорим о т.н. выделенном цве
товоспроизведении.
219
ГЛАВА 11 УРОВНИ ЦВЕТОВОСПРОИЗВЕДЕНИЯ
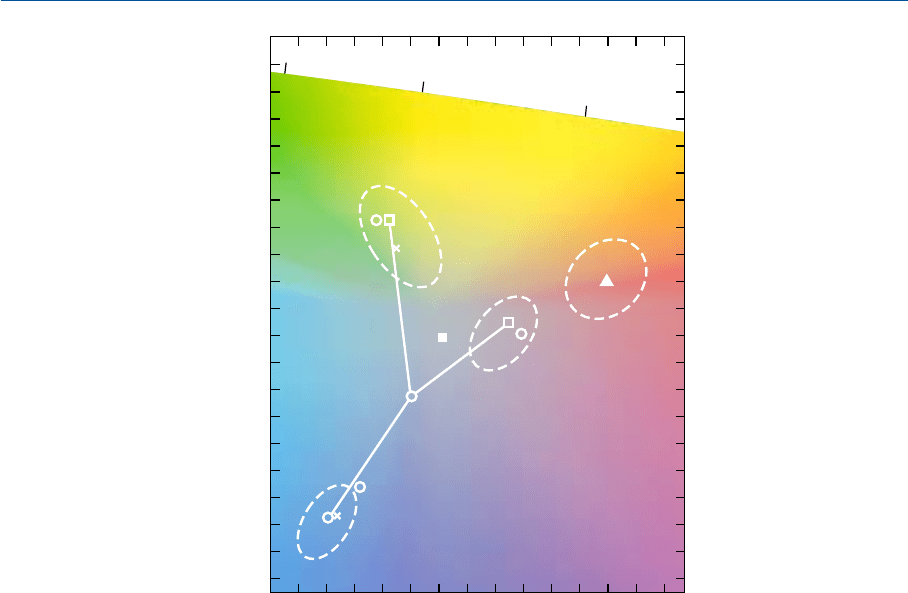
ð
Выделенное цветовоспроизведение — это процесс воспроизведения цветовых сти
мулов, которые во имя достижения максимального комфорта их визуального
восприятия отклоняются как от абсолютного, так и от относительного равенства
по восприятию со стимулами оригинала.
На рис. 11.7 и 11.8 даны цветности стимулов репродукции ясного неба, летней травы
и кожи европеоида, позволяющие достичь уровня выделенного цветовоспроизведения
(Hunt, Pitt и Winter, 1974). Приведенные данные получены из снимков уличных сцен, а
необходимый эффект достигнут путем применения пары непрозрачных масок: одной
затемнявшей интересующий участок изображений, другой — оставшуюся их часть.
220
ГЛАВА 11 УРОВНИ ЦВЕТОВОСПРОИЗВЕДЕНИЯ
0,15 0,20 0,25 0,30
0,40
0,45
0,50
0,55
560
570
580
Òðàâà
Êîæà
Íåáî
Êîæà íà ýêðàíå
òåëåâèçîðà
S
C
v'
u'
Рис. 11.7 ð — цветности репродукционных стимулов летней травы, ясного неба и типичной
кожи европеоида, обеспечивающих уровень выделенного цветовоспроизведения при печати на
отражение; ¡ — цветности стимулов, полученных от фактических летней травы, синего неба и
кожи европеоида; ´ — цветности репродукционных стимулов травы, ясного неба и типичной
кожи европеоида, воспроизеденных обычной негативнопозитивной фотографической систе
мой; во всех случаях — осветитель S
C
.
Дополнительно:
p — цветность репродукционного стимула кожи европеоида, обеспечивающего уровень
выделенного цветовоспроизведения телеэкраном (при тусклом просмотровом освещении лам
пами накаливания); ¢ — субъективно нейтральная точка телеэкрана.
Пунктирные линии отграничивают области цветностей стимулов, дающих приемлемый ре
зультат цветовоспроизведения. Относительные фотометрические яркости выделенных стиму
лов: трава — 27%; небо — 30%; кожа — 39%. Типичные величины относительных яркостей
реальных объектов: трава — 13%; кожа — 35%.