Санагурський Д.І. Об’єкти біофізики: монографія
Подождите немного. Документ загружается.

351
глутамінова та аспарагінова (збуджуючі амінокислоти); гама-
аміномасляна кислота (ГАМК) i гліцин (гальмівні амінокислоти).
Протягом тривалого часу вважали, що нервові закінчення одного й
того ж нейрона виділяють один i той самий нейромедіатор
(принцип Дейла). Але доведено, що нейрони, які розвиваються,
синтезують i виділяють більше, ніж один нейромедіатор. Разом з
низькомолекулярними медіаторами виділяються також нейро-
активні пептиди.
Взаємодія нейромедіатора з рецепторами постсинаптичної
мембрани приводить до виникнення постсинаптичного поте-ціалу.
Постсинаптичні потенціали можуть приводити як до деполяри-
зації, так i до гіперполяризації.
Потенціали, що зумовлюють деполяризацію є збуджуваль-
ними, а гіперполяризацію – гальмівними. Їх позначають від-
повідно ЗПСП та ГПСП (П.Г. Костюк, 1986).
Виникнення ЗПСП переважно пов’язане зі збільшенням
провідності постсинаптичної мембрани для йонів натрію, калію i,
можливо, кальцію, зумовлене активуванням йонних каналів, що
мають низьку селективність. Потенціал реверсії для синаптичного
струму, який виникає, дорівнює приблизно 0 мВ. Сумарний струм
через постсинаптичну мембрану направлений у клітину. Інколи
ЗПСП виникає в результаті зниження провідності мембрани для
йонів калію. Водночас взаємодія нейромедіатора з рецептором
приводить до появи всередині клітини вторинного посередника,
який зумовлює низку подій, унаслідок яких закриваються калієві
канали.
352
Нейромедіатори, що є причиною виникнення ГПСП,
активують хлорні або калієві канали. Потенціали реверсії для цих
іонів у нервових клітинах (Е
CI
i Е
к
) становлять віповідно –60 i
–70 мВ, а ПС – приблизно –50 мВ.
Активування як хлорних, так i калієвих каналів спричиняє
гіперполяризацію мембрани. Синаптичний струм під час ГПСП
протікає через активовані нейромедіатором канали з клітини.
Для вивчення властивостей іонних каналів, які
активуються нейромедіаторами, реєструють струми, що
протікають через поодинокі йoннi канали за умови фіксування
потенціалу на фрагменті мембрани (петч-клемп).
Іонний канал ГАМК
А
-рецептора є лігандзв’язаним
каналом, який активується гама-аміномасляною кислотою,
нейромедіатором у численних синапсах ЦНС ссавців. ГАМК
А
-
рецептор являє собою глікопротеїн, який складається з кількох
субодиниць п'ять з яких – це α, β ,γ, та ρ. ГАМК
А
-рецептор – це
йонний канал, який є вибірково проникним для йонів хлору.
Функцією ГАМК
А
-рецептора в ЦНС дорослих хребетних є швидке
гальмівне синаптичне передавання внаслідок зв’язування двох
молекул ГАМК i швидкого зростання проникності мембрани для
іонів хлору.
Слід зазначити, що ГАМК діє також через рецептори,
зв’язані з G-білками (ГАМК
В
-метаботропні рецептори) i ГАМК
С
-
рецептори, які є нечутливими до дії бікукуліну (йoнотpoпнi
рецептори) та проникними для аніонів хлору.
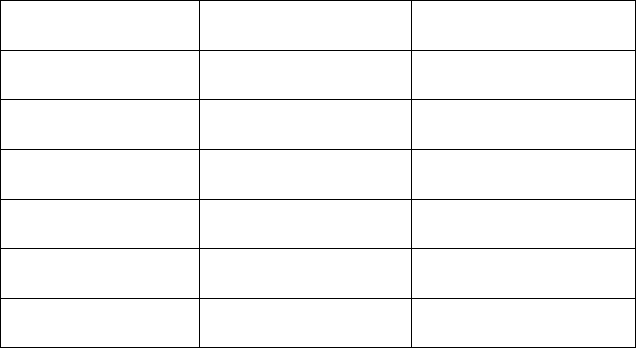
В таблиці 3.1 подано фармакологічні особливості ГАМК-
рецепторів.
353
Відомо, що серед похідних амінокислот ГАМК наявна у
значній кількості в пресинаптичних нервових закінченнях
хребетних та безхребетних тварин (Дж. Экклс, 1966; D. Belelli et al.,
1996; K.W. Gee, 1988; J.J. Lambert et al., 1995; G.G. Lunt,1991;
J.A. Peters et al., 1988). ГАМК збільшує проникність постсинаптич-
них мембран для К
+
, і, отже, віддаляє МП від порогового рівня,
при якому виникає ПД.
Таблиця 10.1
Фармакологія ГАМК-рецепторів
–
ГАМК
А
-рецептори ГАМК
В
-рецептори
Агоністи
ГАМК, мусцімол,
ізогуацин, ТНІР
ГАМК, баклофен
конкурентні
антагоністи
бікукулін факлофен
блокатори
каналів
пікротоксин –
алостеричні агоністи
бензодіазепін,
барбітурат
–
зворотні агоністи за
бензодіазепіном
бета-карболін (DMCM) –
антагоніст за
бензодіазепіном
Ro 15 1788 –
Примітка. ТНІР: 4, 5, 6, 7- тригідроізоксазолпіридин
ГАМК-рецептори відповідають за гальмування електричної
активності. Утворення ГАМК проходить при метаболічних
перетвореннях альфа-кетоглутарової кислоти в гама-амінобу-
тиратному шунті (ГАМК – H
2
NCH
2
-CH
2
-CH
2
-СООН).
Утворення ГАМК може здійснюватися також iз диаміна
путресцина (Н.Е. Кучеренко, 1988).
354
ГАМК перетворюється при декарбоксилюванні глютамату
в реакції, яка каталізується глютамат-декарбоксилазою
(M.G. Darlison, 1992). Простетичною групою цієї декарбоксилази
служить піридоксальфосфат.
Гама-амінобутират інактивується шляхом трансамінування
з утворенням півальдегіду бурштинової кислоти, який далі
окиснюється в сукцинат. Після перших доповідей у 1987 та 1989
роках про наявність ГАМК
А
-та глютаматних рецепторів на
ізольованих клітинах, виділених з мозку хребетних тварин,
з’явилася велика кількість публікацій, пов’заних з доповненнями у
послідовності кожного з поліпептидних ланцюгів, або відповідних
субодиниць, виявлених у інших хребетних.
Було описано 14 субодиниць ГАМК-рецепторів та 12 суб-
одиниць глютаматних рецепторів (H. Luddens, W. Wisden, 1991;
D. Wilson-Shaw et al., 1991).
У 1991 році було опубліковано дані про амінокислотну
послідовність двох різних поліпептидів ГАМК- та глютаматних
рецепторів (H. Luddens, W. Wisden, 1991; M.G. Darlison, 1992;
D. Wilson-Shaw et al., 1991). Експерименти, проведені на ізольо-
ваних нейронах Lymnea stagnalis та Drosophila melanogaster, дали
змогу описати структуру поліпептидів ГАМК
А
-рецепторів та
простежити за цією послідовністю в еволюційному аспекті.
Збуджуючі амінокислоти, такі як глютамат i аспартат,
містяться в багатьох синапсах центральної нервової системи
хребетних. За даними Kepтica (H.J. Curtis, K.S. Cole, 1942) глютамат
спричиняє деполяризацію в нейронах спинного мозку. Цей ефект є
355
швидким i може бути зумовлений також іншими амінокислотами
(L-аспартат, L-гомоцистат, L-цистеїн-сульфат).
Швидкість цього ефекту свідчить про те, що глютамат та
інші агоністи зумовлюють деполяризацію, діючи безпосередньо на
канальні рецептори. 3а допомогою методів петч-клемп
(конфігурація “outside-out”) було підтверджено цю гіпотезу.
Доведено, що глютамат ефективно діє навіть за браком будь-яких
внутрішньоклітинних компонентів (С. Hammond, 1996).
Виділяють дві ocновнi групи канальних рецепторів, які
активуються збуджуючими амінокислотами. Таке визначення
основане на спорідненості рецептора й глютаматних вибіркових
структур аналогів, особливо з N-метилом-D-аспартат (NMDA).
Отже, таке визначення зроблене між тими каналами глютаматних
рецепторів, які активуються NMDA-, й тими каналами
глютаматних рецепторів, які активуються АМРА, або каїнатом.
Слід зазначити, що глютамат діє також через рецептори,
що зв’язані з G-білками (глютаматні метаботропні рецептори,
mGluR). Дослідження, проведені на мозку щура та миші, дали
можливість ідентифікувати субодиниці АМРА-рецепторів,
названих GluR-1, GluR-2, GluR-3, GluR-4 та субодиниці каїнатних
рецепторів (GluR
5
, GluR
6
, GluR
7
та КА-1, КА-2) i субодиниці
NMDA-рецепторів (NR1, NR2).
За допомогою методу петч-клемп, досліджуючи in vitro
нейрони, які формують синапси спинного мозку та гіпокампа
хребетних тварин, інші вчені прийшли до висновку, що вхідні
постсинаптичні струми через NMDA-та нe-NMDA-рецептори, які

356
є результатом активування збуджуючих рецепторів, мають два
компоненти (С. Hammond, 1996). Зважаючи на властивості
NMDA- та не-NMDA- рецепторів (табл. 3.1), досліджено, що на
компонент не-NMDA-рецептора не впливають зміни
зовнішньоклітинної концентрації катіонів Mg
2+
, або дія APV.
Встановлено, що не-NMDA компонент зникає за наявності CNQX-
селективного антагоніста АМРА-рецепторів. 3 іншого боку, NMDA
компонент генерується у безмагнієвому середовищі та зникає під
впливом APV- конкурентного антагоніста NMDA-pецепторів.
Таблиця 10.2
Агоністи та антагоністи NMDA- та не-NMDA-рецепторів
не-NMDA канали
Рецептори
АМРА-рецептори Каїнатні
NMDA
канали
найбільш
селективні агоністи
АМРА, квісквалят каїнат NMDA
змішані
агоністи
глютамат,
аспартат
глютамат,
аспартат
глютамат,
аспартат
конкурентні
антагоністи
CNQX,
NBQX
CNQX,
NBQX
D-APV
(або DAPS)
CPP
блокатори
каналів
– – МК 801
неконкурентні
антагоністи за
гліциновим
сайтом
– –
7-хлор
кінуренат
Примітка. 1. АМРА – α-аміно-3-гідрокси-5-метил-4-ізоксалон пропіонат; 2. D-AP
5
або D-
APV – D-2-aміно-5-фосфонопентанат; 3. CNQX – 6-ціано-7-мітроквіноксалін-2,3-діон; 4.
NMDA – N-метил-D-аспартат; 5. МК801 – 5-метил-10,11-дигідро-5Н-дибензоциклогептен-
5,10-імін малат; 6. NBQX – 6-нітро-7-сульфамобензоквіноксалін-2,3-діон; 7. СРР – (2-
карбоксипіперазин-4-ил) пропіл-1-фосфонічно кислий
357
Таким чином, при стимулюванні нейрона у безмагнієвому
середовищі виникає вхідний струм, який має два компоненти:
перший компонент має велику амплітуду та швидко інактивується,
а другий – повільний компонент.
Наявність іонів Mg
2+
, або їxній брак, та наявність APV не
впливають на перший компонент, але блокують другий
(С. Hammond, 1996). Встановлено, що ранній компонент є
результатом синаптичного активування АМРА-рецепторів. Пізній
компонент спричинений активуванням NMDA-pецепторів,
оскільки за наявності катіонів Mg
2+
цей компонент є дуже
пом’якшений негативними потенціалами, але він є при вcix
позитивних струмах. За наявності APV пізній компонент зникає
при будь-яких струмах. Отже, пізній компонент вхідного
синаптичного струму спричинений активуванням NMDA-
рецепторів.
Канал NMDA-рецептора є “подвійно-ворітним”
(С. Hammond, 1996). Відомі лише дві умови, за яких відбувається
його активування: наявність ліганда глютамату (i, можливо,
гліцину) та деполяризація мембрани. Слід зауважити, що NMDA
канали блокуються йонами Mg
2+
при струмі близькому до
потенціалу спокою клітини. Чутливість до струму NMDA каналів,
спричинена катіонами Mg
2+
, радикально відрізняється від
чутливості до струму Na
+
i Ca
2+
каналів, що є внутрішньою
особливістю білка, коли не спостерігається зовнішньо- та
внутрішньоклітинне блокування йонів. Отже, виникає питання, чи
наявність нейротрансмітера у синаптичній щілині є достатньою,
щоб спричинити постсинаптичну реакцію NMDA.
358
Відомо також, що не-NMDA- та NMDA-рецептори
співіснують на постсинаптичній мембрані (C.J. McBain, M.L. Mayer,
1994). Так, Едмондс та ін. (В. Edmonds et al., 1995) спробували дати
відповідь, якою буде частина синаптичного струму, що формується
активуванням NMDA-pецепторів. Нейротрансмітером у більшості
синапсів ЦНС хребетних слугує глютамат. Вивчаючи вплив
ендогенного глютамату на не-NMDA- та NMDA-рецептори у разі
фіксування петч-клемп на постсинаптичних нейронах та
порівнюючи з ідентифікованими пресинаптичними нейронами,
вчені висунули таку гіпотезу.
Глютамат виділяється в синаптичну щілину, де зв’язується
з постсинаптичним NMDA-рецептором так само, як i з нe-NMDA-
рецептором. Коли мембранний потенціал наближається до
потенціалу спокою клітини, велика щілина NMDA-рецептора
блокується йонами Mg
2+
, які наявні в синаптичній щілині. Тому
виділений глютамат активує нe-NMDA-рецептори (В. Edmonds et
al., 1995).
Доведено (С. Hammond, 1996), що стимулювання
пресинаптичних нейронів гіпокампа в разі браку йoнів Mg
2+
зумовлює постсинаптичну деполяризацію, що є результатом
синаптичного вхідного струму та пов'язана з перехрещенням
NMDA та не-NMDA каналів. Якщо немає катіонів Mg
2+
та за
наявності APV, спостерігається зниження тривалості
постсинаптичної деполяризації.
Компонент, поява якого спричинена активуванням
NMDA-рецепторів, має повільнішу фазу зростання деполяризації
й триває довше, ніж компонент, опосередкований нe-NMDA-
359
рецепторами. Отже, коли NMDA-рецептори є активовані, пік
постсинаптичної деполяризації не завжди простежується, але
тривалість такої деполяризації є значно більшою.
Використання селективних антагоністів NMDA- чи не-
NMDA-рецепторів дало можливість вивчати кожен з цих
рецепторів окремо (під час синаптичних відповідей) у разі дії
ендогенних збуджуючих кислот. Відомо, що клас нe-NMDA-
pецепторів поділяється на дві групи: рецептори, які активуються
АМРА та квісквалятом i мають до них високу спорідненість
(В. Edmonds et al., 1995; С. Hammond, 1996; C.J. McBain, M.L. Mayer,
1994). Канал, активований квісквалятом (10 мкмоль/л), у
зовнішньоклітинному середовищі на культурі спинних нейронів
щура, є проникним для одновалентних катіонів: використання
квіскваляту при МП –60 мВ спричиняє одиничний вхідний струм,
який є результатом входу йонів Na
+
та виходу йонів К
+
через той
самий канал (причому вхід натрію більший, ніж калію)
(C.J. McBain, M.L. Mayer, 1994). Кінетика струмів є різною. Цей
АМРА-рецептор має незначну проникність для катіонів Са
2+
, а
його провідність лише частково залежить від струму. Тим паче,
дослідження, проведені на інших об’єктах, засвідчили, що деякі
АМРА-рецептори є проникними для йонів кальцію (С. Hammond,
1996).
Більшість каналів, які активовані квісквалятом, мають
провідність 8 пСм, яка є малочутливою до змін мембранного
потенціалу. Крім того, одиничний струм реверсує при потенціалі,
близькому до 0 мВ, якщо зовнішньо- та внутрішньоклітинні
оточуючі середовища містять подібну концентрацію
360
одновалентних катіонів. Це значить, що канал, який активується
квісквалятом є проникним для катіонів. Щоб перевірити цю
гіпотезу, потенціал реверсії струму був зафіксований у різних
внутрішньо- та зовнішньоклітинних концентраціях іонів Na
+
та К
+
.
Коли зовнішньоклітинна концентрація Na
+
зменшується від 140 до
50 ммоль/л, замінюючи йони Na
+
холіонами, потенціал реверсії
стає негативним (він зменшується від 0 до –20 мВ). Якщо йони
Cs
+
замінюють внутрішньоклітинні йони К
+
, реверсія струму не
відбувається. Ці результати дають змогу переконатися, що канал,
активований квісквалятом, є проникним для йонів Na
+
, К
+
та Cs
+
i
непроникний для холіну.
Було доведено (P. Jonas, N. Bumashev, 1995), що цей канал
має низьку проникність для двовалентних катіонів, особливо йонів
Ca
2+
. Отже, зміни, здійснені цими іонами, не впливають на
потенціал реверсії квісквалятного потоку, зафіксованого на
нейронах у конфігурації “whole-cell”. Подібним чином лише
незначні зміни фотометрично зафіксованої внутрішньоклітинної
концентрації Са
2+
можуть бути виміряні під час відповіді,
зумовленої квісквалятом при струмі –60 мВ. Автори звертають
увагу на важливість фіксування потенціалу в клітинах, оскільки
поріг активування для потенціалзалежних Са
2+
каналів, є нижчим,
ніж поріг входження йонів Ca
2+
через цi канали.
Дарлісон (M.G. Darlison, 1992) вивчав фармакологічні
властивості ГАМК- та глютаматних рецепторів на молюсках та
дрозофілі, що стало стимулом до дальших досліджень в
електрофізіології та фармакології білків іонних каналів у
безхребетних.