Scheffler Immo E. Mitochondria
Подождите немного. Документ загружается.

HUMAN EVOLUTION 421
There are additional disputes about the calibration of the clock. Is the rate
of change the same at all sites where changes have been observed? Do non-
coding regions (D - loop) change more rapidly than coding regions? Are the
changes in the third positions of the codons occurring at a faster rate? What
about the relative rates of transitions versus transversions in a DNA sequence?
The challenge was taken up by Hasegawa et al. (17) to predict a more accurate
time scale for human mtDNA, making the additional assumption that the
divergence of humans from our closest primate ancestor, the chimpanzee, was
about 4 million years ago. The emergence of modern humans out of Africa
was moved to an even more recent date: 89,000 ± 69,000 years ago. Such a
result also put the authors squarely at odds with the multiregional hypothesis
of modern human origin, which has gained adherents in recent years. Uncer-
tainties and controversies related to the mutation rate in mitochondria have
by no means been resolved to everyone ’ s satisfaction, and as a result the esti-
mates for the age of the mitochondrial Eve range from < 100,000 years to
almost a million years (see below).
After the African mitochondrial Eve was declared “ wounded but not dead
yet ” (18) in 1992, based on the ambiguity surrounding the construction of
phylogenetic trees with mtDNA sequences, supporters of the hypothesis have
assembled independent support. The controversy has at times been heated.
An attempt by a respected expert to debunk the “ myth of Eve ” (19) was fol-
lowed a few issues later by a rebuttal by another expert accusing the fi rst of
perpetuating another myth (20) . As Ayala states succinctly, the coalescence of
the mtDNA sequences from over 100 ethnically diverse individuals to one
ancestral sequence, that of the “ mitochondrial Eve, ” is a property of all gene
genealogies, but does not mean that all humans descended from a single
mother. While Wilson and his colleagues probably also did not mean to be
taken literally, the statement had an attention - catching appeal, and the popular
press made the most of it. If not a single mother, there is still the question
about the size of the pool of humans from which the present - day human
groups are descended. The current controversy therefore centers around two
or three problems. Where is the mtDNA genealogical tree rooted? Evidence
points strongly toward Africa, but it is not yet conclusive. The second question
is concerned with the existence of a bottleneck, as well as the size of the bottle-
neck of those individuals who are the ancestors of all modern humans. Finally,
there may have been multiple bottlenecks with subsequent mixing of the sur-
viving populations. The arguments about multiregional models require exten-
sive migrations and interbreeding between populations to account for a single
gene pool (19) .
Ingenious independent analyses have strengthened the out - of - Africa
hypothesis. To establish the true root of the phylogenetic tree, the true human
mtDNA ancestor, one must choose an outgroup sequence from a different
species more ancient than humans, but still closely related. The chimpanzee
mtDNA has served such a purpose; but in the control region used routinely,
the rate of change may be too fast and too extreme. The creation of multiple
422 MITOCHONDRIAL DNA SEQUENCING AND ANTHROPOLOGY
and parallel sequence changes can complicate and obscure the analysis. A
novel fi nding was exploited elegantly by Zischler et al. (21) to identify a supe-
rior outgroup for rooting the human mtDNA tree. Following the original
discovery of Fukuda et al. (22) , several investigators have confi rmed that the
human nuclear genome contains several hundred copies of sequence frag-
ments closely resembling mtDNA. Such pseudogenes have also been found in
other primates and rodents, and their distribution and sequence alterations
make them useful molecular fossils in the nucleus (23) . Zischler et al. (21)
specifi cally searched for and found a nuclear copy of the human mitochondrial
D - loop which had been transferred to the nucleus after the divergence of
humans from the chimpanzee, but before the “ growth ” of the human mito-
chondrial phylogenetic tree. The nuclear D - loop sequence became a superior
out group for rooting the tree in Africa (24) .
The mtDNA data assembled by many groups — prominently among them
the group at Emory University, as well as Stoneking and colleagues at Penn-
sylvania State University — have generally supported the following scenario
for the expansion of Homo sapiens around the globe (25) : The migration
started out of Africa about 100,000 to 200,000 years ago. It split somewhere
in the middle east into a group headed for central and northern Europe
(∼ 50,000 years ago), and another group headed toward southern and southeast
Asia. Migrations from present day China via the Indonesian Archipelago
toward New Guinea and Australia also occurred a long time ago. Somewhat
later the populations expanded north along the eastern rim of Asia, and even-
tually groups reached the Bering Strait to cross into North America. There
may have been several crossings of the Bering Strait (see below). Expansion(s)
from Alaska to the south populated in order North America, Central America,
and South America. Quite recent in human history, another wave of expansion
occurred from the islands surrounding New Guinea and North Australia into
the northern Pacifi c (Polynesia and Hawaii).
Of course, many of these deductions were not completely new. Data from
paleoanthropology and archeology had already been used to make broadly
similar deductions. Linguistic comparisons have also contributed greatly. Nev-
ertheless, mtDNA sequence data can often provide detailed and convincing
support. For example, a 9 - bp deletion in the small intergenic region between
COXII and tRNA
Lys
is believed to have arisen in Asia about 50,000 years ago.
Redd et al. (26) have traced a specifi c set of control region mutations ( “ the
Polynesian motif ” ) associated with the 9 - bp deletion to a Taiwan origin (about
6000 – 8000 years ago), and from there across the Philippines, Indonesia, along
the coast of New Guinea, and fi nally into Polynesia (26) . These molecular data
are in excellent agreement with the postulate of a southeast Asian origin of
Polynesians based on archeological and linguistic evidence.
The contention about rooting the human phylogenetic tree may have
approached a consensus, or at least the African root has acquired the support
of the majority. It is yet another issue to date the period of the existence of
the mitochondrial Eve. Determining the age of Eve is important, because an
HUMAN EVOLUTION 423
accurate estimate of the time span to the present serves in arguments about
other theories, specifi cally the multiple origins theory. Since the fossil record
reveals the existence of Homo erectus as long as at least a million years ago,
and such evidence is found in Europe, the Middle East, and Asia, one is forced
to conclude from the genetic evidence that the modern humans (our ances-
tors) displaced all of these other human populations completely without any
detectable genetic exchange. If the lowest estimate for the age of the mito-
chondrial Eve is accepted ( < 100,000 years), this scenario would be quite aston-
ishing. On the other hand, if the mitochondrial Eve lived more than 500,000
years ago, a new set of questions and theories can be entertained about mul-
tiple origins, shifts in the composition of the human gene pool, and selective
pressures that might have contributed to the unusual evolutionary pattern. A
thoughtful reexamination of existing data sets by Wills (27) comes to the con-
clusion that Eve may be between 436,000 and 806,000 years old, an estimate
that is also very dependent on the estimate of the time of the split between
the human and chimpanzee lineages.
In the past decade there has been an avalanche of sequence data for
mtDNA, the Y chromosome, and autosomes from a wide range of human
donors. An up - to - date analysis of the conclusions has appeared from Garrigan
and Hammer (28) . These authors clearly distinguish two epochs in the evolu-
tion of H. sapiens . The fi rst, starting about 2 million years ago, leads to the
emergence of the “ anatomically modern human. ” The second epoch covers
the geographic distribution of the modern humans after their emergence from
Africa. Sequence analyses are used to construct gene trees, and these in turn
can be interpreted to estimate the “ time to the most recent ancestor ” (TMRCA),
with certain assumptions about the molecular clock for accumulating nucleo-
tide changes. The mtDNA tree from the complete sequences of a global sample
of humans yields a TMRCA of ∼ 170,000 years. From the tree based on the
nonrecombining regions of the Y chromosome the estimate for the TMRCA
is roughly half this value, and various explanations have been offered (see
reference 28 ). As more and more sequence data from loci on the X chromo-
some or autosomes become available, the TMRCA can be pushed back to
500,000 – 800,000 years and thus include H. neanderthalensis, H. heidelbergensis ,
and other now extinct lineages of H. sapiens . Some of these results may con-
tradict the “ single origin model ” derived from mtDNA and Y chromosome
trees, and the multiregional evolution model may gain favor in the future. The
interested reader is directed to this highly technical paper for a consideration
of alternate models. Some support for the claim that modern humans from
Africa actually had “ close physical contact ” (does that mean sex?) with archaic
humans ( H. erectus ) comes from an unexpected direction. A genetic analysis
of lice revealed that there are two subspecies, one thought to have lived on H.
erectus and the other on H. sapiens . After a relatively long period of isolation
of both the lice and the humans, physical contact between these two groups of
humans before H. erectus became extinct must have taken place to explain the
present - day distribution of these louse subspecies in the world (29, 30) .
424 MITOCHONDRIAL DNA SEQUENCING AND ANTHROPOLOGY
Mitochondrial DNA sequence analysis has been perfected to the level
where analyses have become possible with samples from a 4000 - year - old
mummy of an Egyptian priest and from a 7000 - year - old human brain from a
Florida peat bog; and if the bones are ever released for analysis before they
are re - buried, there are paleobiologists waiting to obtain samples from a skel-
eton found on the northwest Washington coast thought to be 9000 years old.
One of the most impressive achievements in the study of human evolution by
the study of human DNA sequences was reported by Krings et al. in 1997 (31) .
The group in Munich, with independent confi rmation by a group in Pennsyl-
vania, reported mtDNA sequences from a Neanderthal person. The Neander-
thals were a race of humans roaming through Europe and Western Asia from
about 300,000 to 30,000 years ago. Considerable discussion had centered
around the question whether Homo neanderthalensis was a distinct and sepa-
rate species from Homo sapiens . Their range overlapped in Europe during the
more recent millenia, and the possibility of a mixing of the gene pools by
interbreeding had not been convincingly excluded. In the landmark paper
published by the Paabo and Stoneking groups the defi nite conclusion was
reached that the Neanderthals went extinct, leaving no trace of their mtDNA
in modern humans. Concentrating on the D - loop region, sequence compari-
sons and phylogenetic analyses were made, including a large number of rep-
resentatives of modern humans as well as the chimpanzee. The divergence of
the Neanderthal from modern humans was estimated from the mtDNA analy-
ses to have occurred about 500,000 years ago, in satisfactory agreement with
other estimates based on the archeological record.
Krings et al. (31) is a landmark for its conclusions, but it probably also set
the standard for future studies of ancient DNA. The DNA was retrieved from
a fossilized bone of an individual living between 100,000 and 30,000 years ago.
As discussed in more detail in this paper and elsewhere, measurements of
amino acid racemization can give reliable estimates about preservation and
the feasibility of obtaining suffi cient DNA from a fossil specimen, and the
Neanderthal bone may be near the limit of what constitutes a useful source.
The quantitative considerations also indicate that only mtDNA sequences can
be expected to be recovered for PCR - based amplifi cation, because it is several
orders more abundant than nuclear DNA. In a commentary on this paper in
the same issue of Cell , T. Lindahl discusses the “ fi asco of DNA from insects in
amber, ” with reference to studies suggesting that amber is a very poor material
for the preservation of DNA (but not of the insect exoskeleton) (32) . Thus,
claims of DNA amplifi cation from million - year - old insects in amber may con-
tinue to stimulate Hollywood, but they cannot stand up to rigorous scrutiny.
Because such skepticism is clearly indicated, the Neanderthal mtDNA paper
went to extraordinary lengths to include controls and to obtain repeatable
results, even when conducted in a distant, second laboratory. Since this paper
appeared, more partial sequences from Neanderthal mtDNA have appeared,
confi rming the original conclusions in favor of the “ rapid replacement ” model
HUMAN EVOLUTION 425
for the origin of modern humans and the absence of any interbreeding between
Neanderthals and modern humans (33, 34) .
Another, more limited application of mtDNA sequence analysis to human
evolution and history is also instructive, but ultimately baffl ing to the outsider
because of still unresolved controversies. Anthropologists have been curious
for some time about the following questions: When were the Americas popu-
lated by humans, and where did these people originate? Before the advent of
molecular genetic analyses, several time - honored approaches had suggested
answers, which cannot be discussed here in detail. The more conventional
archeological studies use the various artifacts (pottery, tools, weapons) to trace
lineages based on designs, patterns, materials used, and so on. A more recent
development is the application of the tools of linguistics to the comparative
analysis of the languages spoken by the various native Americans (North and
South America). Such an analysis had suggested three distinct linguistic divi-
sions: Amerind (spoken by American Indians in North, Central, and South
America), Eskimo - Aleut (spoken by the Inuit (Eskimo) and Aleutian Island-
ers), and Na - Dene (spoken by the by people of the Northeast Coast of Canada
and the United States as well as the Navajo and Apache, believed to have
migrated south to their present location around 1000 bc ). An interpretation
was that there were three distinct waves of immigration across the Bering
Straits, in agreement with other archeological data, such as distinctive features
of molar shapes. Mitochondrial DNA data assembled by Wallace, Torrioni, and
colleagues (reviewed in reference 25 ) not only confi rmed the Asian origin of
the native Americans, but appeared to lend strong support to the three waves
of migration hypothesized by the linguists. Four haplogroups were found based
on extensive restriction mapping, and the relative abundance of these groups
could be interpreted in terms of three distinct migrations separated in time by
thousands of years.
Somewhat ironically, an ex - student of Wallace has collected additional data
that challenge the interpretation of his mentor (35) . The new fi ndings of all
four haplotypes among the three linguistic groups (mtDNA samples from 1300
American Indians) appear to fi t better to a scenario in which a single wave of
immigration brought women with all four haplotypes across the Bering Strait.
Subsequent migrations, along with the possibility of severe temporary reduc-
tions in one or more local populations due to environmental infl uences, may
have caused the loss of particular haplotypes (another bottleneck), resulting
in the current distributions.
The discussion continues. It is not the intent here to present the defi nitive
conclusion, if there is one, but to make the reader appreciate the wealth and
diversity of ideas derived from this research.
There are now well in excess of 2100 mitochondrial DNA sequences in the
Human Mitochondrial Genome Database (mtDB) ( http://www.genpat.uu.se/
mtDB ), including more than 1544 complete sequences (36) . The database also
lists more than 3000 sequence polymorphisms from human population groups
426 MITOCHONDRIAL DNA SEQUENCING AND ANTHROPOLOGY
all over the globe. This is a treasure trove for human population geneticists
and medical researchers interested in correlations between mtDNA haplo-
types and phenotypes. The “ harvesting of the fruit from the human mtDNA
tree ” is in full swing (37) , with new disciplines, “ archeogenetics, ” “ phylogeo-
graphy, ” and so on, being created. Another exhaustive database is discussed
by Attimonelli et al. ( http://www.hmdb.uniba.it ) (38) .
Regional variation in mtDNA haplotypes has generally been attributed to
genetic drift and migration. The question has been raised as to whether some
of these sequence variations in coding regions might have functional signifi -
cance (39) and whether a selective advantage could arise for a population in
a specifi c environment (40) . The authors conclude that some mtDNA lineages
found in European, Asian, Siberian, and Native North Americans might refl ect
adaptations to a cold climate. Such an advantage could be achieved by a slight
decrease in the coupling of electron transport and ATP synthesis in favor of
thermogenesis. However, at this time such a conclusion must remain quite
speculative.
8.3 PRIMATE EVOLUTION
Phylogenetic analysis of mitochondrial DNA sequences can of course be
pushed back in time to include other species. No signifi cant surprises about
evolutionary history may be expected when comparing different vertebrates,
or even different mammals. However, for calibrating molecular clocks and for
establishing more precisely the time of divergence of the nonhuman apes, a
comparison of the mtDNA sequences of humans, chimpanzee, pigmy chim-
panzee, gorilla, and orangutan has been very informative. Horai et al. (41)
found these genomes to be highly homologous, with a synonymous substitu-
tion rate of 3.89 × 10
− 8
/site per year, if one assumes that the orangutan and
the African apes diverged 13 million years ago. In the D - loop region this rate
was almost twice as high. The analysis also included three human mtDNA
sequences from Africans, Europeans, and Japanese to shed light on the origin
on modern humans. From the estimated substitution rates the age of the last
common ancestor of modern humans was calculated to be 143,000 ± 18,000
years, and the hypothesis of the African origin of Homo sapiens sapiens
received additional support.
The most detailed examination of the noncoding control region (human
nucleotide positions 16053 – 16465) of the nine taxa of African or African -
derived hominoids (humans, chipanzees, bonobos and gorillas) has been
undertaken by Gagneux et al. (42) , who examined 1158 unique haplotypes
including 811 humans from around the world. A phylogenetic tree was con-
structed which also included orangutans and the newly established sequence
from Homo neanderthalensis (Figure 8.1 ). For details on the computer analysis
and related analytical methods, the reader is referred to the original paper.
The fi nal tree shown was also subjected to various degrees of topiary pruning
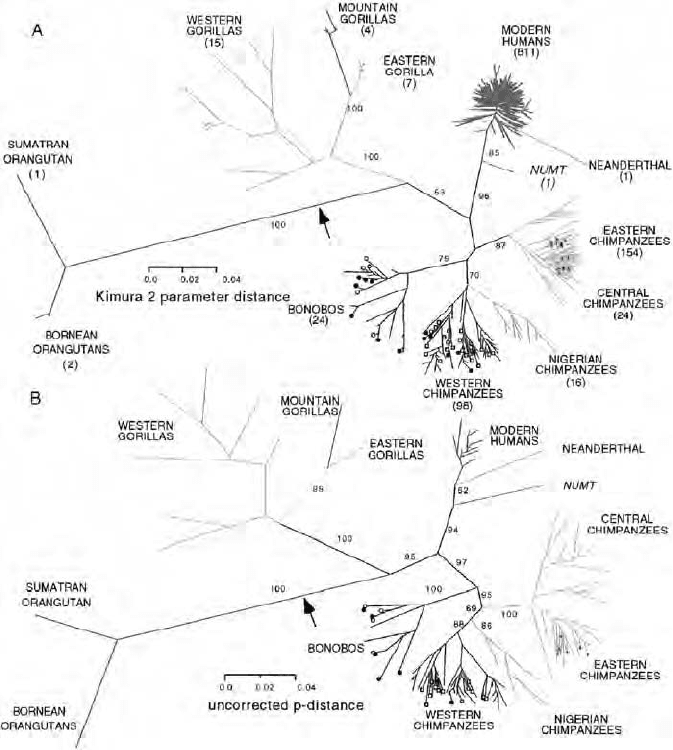
Figure 8.1 Primate evolution based on the analysis of mtDNA. An unrooted phylo-
gram of the neighbor - joining tree of 1158 different mitochondrial control region
sequences was constructed (A) and then subjected to topiary pruning (B). See Gagneux
et al. (42) for details. [The fi gure was generously provided by Drs. Gagneux and Wills
prior to publication (with permission).]
(16) , but only one intermediate level is shown here. The tree illustrates in
dramatic fashion some previous conclusions, and it adds some novel insights.
It is clearly supportive of the closer relationship between humans and chim-
panzees relative to the other great apes. Most remarkably, it reveals deep
branches between chimpanzees in various African locations, corresponding to
the existence of several subspecies, and a previously unrecognized subclade of
PRIMATE EVOLUTION 427
428 MITOCHONDRIAL DNA SEQUENCING AND ANTHROPOLOGY
chimpanzees in Nigeria and northern Cameroon. There is signifi cantly more
variation between the major chimpanzee clades than between the entire
human population. The appearance of the tree and specifi cally the relatively
shallow human branches are consistent with the idea of a demographic bottle-
neck in human evolution — in other words, consistent with the idea that the
present human lineage arose from a small effective population of relatively
recent origin. A mitochondrial DNA fragment that entered the nuclear genome
was also included: It originated after the divergence of humans from the
great apes, and even after the divergence of present humans from the
Neanderthals.
An interesting corollary to these studies was published by Kenyon and
Moraes (43) . These authors used human ρ
0
cells (mtDNA - less) in fusions with
enucleated cells from the hominoid apes chimpanzee, pigmy chimpanzee,
gorilla, and orangutan to produce “ xenomitochondrial cybrids. ” Cybrids with
chimpanzee or gorilla mtDNA and a human nucleus could respire and carry
out oxidative phosphorylation. In contrast, the orangutan mtDNA and the
human nuclear genome were no longer compatible. The results support the
notion that the nuclear and mitochondrial genomes have to co - evolve at a
signifi cant number of loci in order to maintain the critical interactions between
OXPHOS proteins encoded by the nucleus and proteins encoded by the mito-
chondria. Thus, the more closely related ape mt genomes could be functionally
expressed to interact with human nuclear gene products, but the orangutan
mt genome had evolved far enough since the distant separation of the lineages
to yield mitochondrial proteins which presumably could no longer interact
with human proteins in the inner mitochondrial membrane. It will now be a
challenge to distinguish simple polymorphisms with no structural – biochemical
consequences from those nucleotide changes (and the resulting amino acid
substitutions) which contribute to the defective interactions between nuclear
and mitochondrial gene products. Unfortunately, the use of recombinant
(human – primate) mtDNA molecules is technically still impossible. On the
other hand, the available crystal structures for complexes III and IV may guide
the search for protein – protein interaction involving nuclear - coded and
mitochondrial peptides and may perhaps reveal critical contact sites.
8.4 HUMAN Y CHROMOSOME VARIATION
A crucial, inherent advantage of the mitochondrial genome in the analyses
described above is (a) the absence of recombination, facilitating the construc-
tion of phylogenies, and (b) the estimation of the time intervals elapsed
between the different branch points, assuming that the molecular clock is
running at a constant rate. Another chromosome that has long been recog-
nized not to participate in recombination is the Y chromosome. The statement
has to be qualifi ed, since a portion at the tip of the short arm, the pseudoau-
tosomal region, does participate in chiasma formation and recombination. The
male - specifi c portion of the Y chromosome, however, offers an alternative and
complementary system for the study of human evolution and migratory path-
ways, and data on polymorphisms on this chromosome have been collected
and interpreted extensively. It is self - evident that the Y chromosome is inher-
ited along the paternal lineage, since it is the male - specifi c sex chromosome
containing the critical gene (SRY) encoding the testis determining factor (44) .
By comparison with mtDNA, the Y chromosome is huge, comprising approxi-
mately 60,000 kb (60 Mb) of DNA. The great bulk of Y chromosome DNA,
especially the long arm (Yq), is made up of constitutive heterochromatin
consisting of various families of repetitive DNA with no obvious genetic
information. It has taken some time to defi ne unique loci and to establish
polymorphisms at these loci in human populations (45 – 50) .
A detailed discussion of human Y chromosome polymorphisms would go
beyond the scope of this monograph. On the other hand, it is worthwhile to
consider how far the deductions from the analysis of mtDNA have been con-
fi rmed by similar studies with Y chromosome sequences, and what kinds of
unresolved discrepancies exist. Independent confi rmations of the reconstruc-
tion of human evolution are of course a source of intellectual satisfaction, and
they strengthen the conclusions drawn. Discrepancies, on the other hand, raise
issues about methodology, interpretation, and limitations of computer analy-
ses, but in the long run they can lead to new insights and progress.
A comprehensive and recent description of the geographic distribution of
human Y chromosome variation can be found in the publication by Hammer
et al. (50) , which includes data from 1500 males from 60 population groups,
subdivided into 15 major groups based on geographic, linguistic, and historical
information. In agreement with other genetic analyses, and specifi cally the
mtDNA data, the Y - chromosome haplotype distributions show that sub -
Saharan African populations are distinct from the rest of the world, and they
exhibit the highest within - group diversity. Globally, the variants found outside
of Africa represent subsets of the haplotypes found within Africa. Subsequent
results from a more extensive sequencing of unique regions of the human Y
chromosome also supported the idea that “ Adam was an African ” (51 – 53) .
Hammer et al. were careful to point out that while such a within - group and
among - group pattern also supports the hypothesis of the origin of modern
humans from an older African population — with migrations of subgroups
leading to the population of distal sites, along with more limited genetic diver-
sity elsewhere resulting from bottlenecks — it may nevertheless be an overly
simplistic view. In the absence of additional information, the genetic data are
equally consistent with multiple short - range and long - range migrations, genetic
drift due to founder effects, and small population sizes (near extinction).
Africa may well have been the origin of more than one migration originating
from different parts of Africa. Notably, the Y chromosome data also are con-
sistent with the relatively recent ( ∼ 100,000 years) origin of our common ances-
tors, but may be a consequence of a small effective population size throughout
the Late Middle Pleistocene, rather than a true defi nition of the period of
HUMAN Y CHROMOSOME VARIATION 429
430 MITOCHONDRIAL DNA SEQUENCING AND ANTHROPOLOGY
origin of Homo sapiens . For additional theoretical as well as methodological
considerations the reader is referred to several publications from Hammer
and colleagues (28, 50) , as well as the numerous additional publications cited
in these papers.
8.5 FORENSIC APPLICATIONS
Under other monarchies the male line takes precedence of the female in tracing
genealogies, but here the opposite is the case — the female line takes the prece-
dence. Their reason for this is exceedingly simple, and I recommend it to the
aristocracy of Europe: They say it is easy to know who a man ’ s mother was, but,
etc., etc.
— Mark Twain, “ Letters from the Sandwich Islands, ” describing the
Hawaiian Monarchy
Mitochondrial DNA sequences also mutate rapidly enough to create distinc-
tions (polymorphisms) between members of the same ethnic group — that is,
close “ relatives ” on an evolutionary scale, though not relatives in the legal
sense. Applications in forensic investigations have been made, and two such
applications will be used as examples to illustrate the methodology and the
potential limitations.
Attention is focused on the mtDNA control region (D - loop). In 1983 in a
relatively small sample of Caucasians and Africans it was fi rst found that there
was ∼ 1.7% nucleotide diversity in ∼ 900 basepairs, with most of the differences
clustered in two hypervariable regions (54) . The 347 - basepair hypervariable
region (within nucleotides 16023 – 16388) was later analyzed by Orrego and in
10 additional unrelated Caucasian individuals of European ancestry. In the
combined data set from Greenberg et al. (54) , Anderson et al. (55) , and Orrego
and King (55a) , 32 sites out of 347 sites compared were found to be variable,
and pairwise comparisons showed differences ranging from 1 to 13 nucleotides
(average 5.9). A formal statistical analysis revealed that the observed and
expected number of sequence differences between pairs of individuals closely
approximated a Poisson distribution, which allows one to calculate that the
probability of two unrelated Caucasian individuals having an identical
sequence of 347 basepairs is 1 in 370 (see reference 56 ) for a discussion of
these results and related issues).
With oligonucleotide primers corresponding to the conserved fl anking
sequences (tRNA
Pro
and tRNA
Phe
), it is a routine matter to amplify the D - loop
region from very small tissue samples. As described by King (56) , a second
amplifi cation with nested but unequal amounts of primers can yield a single -
strand product ready for DNA sequencing. Optimization of conditions to
reduce artifacts from polymerase errors is part of the standard methodology,
and mosaics created with unintended target sequences can be detected and
eliminated because the normal target is so well known. A successful test analysis