Scheffler Immo E. Mitochondria
Подождите немного. Документ загружается.

the phenomenon for the present discussion has been the well - documented
involvement of mitochondria or mitochondrial functions.
It has become useful to subdivide the process of apoptosis into several
phases or stages. The fi nal phase encompasses a number of characteristic
changes such as cell shrinkage, chromatin degradation and nuclear fragmenta-
tion, and loss of plasma membrane integrity. It appears that these changes are
essential for rapid recognition of the dying cell by its neighbors, which phago-
cytose the remnants and effi ciently eliminate any debris that would otherwise
cause an infl ammatory response by the immune system. As a consequence,
“ dead ” cells from apoptosis are not readily seen in vivo in intact tissue, and
the description of this terminal phase rests largely on observations in tissue
culture.
The fi rst phase includes the description of the various stimuli or signals
that induce apoptosis in different cells. The stimulus may be an external one,
received by a cell surface receptor, or an internal one, as the result of the action
of a drug, toxin, or radiation damage. Of course, drugs, toxins, and radiation
are also ultimately external infl uences, but the difference between these two
mechanisms becomes apparent during the following phase: the signal trans-
duction, which leads to changes in metabolic state, induction of various enzymes
(proteases, nucleases), and the morphological changes preceding cell death.
Needless to say, these two signaling pathways may converge at some level, but
at the same time the above description should not give the impression of a
simple, universal and invariable pathway operating in all cells under all condi-
tions. Deprivation of growth factors (e.g., interleukin - 3 for hematopoietic
progenitor cells) induces apoptosis; that is, unoccupied receptors can stimulate
apoptosis. Failure to stimulate germinal center B cells by antigens causes them
to die. Transforming growth factor β and tumor necrosis factor can trigger
apoptosis in various cell types. Many more examples could be given (247, 250) .
It should be clear that these signal transduction pathways include most, if not
all, of the following known reactions, intermediates, and interactions: tyrosine
kinases, phosphatases, steroid receptors, inositol phosphates, transcriptional
activators and factors, and so on.
There is usually a signifi cant time interval (several hours) between the
delivery of the fi rst stimulus and the observation of morphological changes in
the targeted cells. It is also during this interval that thresholds or checkpoints
are reached beyond which the process becomes irreversible. Many details of
events occurring during this effector phase remain elusive and are the subject
of a very active research.
As mentioned above, the nematode, C. elegans , continues to be a useful
model system, and it was in this model system that the fi rst genes specifi cally
associated with apoptosis were isolated (ced1, ced - 2, ced - 3, . . . , ced - 10, etc.).
Many of these genes were subsequently shown to be required for the disposal
of the dead cell(s), and hence their detailed description will be omitted here.
The Ced - 3 gene, however, was identifi ed to encode a cysteine protease, and
many different homologues have since been found in mammalian cells. The
MITOCHONDRIA AND APOPTOSIS 391
392 MITOCHONDRIAL MUTATIONS AND DISEASE
fi rst was interleukin 1 β converting enzyme (ICE), and hence this group of
aspartate - specifi c cysteine proteases was initially referred to as ICE x , where
x is a distinguishing subscript. More recently, this family of proteases has been
referred to as caspases. In a simplifi ed view, apoptosis is accompanied by
extensive proteolytic cleavage, and there is in fact a cascade of such cleavages,
since these ICE enzymes exist as inactive precursors (pro - caspase) that must
be cleaved after a specifi c aspartate residue before they can be assembled into
heterotetramers and active proteases. The other targets of these proteases are
still largely unknown, although it was intriguing that one of these proteases
(CPP32) can cleave the poly - (ADP ribose) polymerase (PARP), an enzyme
involved in DNA repair. However, PARP knockout mice develop normally.
The ICE proteases are constitutively expressed in many cells, and their activa-
tion is a post - translational event, although some additional transcriptional
activation following the death stimulus may also occur. It is apparent, there-
fore, that there must be an upstream signal for post - translational activation
which remains to be more clearly defi ned in mammalian cells. Certain broad -
range protease inhibitors can prevent apoptosis in response to common stimuli,
but other highly specifi c and restricted inhibitors may interfere with the signal-
ing pathway initiated from only a limited number or a unique inducer. Apop-
tosis induction may have “ common ” as well as “ private ” pathways.
The ced - 9 mutation in nematodes led to the characterization of the corre-
sponding nematode gene and the potential functional identifi cation of the
human homologue Bcl - 2. The Bcl - 2 protein was localized in the outer mito-
chondrial membrane (although see below), and this fi nding was one of the fi rst
links to mitochondria. The mammalian Bcl - 2 gene was initially implicated in
tumorigenesis because it was found to be the location of a breakpoint (18q21)
in a reciprocal translocation found persistently in human B - cell tumors (251 –
254) . Bcl - 2 protein became overexpressed under the infl uence of an immuno-
globin promoter. A whole family of bcl - 2 - like proteins (Bcl - 2, Bcl - x
L
, Mcl - 1,
Bfl - 1, A1, . . . ) were subsequently defi ned and characterized as inhibitors of
apoptosis by transfection studies and overexpression in various cells (254) .
Other family members have also been localized in the outer mitochondrial
membrane (255) . In addition, factors promoting apoptosis have also been
found: Bax, Bcl - x
S
, Bad, Bak, Bik, . . . and so on (see below).
Before discussing Bcl - 2 and mitochondria in more detail, some other
approaches related to the study of apoptosis and mitochondria must be intro-
duced. Active, healthy mitochondria exhibit a transmembrane potential ( ΔΨ
m
)
across the inner mitochondrial membrane which is of fundamental importance
in the discussion of the mechanism of oxidative phosphorylation (see Chapter
5 ). A pH and electrochemical gradient across the inner membrane is set up as
the result of electron transport through the electron transport chain, with
excess positive charge on the outside — that is, in the intermembrane space.
The discovery was made some years ago that certain lipophilic dyes such as
rhodamine 123, 3,3 ′ - dihexyloxacarbocyanine iodide (DiOC6(3)), 5,5 ′ ,6,6 ′ -
tetrachloro - 1,1 ′ ,3,3 ′ - tetraethylbenzimidazolcarbocyanine iodide (JC - 1), and
chloromethyl - X - rosamine (CMXRos) (see Chapter 5 for a more detailed
description) can accumulate in the matrix. Under controlled conditions the
fl uorescence intensity can be correlated with the magnitude of the membrane
potential ( ΔΨ
m
), and these dyes have therefore been used as convenient and
powerful indicators of mitochondrial ΔΨ
m
. When such dyes were applied to
cells undergoing apoptosis, a fall in ΔΨ
m
was observed as one of the relatively
early events in many different cell types, prior to DNA fragmentation. In fact,
the collapse of the transmembrane potential was correlated in time with the
point of no return — that is, the point at which apoptosis could no longer be
reversed by withdrawal of the stimulus (256) . The breakdown of the ΔΨ
m
can
be shown to involve the so - called mitochondrial “ permeability transition ”
(PT) (257) . Briefl y, it is believed to be the result of the opening of a PT pore,
which can be measured electrophysiologically by the opening of a “ megachan-
nel, ” large enough to allow solutes of up to 1500 Da to pass through along a
concentration gradient. This PT pore is hypothesized to be regulatory with
diverse functions such as voltage sensing, thiol sensing, redox sensing, pH
sensing, sensing of adenine nucleotide levels, and regulation of calcium homeo-
stasis. Normally, only a brief opening or transient assembly of a small number
of such pores may be required, but the massive opening of many such pores
will collapse the ΔΨ
m
, stop oxidative phosphorylation, arrest the import of
proteins into mitochondria, and induce leakage of cytochrome c out into the
cytosol (see below). Thus, a long - term, massive opening of the PT pores can
be expected to do permanent damage to mitochondria, and the PT was pos-
tulated to be a central, rate - limiting event in the apoptotic cascade (255 – 257) .
In simple terms, it made sense to think that turning off the power supply was
a logical fi rst step in programmed cell death. A study of respiration defi cient
mammalian cell mutants has shown that such cells die within a day when they
are switched from a high glucose medium to medium containing galactose. In
the absence of oxidative phosphorylation, cells are sustained by a high rate of
glycolysis (30) which is not possible with galactose (see Section 1.2 ).
On the other hand, several experiments in cell free systems suggested a
more complex picture. Isolated nuclei can be induced to undergo apoptosis
when exposed to cytoplasmic extracts from cells in the process of apoptosis.
Controversy existed about whether such extracts can be free of mitochondria
or whether enrichment with mitochondria accelerates the process. Subse-
quently, the presence of “ damaged ” mitochondria was shown to be required,
and it was hypothesized that such mitochondria release an apoptogenic protein
(apoptosis inducing factor, AIF). This protein has been characterized as an
∼ 50 - kDa intermembrane protein, with protease or protease activating activity
but no nuclease activity, which can induce apoptosis in isolated nuclei more
effi ciently than ceramide, another mitochondrial constituent apparently
released by such organelles (255) .
A major breakthrough from in vitro studies of characteristic events associ-
ated with apoptosis (DNA fragmentation in isolated nuclei, activation of cas-
pases) was the discovery of the crucial role of cytochrome c, initially based on
MITOCHONDRIA AND APOPTOSIS 393
394 MITOCHONDRIAL MUTATIONS AND DISEASE
the fractionation of the cytoplasm and further demonstrated by immuno -
depletion and reconstitution experiments (258, 259) . These and subsequent
studies have suggested that the cytochrome c is involved in triggering the
cleavage and activation of DEVD - specifi c caspases. In short, cytochrome c is
now known to interact with Apaf - 1, leading to the formation of the multimeric
“ apoptosome ” in the presence of ATP or dATP (260, 261) . The apoptosome
activates the initiator caspase, caspase 9, which subsequently cleaves and acti-
vates the effector caspases 3 and 7. Many more studies have since defi ned
cytochrome c - dependent activation of apoptosis and cytochrome c - indepen-
dent pathways, the latter requiring proteins such as AIF and endonuclease G.
These proteins and others (Smac, Omi, adenylate kinase - 2) are found in the
intermembrane space of mitochondria.
A central question that was raised immediately following these observa-
tions was, How do these proteins get released into the cytosol? Much potential
confusion was generated by attempts to link the mechanism of cytochrome c
release to the breakdown of the ΔΨ and the permeability transition (PT). The
release of cytochrome c requires the permeabilization of the outer membrane,
while the PT occurs in the inner membrane. A very clear formulation of the
problem and a partial resolution of this problem can be found in the authorita-
tive recent review by Green and Kroemer (262) . The authors clearly distin-
guish between mitochondrial outer - membrane permeabilization (MOMP) and
the PT causing a breakdown of ΔΨ . Which one comes fi rst? It depends.
In one scenario the PT is associated with an equilibration/fl ux of ions and
water, causing the mitochondria to swell and leading eventually to the rupture
of the outer membrane. The PT - associated MOMP could be prevented by
specifi c inhibitors such as bongkrekic acid and cyclosporin A, which target the
adenine nucleotide transporter (ANT) and the protein cyclophilin D (CyP - D,
a peptidyl - prolyl - cis - trans isomerase), respectively. Thus, ANT and cyclophilin
D became postulated constituents of the PT pore, although this pore has has
not been isolated and well - hdefi ned even today. In current thinking the PT
pore is opened by Ca
+2
overloads, and prolonged opening causes cell death by
necrosis (263) . CyP - D - knockout mice develop normally and show no protec-
tion against a range of apoptotic stimuli, suggesting that the PT pore does not
play a role in most forms of apoptosis (263) . There is also recent evidence for
a PT pore forming in the absence of ANT, and a participation of the porin
(VDAC) channel in the outer membrane has been considered (262) .
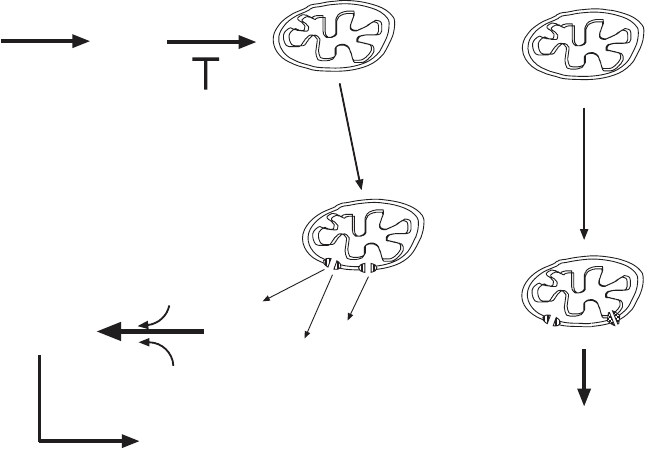
The more dominant mechanism for MOMP nowadays excludes the PT and
events at the inner membrane, and a breakdown of ΔΨ probably is a later
event following cytochrome c release and caspase activation (Figure 7.2 ). In
this mechanism the anti - apoptotic Bcl - 2 proteins (Bcl - 2, Bcl - x
L
, Bcl - W. . . . ) and
the pro - apoptotic Bax family members (Bax, Bak, Bad, Bid . . . ) play a very
prominent role. Two problems must be addressed: (1) What constitutes the
channel in the outer membrane through which proteins of considerable size
( > 50 kDa) can escape from the IMS to the cytosol? (2) What mechanisms
induce the formation of such channels that are obviously not present in normal
mitochondria? The literature on these questions is enormous, and it is a chal-
lenge to review for the uninitiated. Controversies and/or uncertainties still
abound. The interested reader should start with some recent reviews (260, 261)
and plunge into the literature. Based on in vitro studies with isolated outer
membranes or vesicles made from mitochondrial lipids, it is now believed that
oligomeric assemblies of Bax or Bak, in combination with active Bid (tBid),
form the channel (264, 265) . The possibility of a participation of porin (VDAC)
has also been considered. However, before the monomeric, cytosolic Bax or
Bad can insert into the outer membrane and form multimers, it must be acti-
vated by interaction with activated Bid (tBid, formed by caspase - 8 - mediated
cleavage of Bid) and by other mechanisms such as phosphorylation and protein
modifi cations by reactions with ROS (266) . The anti - apoptotic factors interact
with Bax or Bad to prevent activation, membrane insertion, and aggregation
Figure 7.2 (A) Schematic view of signaling in apoptosis. In this view the mitochon-
drial permeability transition takes a central position on which multiple early pathways
converge. (B) (Left) In this view the outer membrane permeabilization occurs before
the collapse of the membrane potential (see reference 262 for a recent review). (Right)
A less - defi ned mechanism leading to OMP and a permeability transition involving the
inner membrane.
Mitochondrial
permeability
transition
Multiple inducers of apoptosis
Collapse of
ΔΨ
Activation of
caspase
cascade
Chromatin condensation
DNA fragmentation
Nuclear fragmentation
Selective proteolysis
Cell shrinkage
A
MITOCHONDRIA AND APOPTOSIS 395
396 MITOCHONDRIAL MUTATIONS AND DISEASE
into channels. Bax has a C - terminal membrane - spanning region that is folded
into a hydrophobic groove in the monomer. Upon activation, this C - terminual
region becomes exposed for insertion into the outer - membrane. A truncation
of the Bcl - 2 protein deleting its C - terminal transmembrane (anchor) sequence
makes the protein incapable of suppressing apoptosis induced by several
agents. Furthermore, this anchor sequence can be replaced by the correspond-
ing anchor sequence from the yeast outer - membrane protein Mas70 (now
Tom . . . ), and Bcl - 2 targeting and function are restored. An important experi-
ment has shown that Bcl - 2 can block apoptosis even in cells that lack mito-
chondrial DNA and are unable to carry out oxidative phosphorylation (267) .
This is not to say, however, that they lack a membrane potential, since such
cells and mutant cells completely defective in mitochondrial protein synthesis
(31, 33) are perfectly capable of mitochondrial protein import.
High - resolution structures for several of these proteins are available (see
reference 261 for a summary). A similarity to the pore - forming domains of
diphtheria toxin and the bacterial colicins became apparent, supporting the
conclusions from the studies performed with liposomes. The anti - apoptotic
members have four conserved BH domains, named after the defi ning region
in Bcl - 2 — that is, Bcl - 2 homology domains (BH1, BH2, BH3, BH4); Bax/Bad
family members lack the BH4 domain, and Bid is a BH3 - only protein. The
c
s
o
c
c
c
c
c
c
c
c
c
c
o
o
o
o
o
s
s
s
s
s
s
s
Bax/Bak-mediated
outer membrane
permeabilization
(OMP)
Omi
Smac
Cyt c
APOPTOSOME
APAF-1
procaspase-9
active executioner caspases
Apoptosis-inducing
signals
BH3-only proteins
Bax
Bak
BH-123 proteins
anti-apoptotic
Bcl-2 proteins
c
s
o
c
c
c
c
c
c
c
c
c
c
o
o
o
o
o
s
s
s
s
s
s
s
mitochondrial
involvement
in necrosis
?
B
Figure 7.2 (Continued)
relative ratios and the interaction of these proteins determine the susceptibil-
ity to MOMP. Many studies with transfected cells and transgenic mice support
such a view. Bid (and other BH3 - only members) serve as “ upstream sentinels ”
ready to receive and interpret death and survival signals (260) .
The examination of mice in which either the Bcl - 2, the Bcl - x, or the Bax
genes were knocked out has yielded a complex picture. Bcl - 2 – / – mice are born
as viable pups but die at a few weeks of age. Bcl - x – / – mice die at ∼ day 13 of
embryonic development, due to massive cell death in the central nervous
system among other tissues. Bax – / – mice appear healthy, with lymphoid
hyperplasia and sterility of males due to lack of sperm cell production. Overall,
such studies support a lineage - specifi c role for these genes, but details remain
to be clarifi ed, and for the moment no signifi cant relationship to mitochondrial
function or activity can be seen from such studies in intact mice. It may have
come as a surprise that live, normal - looking mice were born.
A new development that remains to be fully explored promises to shed new
light on the mechanism of IMS protein release in the process of apoptosis. Up
till now, Bax and Bak proteins were thought to be involved exclusively in
apoptosis, by relocating from the cytosol and from a position on the ER to
induce MOMP. A poineering new study by Karbowski et al. (268) has impli-
cated these proteins in the normal and highly dynamic process of mitochon-
drial fusions. These results and some related investigations by Delivani et al.
(269) indicate that Bax and Bad coordinate both fusion and fi ssion of mito-
chondria in healthy cells. Karbowski and colleagues found that Bax/Bak
double knockout mice have fragmented but still functional mitochondria. In
contrast, Delivani et al. showed that overexpression of the anti - apoptotic
protein Bcl - xL (or the nematode homologue CED - 9) in mammalian cells
promoted mitochondrial fusion. A working model that will undoubtedly
become more complex in the future has Bax/Bak act on the factor Mnf2 which
is required for fusion. The absence of Bax/Bak in DKO mice can be compen-
sated for by overexpression of Mnf2. At the same time, Bax/Bak are postulated
to inhibit a factor that is involved in fi ssion, a process that proceeds unchecked
in the absence of these pro - apoptotic factors.
It may be too early to dismiss the idea that Bax monomers form aggregates
in the outer membrane that constitute the channel for the escape of cyto-
chrome c and other IMS proteins. The channels demonstrated in liposomes
may be in vitro artifacts, and so far they have been shown to allow small ions
and fl uorescent dyes to pass through, but the passage of a 50 - kDa protein may
be a greater challenge. Since we do not understand the process of mitochon-
drial fusion and fi ssion in much detail (see Chapter 3 ), we cannot readily
explain the release of the IMS proteins during apoptosis by invoking mecha-
nisms active in fusion. However, it should be noted that fusion, fi ssion, and
MOMP all involve major perturbations of the outer membrane (and the inner
membrane). Under conditions where normal fusion and fi ssion occur, the
process may be suffi ciently orderly/controlled to prevent the loss of any pro-
teins from the IMS, but excess recruitment of pro - apoptotic Bax may lead to
MITOCHONDRIA AND APOPTOSIS 397
398 MITOCHONDRIAL MUTATIONS AND DISEASE
large enough perturbations to allow leakage of such proteins. The process
could be autocatalytic as downstream caspases become activated.
7.6 FUNGAL SENESCENCE
Many fungi appear to be able to divide mitotically for an almost unlimited
period of time, growing into large clones whose size may be limited only by
physical parameters. In the wild, Armillaria bulbosa colonies in excess of 0.5 -
km diameter have been found. On the other hand, some strains in the labora-
tory exhibit the phenomenon of senescence, that is, they cannot be serially
transferred indefi nitely in culture. In a few prominent cases the phenomenon
has been investigated genetically. Since mitochondrial genomes were found to
play a large role in the process, it has been tempting to draw parallels with
aging in humans; but as our understanding has advanced, the resemblance is
only at the physiological level: the inability of cells to undergo further mitotic
divisions and a generalized cellular decay at the end. The best - studied exam-
ples have been expertly reviewed by Griffi ths (270, 271) , and some salient
features are appropriate for the present chapter. Although nuclear genes can
play a role, a common theme is that senescence in fungi is associated with
cytoplasmic inheritance. A triggering event appears to be an alteration in the
mitochondrial genome which eventually leads to a severe mitochondrial
degeneration and dysfunction and hence senescence.
There has been a renewed interest in using yeast as a model system for
life - span research, in connection with calorie restriction and the role of the
gene SIR2 (encoding a NAD - dependent deacetylase (272) . A full discussion
is beyond the scope of this book. It should be noted that there are two life
spans to be considered for yeast: (a) a replicative life span limiting the number
of times a given mother cell can produce buds and (b) a chronological life
span, which is a measure of the capacity of stationary cultures to remain viable
over time. Piper (273) discusses the relevance of studies with yeast to our
understanding of aging in more complex organisms.
The existence of mitochondrial plasmids in many organisms, and fungi in
particular, has already been discussed in Section 4.1.6 . Even at relatively high
copy numbers, they may act as relatively innocuous molecular parasites,
expressing functions primarily devoted to their own autonomous replication,
but they can also contribute to the phenomena of senescence and hypoviru-
lence. In Neurospora crassa and Neurospora intermedia , plasmids isolated
from wild strains have been shown to be responsible for senescence. The kalilo
plasmid ( ∼ 9 kb) isolated originally in Hawaii causes N. intermedia to stop
growing, and in the terminal stages, abnormalities in cytochrome spectra and
content, defi ciencies in large ribosomal subunits, and loss of cyanide - sensitive
respiration are observed. It is now clear that the plasmid can insert at one of
several locations in the mitochondrial DNA. There may be more than one
insertion in the process of senescence, and an mtDNA population may evolve
until at death a predominant insert is observed. A similar picture is seen with
the maranhar plasmid isolated in an N. crassa strain in India, as well as with
other plasmids found in a large sample of natural isolates (274) . While there
appears to be no doubt that the kalilo and maranhar plasmids are the respon-
sible agents, other plasmids in Neurospora are neutral. The challenge therefore
is to understand why some plasmids can and do insert themselves in the
mtDNA. According to Griffi ths, the “ initial insertion event is probably best
viewed as a mistake. ” The mistake, however, is irreversible and ultimately
lethal. Does it have any biological signifi cance, especially outside of the
laboratory?
Historically, senescence in the fungus Podospora anserina was one of the
fi rst to attract attention, since all natural isolates died, but they each exhibited
distinct and reproducible life spans. Cytoplasmic inheritance was also observed,
but heterologous, extragenomic mitochondrial plasmids cannot be detected.
Instead, as senescence progresses, circular DNA molecules derived from the
mtDNA are observed. The triggering event is unclear, but the net effect is that
one of several different segments of mtDNA (labeled α , β , γ , etc.) becomes
excised, associated as head - to - tail multimers, and circularized to form a series
of senDNAs. At the same time, intact mtDNA molecules disappear. The most
common sequence is found in a - senDNA, which consists of monomers identi-
cal in sequence to the fi rst intron of the CO1 gene (cytochrome oxidase). It
may be signifi cant that this group II intron encodes a protein with homology
to reverse transcriptase, but generalizations of the mechanisms for senescence
in Podospora anserina must include an explanation for the β - , γ - , δ - , etc.
senDNAs, which are not restricted to introns and do not encode obvious func-
tions (270) . Curiously, a senescing culture can be rejuvenated by treatment
with ethidium bromide, as if the intercalator were capable of selecting for
wild - type mtDNA molecules, since the sen plasmid present at the time of
addition is eliminated. When ethidium bromide is removed again, senescence
recurs in association with the appearance of a different senDNA type.
Numerous studies have sought to identify strains with different life spans,
and to identify nuclear or mitochondrial mutations responsible for this varia-
tion. Details can be found in the reviews by Griffi ths (270, 271) and by Bertrand
(275) . For example, a short - lived strain dies only on solid medium, but not in
liquid culture. After repeated transfers in liquid cultures, immortal cultures
could be obtained. These were female sterile and incapable of transmitting the
immortality as males. Mitochondrial mutations were likely responsible for the
altered phenotype. A publication on Podospora anserina (276) can serve for
an interested reader as an entry point into the recent literature.
In conclusion, senescence in Podospora anserina can be linked to mitochon-
drial DNA instability and mitochondrial energy metabolism; but here also the
triggering mechanism, the transition from the nonsenescent (juvenile) state to
the senescent state, remains to be elucidated. As in the case of the Neurospora
plasmids, the true biological signifi cance of the phenomenon of this induced
aging remains another puzzle.
FUNGAL SENESCENCE 399
400 MITOCHONDRIAL MUTATIONS AND DISEASE
7.7 CYTOPLASMIC MALE STERILITY IN PLANTS
Cytoplasmic male sterility (CMS) has been presented briefl y in a different
context in Chapter 4 . The integration of specifi c plasmids into mtDNA can
cause CMS in maize (277) . Another genetic mechanism, independent of the
existence of extragenomic plasmids, will be illustrated here with a select
number of examples. It should be noted that the phenomenon has been
observed in more than 150 plant species. It is widely exploited commercially
as an expedient method to produce pollen - sterile plants (278) . Some authors
have referred to it as a “ molecular arms race between two confl icting genomes
with different modes of inheritance (279, 280) . The involvement of the mito-
chondrial genome in all cases points to a crucial role of mitochondrial
function(s) in the development of the male reproductive system and pollen
formation in particular. The following discussion will give the reader a fl avor
of the kinds of molecular mechanisms being studied. It cannot be exhaustive,
but the reader will fi nd much up - to - date information in an expert review by
Chase (281) .
The exceptionally large mitochondrial genomes in plants have already been
discussed (Chapter 4 ) in terms of their high recombination rate, thought to
include a population of large circular mtDNA molecules, each representing a
portion of the genome. Specifi c “ recombination repeat ” sequences typically
greater than 2 kb are responsible for homologous recombinations capable of
permuting the genome without mutating the coding regions. Occasionally,
illegitimate (?) recombination events involving much shorter repeat sequences
can occur, and they may be responsible for the evolutionary changes in the
plant mt genomes. Even more rarely, such recombination events can create
new chimeric genes responsible for CMS. The search for such new sequence
arrangements in the large plant mtDNA is a formidable task.
L ’ Homme et al. (282) have thrown considerable light on the situation in the
oilseed rape (canola) species Brassica napus , and the results may exemplify
the general nature of the genetic changes associated with CMS (Figure 7.3 ).
The fertile B. napus plant has a cytoplasm (mitochondrial genome) referred
to as cam ; two infertile strains (male sterility) have cytoplasms (mitochondrial
genomes), nap and pol . The cam cytoplasm may have originated from the
diploid B. campestris progenitor of the amphidiploid B. napus . The nap cyto-
plasm makes plants male - sterile only when certain nuclear Rf s (restorer of
fertility) are missing, but the pol cytoplasm makes most B. napus cultivars male
sterile. The whole mtDNA sequence is not yet available, but many segments
containing known genes have been identifi ed and physical maps of restriction
fragments are available. The task was to fi nd differences between the cam, nap ,
and pol mt genomes.
Several lines of evidence have pointed at the vicinity of the atp6 gene as
the region differing in the cam, pol , and nap mtDNA. Detailed restriction
mapping, sequencing, and the analysis of chimeric transcripts found in the
various strains has yielded a picture of the arrangement of sequences in the