Barnes D.J., Chu D. Introduction to Modeling for Biosciences
Подождите немного. Документ загружается.


4.3 Differential Equations 153
Another, slightly clearer way of writing this is:
d
dt
(x +ay) =0 (4.25)
This means that the expression x +ay is constant in time, because the derivative
is constant. Since (4.25) is true at any time during the evolution of the system, it is
certainly true at time t =0. Denoting x(0) by x
0
we get:
x +ay =x
0
+ay
0
(To interpret this equation, it is useful to remember that x is an abbreviation
for x(t).) We can now use this equation to express y as a function of x.
y =
x
0
+ay
0
a
−
x
a
(4.26)
We have now nearly succeeded in decoupling the differential equations in (4.24).
All that remains to be done is to substitute the newly found expression for y into the
differential equation for ˙x, abbreviating x
0
+ay
0
by b.
˙x =bx −x
2
(4.27)
We have now genuinely reduced the coupled system (4.24) to a single differential
equation. We only need to solve this differential equation because we can then obtain
the solution for y(t) by using (4.26).
The solution, as follows, can be obtained by using the separation of variables
technique again.
x(t) =
bC exp(bt)
C exp(bt) −1
(4.28)
4.9 Use the separation of variables technique to show that the solution for x(t) is
(4.28).
We still need to determine the integration constant C. As usual, we do this by
specifying the initial conditions.
x(0) =x
0
=
(x
0
+y
0
)C
C −1
=⇒ C =
x
0
−x
0
−y
0
+x
0
=
x
0
x
0
−b
4.10 What happens when x
0
=b?
The steady state behavior can again be obtained by considering the behavior at
infinite times. In this case, the exponential terms become very large and the fraction
will approach 1. Therefore, at steady state, the size of the bacterial colony will be
given by,
x(∞) =b (4.29)
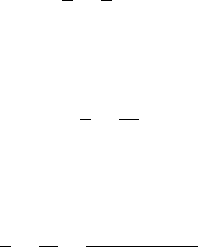
154 4 Differential Equations
We can obtain the same result directly from the differential equation (4.27) by ana-
lyzing the case of no change of the state variables, in this case ˙x =0.
˙x =0 =bx −x
2
=⇒ x =b
In addition to x
∗
= b the system also allows a trivial steady state given by x
∗
= 0
(steady states are commonly indicated by the variable name followed by a star).
Now that we have the solution for the time behavior of the bacterial colony, we
can also solve for the concentration of nutrients. For this we use our solution for
x(t) and (4.26).
y =
b
a
−
x
a
Just to convince ourselves of the soundness of the solution, let us check that the
initial and steady state values are correct.
y
∗
=y(∞) =
b
a
−
x
∗
a
=0
This can be obtained by substituting the steady state solution of x(t) for x
∗
.Forthe
initial value we get,
y
0
=y(0) =
b
a
−
x
0
a
=
x
0
+ay
0
−x
0
a
=y
0
For illustration, we plot the solution of our differential equation in Fig. 4.6 for
various initial conditions.
Note that, in this particular model the steady state depends on the initial con-
ditions. This is essentially due to the fact that there is a conserved property in the
model, as discussed above. This is not the typical case for differential equation mod-
els in biology. Normally, cells die or molecules are broken down. This introduces
dissipative elements into the model which prevent conservation relations from re-
maining valid. Yet, even when there are no conservation properties to be obeyed, it
can still be the case that the steady state depends on the initial conditions. This is
the case when the system supports more than a single steady state. This situation
partitions the phase space of the system into so-called basins of attraction for the
relevant steady states. We have already encountered a trivial example of this above,
in the case of the bacterial growth model. Any initial condition corresponding to no
bacteria being in the system causes the system to remain in this state forever. In this
instance, the basin of attraction corresponds to all states x(0) =0. We will see more
interesting examples of this below.
4.4 Case Study: Malaria
With these introductory remarks behind us let us now look in more detail at some-
what more advanced models. As a specific system let us consider a model of Malaria
4.4 Case Study: Malaria 155
Fig. 4.6 The dynamic behavior of (4.28)forx
0
= 0.01, a = 1 and various initial amounts of
nutrient
(or, in fact, any other vector borne disease) and its spread. Extensive details of a sim-
ilar model were given in Chap. 2, in the context of agent-based modeling. We have
vectors of infection (the mosquitoes) that infect humans by biting them; mosquitoes
are only infectious if they have been infected themselves. We assume that both
mosquitoes and humans remain infected for a certain amount of time before they
return to health again.
This rather qualitative information does not completely specify the differential
equation model we want to formulate, but it helps us to develop a first model that
will then enable us to obtain an idea of the behavior of the system. We approach the
modeling process as in the examples above; that is, we start by thinking about how
changes in the variables affect the spread of an infection.
Considering the infection dynamics for humans first. Qualitatively we can state
that the more infected mosquitoes there are, the more humans will get infected.
To be more precise: The greater the number of infected mosquitoes there are the
higher the chance per time unit that an uninfected human becomes infected. It is
unclear, at first, exactly what the dependence of the infection rate on the prevalence
of infection among mosquitoes is. As a first model and in the absence of better data,
we can assume linear proportionality. Since only uninfected humans can become
infected, we need to assume that the infection rates among humans also depend on
the number of uninfected individuals (rather than on the total number of humans).
Next we need to decide on how humans lose their infection. In reality, there are
two possible routes. Either one is cured or one dies. Once one has suffered a bout
156 4 Differential Equations
of Malaria one tends to acquire some immunity against the disease, making future
infections both less likely and less severe. Including this in a differential equation
model would require introducing a third category of humans beside the infected and
the uninfected ones. This is fully feasible, yet we are not primarily interested in
Malaria here, but in the general process of how to construct differential equation
models. Therefore, we will ignore this additional complicating factor. Similarly, we
will not explicitly model the fact that infected sufferers can die of the disease. Our
model population will be constant and we will assume that individuals lose their in-
fection after some time and return to the pool of uninfected individuals. Differential
equations can of course not keep track of individuals, but always consider large pop-
ulations. The way to model the “curing” process is to assume that infected humans
lose their infection at a certain constant rate. Formally, the problem is equivalent
to the case of protein decay above. In any time period, the actual number of curing
events will depend on the number of infected individuals as well as the curing rate.
Hence, altogether, the total rate of curing is proportional to the fixed curing rate
constant and the number of infected humans.
As far as the mosquitoes are concerned, we can make exactly the same arguments
regarding the infection and curing rates, although with different proportionality con-
stants. This reflects that the infection dynamics of mosquitoes should be assumed to
be distinct, if for no other reason than that they live for a much shorter time.
Before we can formulate our model mathematically, we need to pay attention to
one detail. As far as the infection rate is concerned, what counts is not the absolute
number of infected individuals, but the proportion of infected individuals in the
entire population. To reflect this aspect in our model let us denote the proportion
of infected humans by s(t); the proportion of healthy individuals is then given by
1 − s(t), assuming that there are no states between healthy and sick. Similarly, let
us denote the proportion of infected mosquitoes by m(t); in close analogy with the
human population, the proportion of uninfected individuals is given by 1 − m(t).
By adopting this form, we do not need to write separate formulas for infected and
uninfected populations, since the proportion of infected and uninfected individuals
can be immediately obtained from one another.
Taking all this into account, we arrive at a differential equation model for Malaria
infection.
˙s =a(1 −s)m −bs
˙m = c(1 −m)s −dm
(4.30)
This model formulates our assumptions about the factors affecting the growth of
infections in Malaria. The first term on the right hand side of the first line reflects
the fact that the growth of the infections among humans depends on the size of the
uninfected population (represented by 1 −s) and the size of the infected mosquito
population m. Note that other terms also have parameters, denoted by a, b, c and d.
These system parameters assign weights to the various growth and shrinkage terms
and normally need to be measured (that is, they cannot be inferred by an armchair
modeler).
In this particular model, the parameter c can be interpreted physically. It sum-
maries two effects, namely the probability of transmitting a disease should a human
4.4 Case Study: Malaria 157
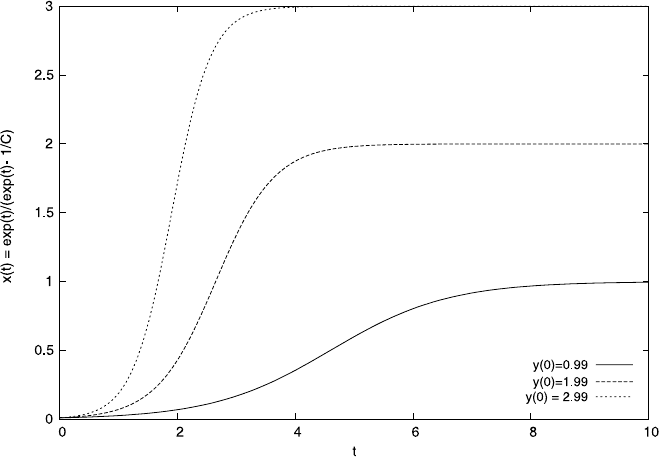
Fig. 4.7 A possible solution for the differential equation system (4.30) describing the spread of
Malaria. Here we kept the parameters, a =1, b =0.4, c =1, d = 2
be bitten by a mosquito, and the probability of a human being bitten in the first
place should human and mosquito “meet.” The parameter a is interpreted in an
analogous fashion. The parameters b and d determine the rate with which humans
and mosquitoes, respectively, lose their infection.
Unlike the simpler models we have encountered so far in this section, this one
cannot be solved analytically with elementary methods, and we will therefore not
attempt to do so. This is a pity, but it is the norm for the typical model one tends to
encounter in biology. Luckily, nowadays it is easy to obtain numerical solutions to
differential equation models using computer algebra systems, such as Maxima (see
Chap. 5). The disadvantage of this approach is that, in order to generate a solution,
we need to specify values for all the parameters of the model (a,b,c,d). Any so-
lution we generate is only valid for this particular set of parameter values, and the
behavior can be quite different for other sets of values (Fig. 4.7). Unless the parame-
ters are well specified, for example experimentally measured, one has to spend quite
some time exploring the possible behaviors of the model by generating numerical
solutions for many combinations of parameters.
Even when an analytic solution is not possible using elementary techniques
alone, one can still often extract some useful general information about the behav-
ior of the system by analyzing its steady state behavior. We can easily calculate the
steady state behavior by setting ˙s =0 and ˙m = 0 and solve the resulting system of
equations in (4.30). From this we obtain a solution for the steady state behavior of
158 4 Differential Equations
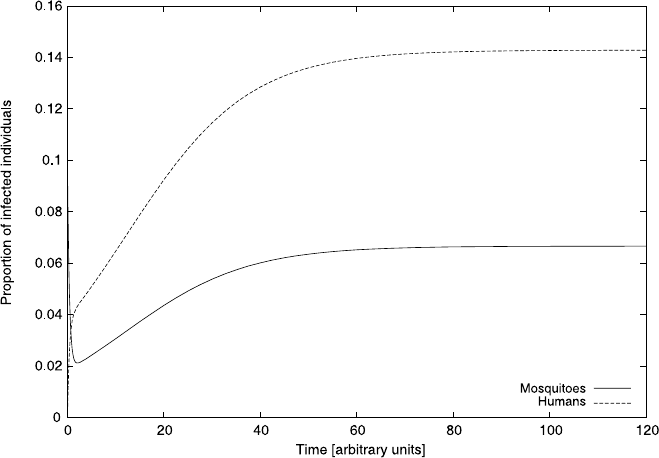
Fig. 4.8 The steady state infection levels for mosquitoes and humans of the model equation (4.30);
these two curves illustrate (4.31). Here we used the parameters, a = 1, c = 1, d = 2fixedand
varied b
the model in terms of the parameters.
s
∗
=
ca −db
c(a +b)
m
∗
=
ca −db
a(c +d)
(4.31)
Figure 4.8 illustrates how the steady state varies with b when we keep the other
parameters fixed at a =1, c = 1 and d =2. From the figure it is clear that positive
solutions can only be obtained when b<0.5. This can also be seen from (4.31).
If we substitute the chosen values of a,c and d into the first equation, then we
obtain the steady state number of infected humans, s
∗
, in terms of b. If we then
set s
∗
= 0 we obtain the value of the parameter b where the steady state reaches
zero. This point is exactly at b =
1
2
. For this value, or indeed larger values of b any
infection that may be introduced into the system will eventually disappear again.
Note, however, that a steady state corresponding to vanishing infection levels is
fully compatible with transient periods where Malaria is present in the system.
Figure 4.9 illustrates this. Here we set the parameter b =
1
2
, which means that
in the long run any infection will die out. The graph shows an example where at
time t = 0 all the humans and 10% of the mosquitoes are infected. Despite the
fact that the infection is not sustainable in the long run, initially the model even
leads to an increase of the infection levels among the mosquitoes. The levels fall
only after some time. In this particular case, the rates of decline of the infection are
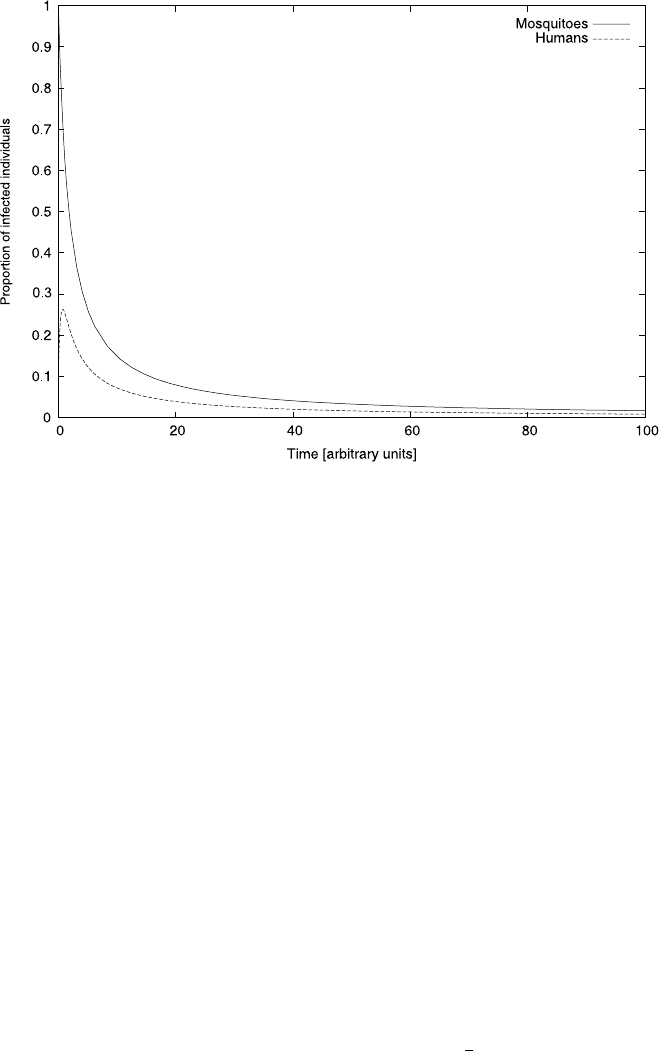
4.4 Case Study: Malaria 159
Fig. 4.9 The transient behavior of model equation (4.30). We started with initial conditions where
all humans and 10% of mosquitoes are infected. We used the parameters, a = 1, b = 0.5, c = 1,
d =2. Initially, the infection level of mosquitoes even rises before it eventually falls
very high at first, but eventually the fall in infections becomes slower and slower.
As a result, even after long transient periods there is still a low level of infection
in the system. Malaria will only be eradicated completely after infinite amounts of
time have passed although, in practice, infection levels will be very low after a long
but finite time. We have encountered this type of slow approach of the steady state
above.
In biological modeling, one is normally only interested in a subset of the math-
ematically possible solutions of differential equations. Typically one wishes to pre-
dict quantities such as concentrations of molecules, particle numbers or, as in the
present case, infection levels. These quantities are all described by positive real
numbers. Similarly, the space of possible or meaningful values of parameters is of-
ten restricted by the physical meaning. For example, as far as the Malaria model is
concerned, we are only interested in positive values for the four parameters a, b,
c and d of the system. These all represent proportionality constants and it would
not makes sense to set any of these to negative values, even though mathematically
there is no reason not to do this. A negative value for any of these parameters would
completely pervert the meaning of the model.
We should, however, spend some time thinking about the areas where our
model might predict negative infection rates, which would make no physical sense.
A glance at Fig. 4.8 reveals, rather troublingly, that for b>
1
2
there will be a negative
steady state. The reader is encouraged to convince herself of this using (4.31).
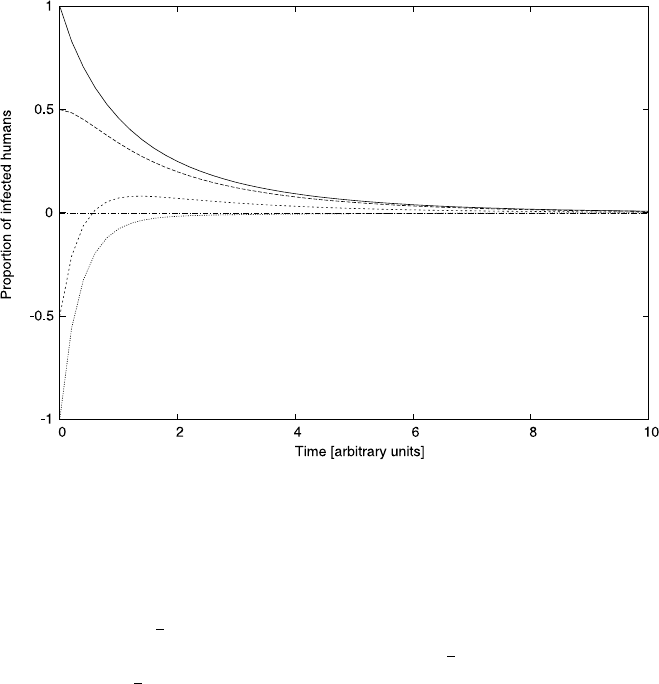
160 4 Differential Equations
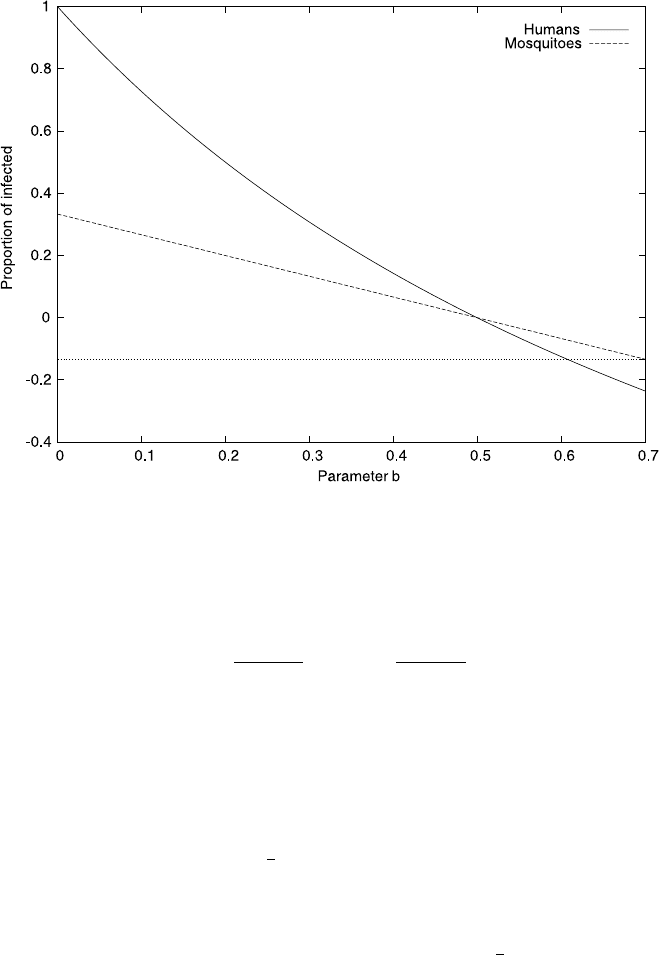
Fig. 4.10 The transient behavior of model equation (4.30) for various initial conditions. We used
the parameters, a =1, b =0.6, c =1, d =2, and an initial condition of m(0) =1
This is potentially a problem for the interpretation of the model with respect
to the real situation. There is nothing inherently unphysical or unreasonable about
assuming that b>
1
2
; the parameter b simply formulates how fast people recover
from Malaria. It might well be that in real systems b<
1
2
, but this does not change
the fact that b>
1
2
still is a meaningful choice of parameter. Still it seemingly leads
to the unphysical prediction of negative infection levels in the long run. If this was
indeed so, then this would indicate a problem of the model: There is no use having
a model that sometimes produces unrealistic behavior.
Let us see what happens when we increase the parameter b beyond 0.5. Fig-
ure 4.10 illustrates the dynamics predicted by the model for b =0.6 for various ini-
tial conditions ranging from s(0) =−1tos(0) = 1 (all graphs shown assume that
m(0) = 1). All the example curves we plotted predict that, in the long run, infec-
tions among humans will vanish, that is the negative steady state is not approached
after all. If we start with negative infection rates then, at least for a transient time
s(t) < 0 but again in the long run, infection levels tend to zero. The negative tran-
sient levels are neither surprising nor disconcerting. After all, by choosing initial
conditions corresponding to physically unrealistic values, we provided garbage as
input, and are accordingly rewarded with garbage as output. On the other hand, if
we start with the physically realistic initial conditions, that is 0 ≤s(0) ≤1 then the
model predicts positive infection rates throughout, only tending to zero in the long
run. This is reassuring. It is not watertight evidence for the correctness of the model
4.4 Case Study: Malaria 161
(whatever we mean by “correct”), but, at least it means that if fed on reasonable
input, the model does not produce meaningless output.
While we can feel a little reassured about the predictions of the model in this
particular case, we should start to have some doubts about the predictions of the
steady state behavior of the model. Following our steady state analysis in (4.31) and
Fig. 4.8 we would expect the model to show a negative steady state. This is clearly
not what we observe, at least in the example depicted in Fig. 4.10.
One immediate resolution for this apparent discrepancy is that the system allows
more than two steady states, as discussed above. One of them corresponds to vanish-
ing infection levels in both mosquitoes and humans, or simply s
∗
=m
∗
=0. While
trivial, this is a physically realistic steady state. The second, for b>0.5, corresponds
to physically infeasible solutions s
∗
< 0 and m
∗
< 0.
In order to understand the situation in more depth, it is necessary to introduce a
new concept, namely that of stability of a steady state point. Mathematically, much
is known about the stability of steady state points. A thorough treatment of the
topic that would do justice to the wealth of available knowledge is far beyond the
scope of this book. The reader who wishes to deepen her knowledge on this issue
is encouraged to consult a more advanced text such as Casti’s excellent and very
accessible two volume “Reality Rules” [5, 6]. Yet, even without entering the depths
of dynamical systems theory, it is possible to convey the main ideas. Despite being
elementary this basic understanding is often sufficient to master the demands of real
world modeling enterprises in biology.
4.4.1 A Brief Note on Stability
Let us assume a population of uninfected humans and mosquitoes, i.e., s(t) =
m(t) = 0. Clearly, unless Malaria is somehow actively introduced into the system,
the infection levels will remain at zero for all time. The question we want to ask
now is the following: What happens to the system if we introduce the infection into
the system at a small level, for example by releasing a few infected mosquitoes, or
by the arrival of a few people who carry Malaria?
There are two answers to this question: (i) It could be that an infection will gain
a foothold and establish itself. Alternatively, (ii) it might not establish itself and the
system reverts, after a short spell of Malaria, to the state where s(t) = m(t) = 0.
In essence, this is the intuition behind what mathematicians call the stability of a
steady state point of a system of differential equations: How does a system at steady
state respond to a small disturbance. The first case, where a small disturbance to
the steady state spreads and leads to the infection establishing itself in the long run,
represents the case where the steady state corresponding to m
∗
=s
∗
=0 is unstable.
The second case, where the disturbance does not spread, the steady state is stable. In
general, when modeling biological systems, we are interested in stable steady states,
because these are the ones we will observe in reality. Real systems are permanently
subject to more or less violent shocks that push them away from any unstable steady
162 4 Differential Equations
states they might inhabit. Unstable steady states persist just as long as a ping-pong
ball would rest at the top of a convex surface, or a pool cue would remain upright
balanced on its tip when exposed to a slight draught or vibration.
As it turns out, there is a fairly simple method to determine the stability of a
steady state. It is based on determining the eigenvalues (see Appendix A.5.1)ofthe
Jacobian matrix of the system at the steady state. Again, very much is known and
can be said about the stability of dynamical systems (this is what systems of differ-
ential equations are sometimes called). However, much can be done with elementary
methods that can be learned with relatively little effort. In the following paragraphs
we will therefore introduce some methods that can be applied without going into
the theory of the topic. Nevertheless, the reader is urged to eventually consult more
specialized volumes to acquire more sophisticated skills backed up by a thorough
understanding of the theoretical background.
Before describing how to determine the stability of steady states, we need to
briefly define what the Jacobian matrix is. Assume a system of differential equations
of the general form:
˙x
1
=f
1
(x
1
,...,x
n
) (4.32)
˙x
2
=f
2
(x
1
,...,x
n
) (4.33)
... (4.34)
˙x
n
=f
n
(x
1
,...,x
n
) (4.35)
This system of equations is nothing but a generalized form of the differential equa-
tions we have encountered before. In the Malaria model (4.30)wehavetotake
x
1
≡s and x
2
≡m, f
1
(s, m) ≡a(1 −s)m −bs and so on.
Using this form the Jacobian is defined as the n × n matrix of derivatives of f
i
with respect to x
j
.
J
i,j
(x) =
∂f
i
(x
1
,...,x
n
)
∂x
j
(4.36)
In the case of n =3, for example, the Jacobian would be a 3 ×3 matrix.
J(x
1
,x
2
,x
3
) =
⎡
⎢
⎢
⎣
∂
∂x
1
f
1
(x
1
,x
2
,x
3
)
∂
∂x
2
f
1
(x
1
,x
2
,x
3
)
∂
∂x
3
f
1
(x
1
,x
2
,x
3
)
∂
∂x
1
f
2
(x
1
,x
2
,x
3
)
∂
∂x
2
f
2
(x
1
,x
2
,x
3
)
∂
∂x
3
f
2
(x
1
,x
2
,x
3
)
∂
∂x
1
f
3
(x
1
,x
2
,x
3
)
∂
∂x
2
f
3
(x
1
,x
2
,x
3
)
∂
∂x
3
f
3
(x
1
,x
2
,x
3
)
⎤
⎥
⎥
⎦
(4.37)
The basic structure of a Jacobian matrix is very simple. It is a list of derivatives
of the right hand sides of the differential equations with respect to all the system
variables.
It is not the Jacobian directly that provides insight into the stability properties of
a steady state, x
∗
=(x
∗
1
,...,x
∗
n
), but rather the set of eigenvalues E of the Jacobian
matrix at the relevant steady state, i.e., the matrix J(x
∗
). The members in the set E
are numbers, possibly complex numbers, although we are primarily interested in the