Barnes D.J., Chu D. Introduction to Modeling for Biosciences
Подождите немного. Документ загружается.


4.5 Chemical Reactions 173
remaining agnostic about the specific value of h. This leads us to a system of two
differential equations.
˙y =−v
max
X
0
y
h
K
h
+y
h
˙z = v
max
X
0
y
h
K
h
+y
h
(4.44)
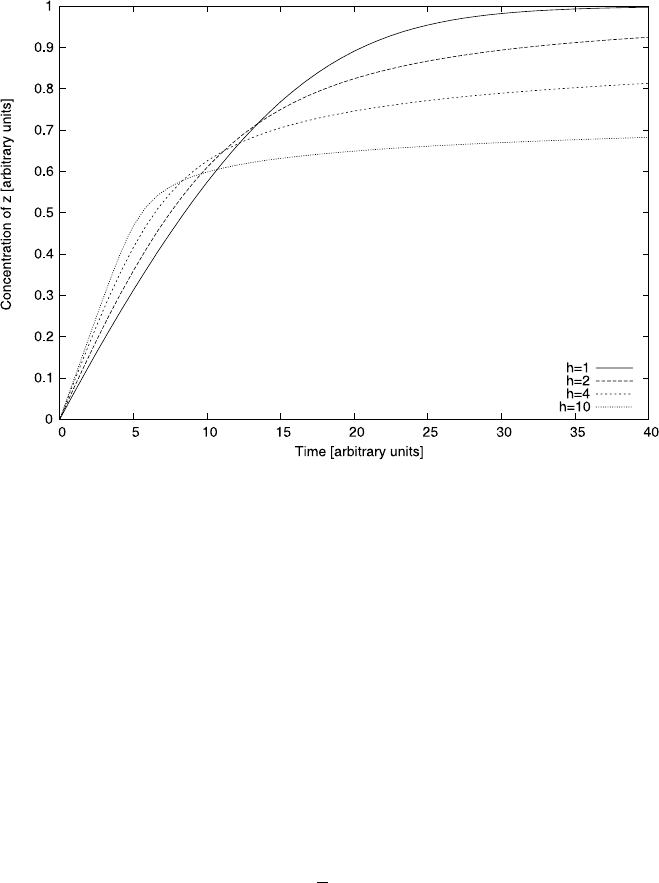
Figure 4.16 plots some solutions for this system assuming initial conditions of
y(0) =1 and z(0) =0. A salient feature of the graph is that the MM-kinetics leads
to initially slower growth. Yet, within the time period shown in the plot it clearly
leads to the highest total conversion. Initial growth is highest for the highest Hill
coefficient (i.e., h =10) shown in the plot, but this one also shows the poorest total
amount of converted substrate after the considered time period.
This qualitative behavior can be easily explained from the properties of the
MM/Hill kinetics. Hill functions become increasingly step-like as h increases (see
Fig. 4.15). For our system, this means that initially, when the substrate concentration
is at its maximum, Hill functions with a high h are much closer to their saturation
value of 1; the MM function, on the other hand, will be comparatively low, resulting
in a lower initial conversion rate. However, as the substrate is depleted and the con-
centration of y falls below y = K, a system with a higher Hill coefficient will see
the conversion rate drop to zero much more quickly. The MM-function, however,
is much less step like and relatively efficient conversion continues even if y is well
below K.
4.15 Formulate the system of differential equations corresponding to (4.44)but
under the assumption that y(t) is replenished at a constant rate.
4.16 What happens qualitatively to the solution of the system (4.44) for very small
and very large values of K. Confirm your conjectures by explicit numerical solu-
tions?
4.17 What happens to the solution of the system (4.44) when t →∞?
4.18 For the system (4.44), plot the conversion rate as a function of time.
4.5.2 Modeling Gene Expression
Enzyme kinetics is an important aspect of biological process modeling. Beyond that,
Hill dynamics is also commonly used to model transcription-factor (TF) regulated
gene expression. As in the case of enzyme kinetics, the use of the MM/Hill function
to describe the regulation of genes relies on the assumption of a separation of time-
scales. We need to assume that binding of TFs to and from the operator site happens
174 4 Differential Equations
Fig. 4.16 A numerical solution for the differential equation (4.44)forv
max
= 1, K = 0.1and
x = 0.1. Note that for higher Hill coefficients it takes longer for the substrate to be used up
much faster than expression of gene products. Generally, this assumption is true. We
will later explicitly derive the Hill equation for gene expression (see Sect. 6.2.2).
For the moment, we have to ask the reader to invest faith in the correctness of our
assertions.
Assume that we have a single gene that is expressed into a product x at a rate a.
To make the model somewhat meaningful, we have to assume that the product is
broken down with a rate b to avoid trivial and unrealistic unbounded growth rates
of the concentration of the product. We have already formulated this differential
equation above.
˙x =a −bx (4.19)
We also have found the general solution to this system.
x(t) =
a
b
+Ce
−bt
(4.20)
This is satisfactory as a very simple model of an unregulated gene. It does not cap-
ture the more important case of a regulated gene. In order to model regulated genes,
we need to extend the model to include the action of a TF Y (that is present with a
concentration y) on the rate of gene expression.
To begin with, let us assume that the promoter of G
x
, the gene encoding pro-
tein X, is controlled by a single binding site for the TF Y which binds to its binding
site with a rate constant of k
on
and dissociates with a constant k
off
. For the moment,

4.5 Chemical Reactions 175
let us also assume that Y is an activator. The gene G
x
will then operate in two pos-
sible regimes, depending on whether Y is bound or not. If it is bound, then it will
express X at a high rate a
H
; otherwise it will only express Y at a “leak” rate a
L
,
where a
L
<a
H
. Both cases can be described separately by a differential equation
of the form of (4.19). If we know the probability P
b
(y) that the TF is bound to its
binding site, then we can combine the two differential equations into one.
˙x =(1 −P
b
(y))a
L
+P
b
(y)a
H
−bx
= a
L
+P
b
(y)(a
H
−a
L
) −bx
The first line just expresses the intuition that if the TF is bound to the operator site
with a probability of P
b
(y) then the gene will express at a high rate for exactly
this proportion of time. The second line is only a rearrangement of the terms on
the first line. The question we need to answer now is how we can express P
b
(y)
mathematically. As it turns out, the MM function can be used to approximate this
probability.
P
b
(y) =
y
y +K
if K =
k
off
k
on
In this case, the MM-constant K is defined by the ratio of the unbinding rate and
binding rate of the TF to its operator site. Altogether we thus obtain a differential
equation describing the expression of a protein from a gene.
˙x =α +β
y
y +K
−bx (4.45)
Here we renamed a
L
and a
H
−a
L
to α and β respectively, for reasons of notational
simplicity. While we have not derived this expression from first principles, at least
for extreme parameters it seems to make sense. For example, if we assume that
the TF exists in an infinite concentration y, then Y is continuously bound to the
operator site. In this case the expression rate will be α +β. However, particularly
for smaller K, the actual expression rate will be close to this value even for y<∞.
In reality, the fraction of time the TF is bound to its operator site will always be
slightly below one, but probably by not very much. Clearly, the higher the binding
rate k
on
of Y to its operator site in relation to the unbinding rate, the smaller is K
and (near) continuous occupation of the binding site can be achieved by lower TF
concentrations.
4.19 Write down and solve numerically the full “unpacked” system of differential
equations corresponding to (4.45), i.e., write out the system of differential equa-
tions for which the MM/Hill function is an approximation. Explore its dynamics
and compare it to the solutions to (4.45).
In many cases a particular operator site has more than one binding site for a
particular TF. Modeling these cases can be somewhat more intricate when each
of the possible occupation configurations of the binding sites is associated with

176 4 Differential Equations
a different expression rate. To consider a simple case, assume that we have two
binding sites and that there are only two possible expression rates, namely, the leak
rate when zero or one of the binding sites are occupied, and a high rate if both
are occupied. Remembering that the MM-function formulates the probability of a
single binding site being occupied, we can now write down the rate equation for our
system.
˙x =α +β
y
N
(y +K)
N
−bx (4.46)
The high expression rate is only relevant when all N binding sites are occupied,
which happens with a probability of
y
N
(y+K)
N
; otherwise the gene is expressed at the
leak rate. While mathematically correct, this form of the kinetic equation is rather
unusual and instead the gene activation function in the presence of several binding
sites of a TF is more often assumed to be of the Hill form.
˙x =α +β
y
h
y
h
+K
h
−bx
Similar to the case of enzyme kinetics, the Hill coefficient is normally measured
rather than derived from first principles and the Hill coefficient carries some infor-
mation about the cooperativity between the binding dynamics of the underlying TF
binding sites. If the TF is a monomer and it occupies N different binding sites on
the operator, then h ≤ N . The Hill coefficient h will reach its maximum h = N in
the case when there is perfect cooperativity between the sites. This means that the
DNA-TF compound is unstable unless all binding sites are occupied. In this case,
binding can only happen if N TF bind to the N binding sites simultaneously, or
at least within a very short window of time. Perfect cooperativity of this kind is
primarily a mathematical limiting case and in practice there will be imperfect co-
operativity. This means that there will be a finite (but probably small) stability of
the DNA-nucleotide compound even if only some of the N binding sites are occu-
pied. The maximum binding times will only be achieved when all binding sites are
occupied. In these cases the Hill coefficient will be smaller than N.
Often, TFs are themselves not monomers but dimers or higher order compounds.
In this case, if the Hill function is formulated in terms of the concentration of the
monomer form of the TF, then h could be higher than the number of binding sites.
So far we have assumed that the TF is an activator; there is also a commonly used
gene activation function for repressors, that is if the gene G
x
is repressed by the TF
species Y .
˙x =α +β
K
h
y
h
+K
h
−bx
The Hill repressor function is simply 1−h(x) and thus shares the general qualitative
features of the original function. Gene expression is highest when the concentration
of the repressor y = 0; for very high concentrations of the repressor there is only
leak expression activity α.

4.6 Case Study: Cherry and Adler’s Bistable Switch 177
Gene activation functions can be extended to describe a gene being regulated
by multiple TFs. This is generally not difficult, but care must be taken to properly
describe how the interaction between the TFs works. For example, if we have two
repressor species Y
1
and Y
2
and both can independently down-regulate gene expres-
sion, then the following differential equation would be a possible way to capture the
behavior.
˙x =α +β
K
h
y
h
1
+y
h
2
+K
h
−bx
Alternatively, if Y
1
and Y
2
only partially repress the expression of the gene, then it
would be more appropriate to describe the gene activation function as a sum of two
Hill repressors.
˙x =α +β
1
K
h
1
β
y
h
1
+K
h
1
+β
2
K
h
2
β
y
h
2
+K
h
2
−bx
The specific function used will depend on the particular biological context of the
system that is to be modeled.
4.20 Write down an activation function for a gene that is activated by Y
1
but re-
pressed by Y
2
. Formulate the various possibilities for how the repressor and the
activator can interact.
4.21 Formulate the gene activation function for the above case when the activator
and the repressor share the same binding site (competitive binding).
4.6 Case Study: Cherry and Adler’s Bistable Switch
Let us now look at an example to illustrate some of the ideas we have developed.
A relatively simple yet dynamically very interesting example is the bistable genetic
switch that was first described by Cherry and Adler [8]. The system consists of
two genes that repress one another via their products. Let us assume the first gene
G
x
produces a protein X that represses the expression of gene G
y
producing Y .
Similarly, Y itself also represses the expression of G
x
. We can immediately write
down the differential equations describing the system.
˙x =
K
h
K
h
+y
h
−bx
˙y =
K
h
K
h
+x
h
−by
(4.47)
Here we make the special assumption that the system is symmetric in the sense that
the gene activation function of both genes has the same parameters K and h and
that both products are degraded at the same rate b.
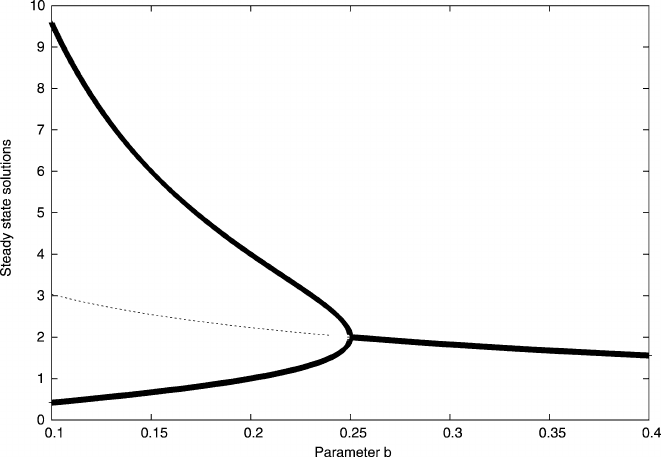
178 4 Differential Equations
Fig. 4.17 The solutions for the system of two mutually repressing genes as defined in (4.47). We
keep K =2 and vary the parameter b
The system is too complicated to be solved analytically and it is necessary to
find the solutions numerically. Given the inherent symmetry of the system, that is
G
x
and G
y
are dynamically identical, we would expect that the solutions for x and
y are always the same. As it will turn out though, for some parameters of the system,
x and y can have different steady state solutions. Figure 4.17 plots the steady state
solutions of the system for a particular choice of parameters. Normally, we would
have to specify which variables we consider but, in the present case, this is not nec-
essary; the symmetry of the model means that x and y are always interchangeable,
which does not mean that their values are always equal. In Fig. 4.17 the vertical
axis records the possible steady state values as the parameter b is varied (always
assuming a fixed K = 2).
At the high end of b the system apparently allows a single solution only. Since
we have two variables, but only one possible solution, this must mean that in this
area x
∗
= y
∗
.Atb<0.25 the single steady state splits into three solutions. The
dashed line are steady state solutions where x
∗
=y
∗
=s
u
; as it will turn out, these
solutions are unstable solutions and we would not expect them to be observed in
a real system. Any tiny fluctuation away from this steady state would lead to the
system approaching one of the other solutions.
The stable solutions s
L
and s
H
of the system correspond to the top and bottom
(thick) lines. In this area, if one of the variables, say x, takes the steady state x
∗
=s
L
then y will take the other stable state y
∗
= s
H
. Yet both steady states are possible
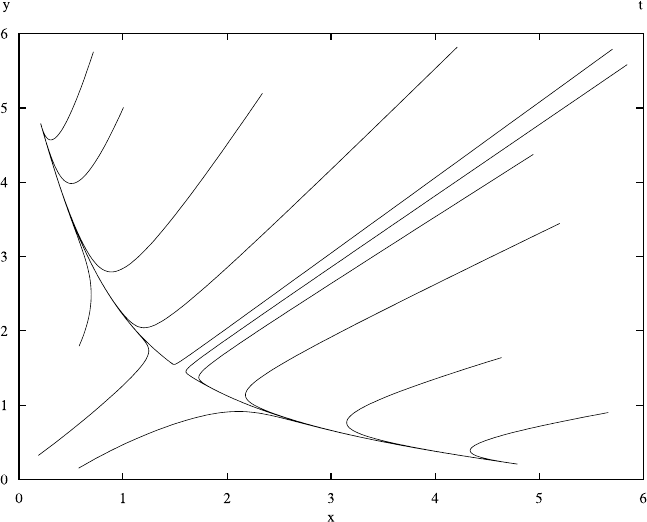
4.6 Case Study: Cherry and Adler’s Bistable Switch 179
Fig. 4.18 The phase-space representation of solutions for (4.47). The parameters are K =2and
b = 0.2. The horizontal axis represents x and the vertical axis represents y. Each line corresponds
to a trajectory of the system starting from an initial condition. Depending on the initial condition,
the system ends up in one of the two steady states. Compare this with the solutions plotted in
Fig. 4.17
for both variables. Which one is taken depends exclusively on the initial conditions
of the system.
Biologically this can be easily understood. If, say, G
x
is expressed at a high
rate, then this means that the concentration of X is high in the cell. Since X is
repressing Y , this also implies that the gene G
y
will not be expressed at a high rate,
but is repressed. Intuitively, one can thus easily understand how the symmetry of the
system is compatible with the products having two different steady states. However,
note that whether or not X or Y is expressed at a high rate will depend on the initial
conditions of the system. If we start with a high concentration of X then we would
expect that Y is suppressed and that the system remains in this steady state. On the
other hand, if we start with Y initially being higher, the system will remember that
and remain in a steady state defined by a low concentration of X.Thisisalsohow
we need to read the results in Fig. 4.17.
The dependence of the steady state on the initial conditions is illustrated in
Figs. 4.18 and 4.19. Figure 4.18 shows solutions for the differential equations in
phase space. The phase space shows x versus y; each point on each of the lines
in this diagram corresponds to a particular time. The various curves in this figure
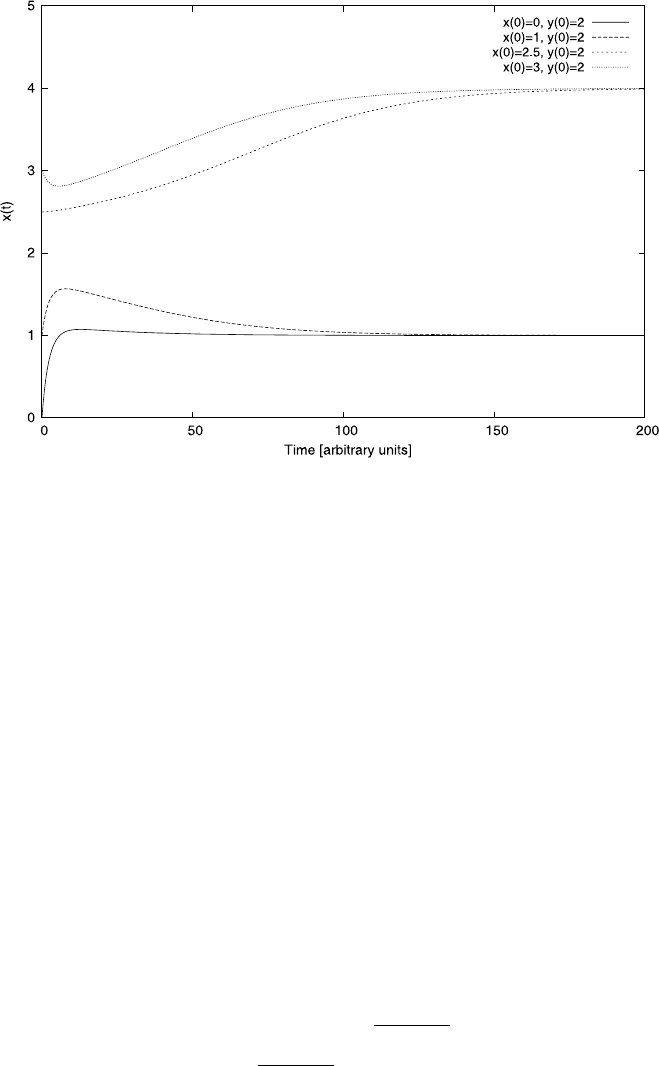
180 4 Differential Equations
Fig. 4.19 Some sample solutions of (4.47) for different initial conditions. The parameters are
K =2andb = 0.2 and we keep y(0) =2. If x(0)>y(0) then it assumes the higher steady state,
otherwise the lower. Compare this with Figs. 4.17 and 4.18
represent trajectories of the system that starts at some point and then move to one
of the steady states. The sample curves in this phase space diagram show a clear
partition of the space into two halves. The curves that originate above the diagonal
defined by x =y end up in the steady state in the upper right corner of the figure.
This steady state corresponds to G
y
suppressing G
x
or a high concentration of Y
and a low concentration of X. Trajectories that start below the diagonal end up in a
steady state where G
x
represses G
y
.
It is instructive to compare this phase-space representation with the normal time
solutions of x(t) showninFig.4.19. In all curves in this figure we kept y(0) = 2.
Depending on the initial conditions, the variable x(t) tends to the high steady state,
when x(0)>y(0) and to the low state otherwise. The graph does not show y(t),
which would take the steady state x does not take.
Let us now obtain another perspective on the system and consider the stability
properties of the steady state solutions. Based on our numerical examples, we would
expect that the steady state x
∗
=y
∗
is only stable when it is the only solution. If the
parameters allow another stable state x
∗
=y
∗
then we expect that one to be stable.
To ascertain this we first need to calculate the Jacobian.
J =
⎡
⎣
−b −
K
h
y
h
h
(y
h
+K
h
)
2
y
−
K
h
x
h
h
(x
h
+K
h
)
2
x
−b
⎤
⎦
(4.48)

4.6 Case Study: Cherry and Adler’s Bistable Switch 181
Let us now analyze the stability of the system using the same parameters as in
Fig. 4.19. We have calculated that this particular choice allows two stable steady
states which we (arbitrarily) assign to x
∗
= 4 and y
∗
= 1 (we could have done
this the other way around as well); Fig. 4.19 shows how these steady states are
approached. To obtain the stability of the first steady state, we need to set in the
values of x
∗
= 4 and y
∗
= 1 along with the parameter values K = 2 and b = 0.2.
Calculating the eigenvalues of the resulting matrix then gives: e
1
=−0.04 and e
2
=
−0.36. Both eigenvalues are negative, which confirms that the steady state is stable.
The second steady state where X is repressed gives the same result, as we would
expect.
We still need to check the stability of the steady state given by x
∗
= y
∗
=
2.229494219. Again, substitution of this value into the Jacobian and calculating the
eigenvalues yields e
1
≈0.02164 and e
2
≈−0.42164. Since one of the eigenvalues
is positive, this confirms that the steady state is unstable, as suspected.
In this particular case of a system consisting of two differential equations only,
there is another way to represent the behavior of the system. The nullclines are the
curves obtained by individually setting the expressions on the right hand sides of
the differential equation (4.47) to zero. They essentially show the possible steady
states for each of the two equations separately; the actual steady states are at the
intersections of the nullclines.
x =
K
h
b(K
h
+y
h
)
x = K
1 −by
by
1
h
Figure 4.20 plots these curves for the particular case of K = 2 and b = 0.2. The
three possible positive and real steady states correspond to intersection points of
the nullclines. From a computational point of view, it is often much easier to plot
the nullclines rather than to explicitly solve the equations to obtain the steady state.
Furthermore, the nullcline equations can also provide a better intuition as to which
parameters influence the number and location of steady states.
4.22 Formulate the differential equation for a self-activating gene, that is a gene
whose product activates its own expression. Assume that the gene product is broken
down at a rate of b. Write down the differential equation and analyze the number of
steady states.
4.23 Formulate the system of differential equations for the import of a nutrient S
that exists in the environment at a concentration s by a gram-positive bacterium.
Assume that it is taken up through porins into the periplasm first and then into the
cytoplasm by different porins. Assume that uptake by porins obeys Hill-dynamics.
Explore different model assumptions, including that ˙s =0.
4.24 Use the previous model, and vary the volume of the periplasm and the cyto-
plasm. What are the effects?

182 4 Differential Equations
Fig. 4.20 The nullclines for the system in (4.47). The parameters are K = 2andb = 0.2. The
horizontal axis represents x and the vertical axis represents y. The intersections between the lines
represent steady states. Compare this with the solutions plotted in Fig. 4.17
4.7 Summary
Differential equations are powerful tools in biological modeling. In this chapter we
have refreshed the reader’s understanding of the basic ideas of differential calculus.
We have also introduced some basic methods of solving the differential equations.
The material presented here necessarily barely scratches the surface of the body of
available knowledge, yet we believe that it should be sufficient to help in formu-
lating a number of biologically interesting models. Eventually, however, the reader
may wish to consider deepening her knowledge of the subject by consulting more
specialist volumes.