Berg J.M., Tymoczko J.L., Stryer L. Biochemistry
Подождите немного. Документ загружается.

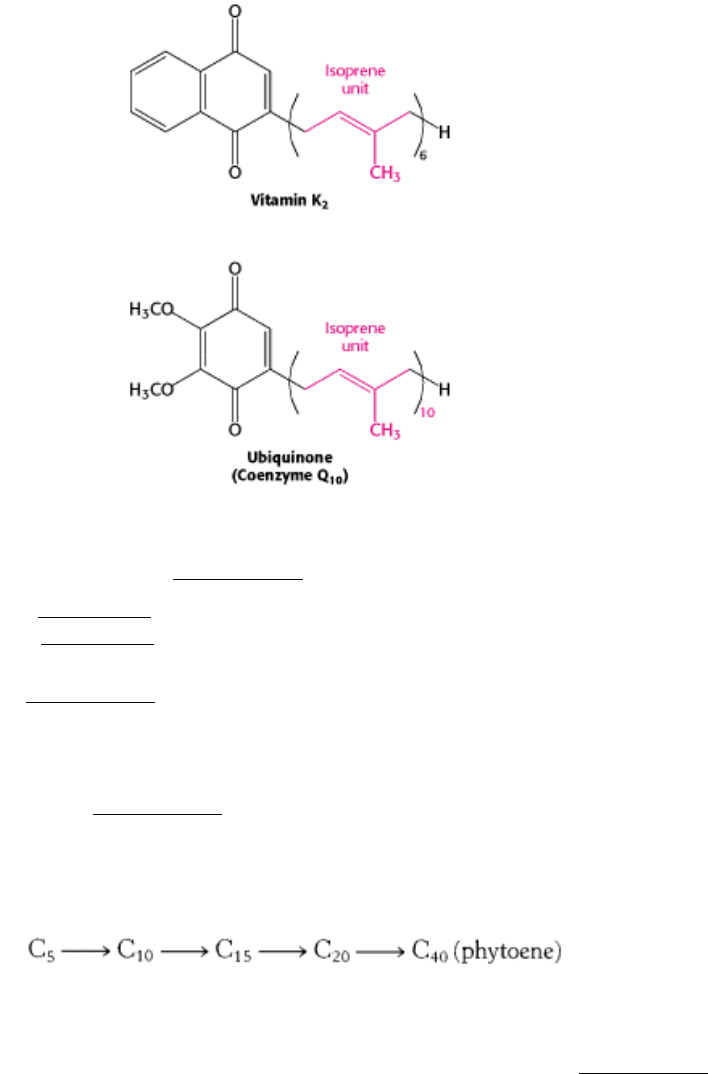
We have already encountered several molecules that contain isoprenoid side chains. The C
30
hydrocarbon side chain of
vitamin K
2
, an important molecule in clotting (Section 10.5.7), is built from 6 isoprene (C
5
) units. Coenzyme Q
10
in the
mitochondrial respiratory chain (Section 18.3) has a side chain made up of 10 isoprene units. Yet another example is the
phytol side chain of chlorophyll (Section 19.2), which is formed from 4 isoprene units. Many proteins are targeted to
membranes by the covalent attachment of a farnesyl (C
15
) or a geranylgeranyl (C
20
) unit to the carboxyl-terminal
cysteine residue of the protein (Section 12.5.3). The attachment of isoprenoid side chains confers hydrophobic character.
Isoprenoids can delight by their color as well as by their fragrance. The color of tomatoes and carrots comes from
carotenoids. These compounds absorb light because they contain extended networks of single and double bonds and are
important pigments in photosynthesis (Section 19.5.2). Their C
40
carbon skeletons are built by the successive addition of
C
5
units to form geranylgeranyl pyrophosphate, a C
20
intermediate, which then condenses tail-to-tail with another
molecule of geranylgeranyl pyrophosphate.
Phytoene, the C
40
condensation product, is dehydrogenated to yield lycopene. Cyclization of both ends of lycopene gives
β-carotene, which is the precursor of retinal, the chromophore in all known visual pigments (Section 32.3.2). These
examples illustrate the fundamental role of isopentenyl pyrophosphate in the assembly of extended carbon skeletons of
biomolecules. It is evident that isoprenoids are ubiquitous in nature and have diverse significant roles, including the
enhancement of the sensuality of life.
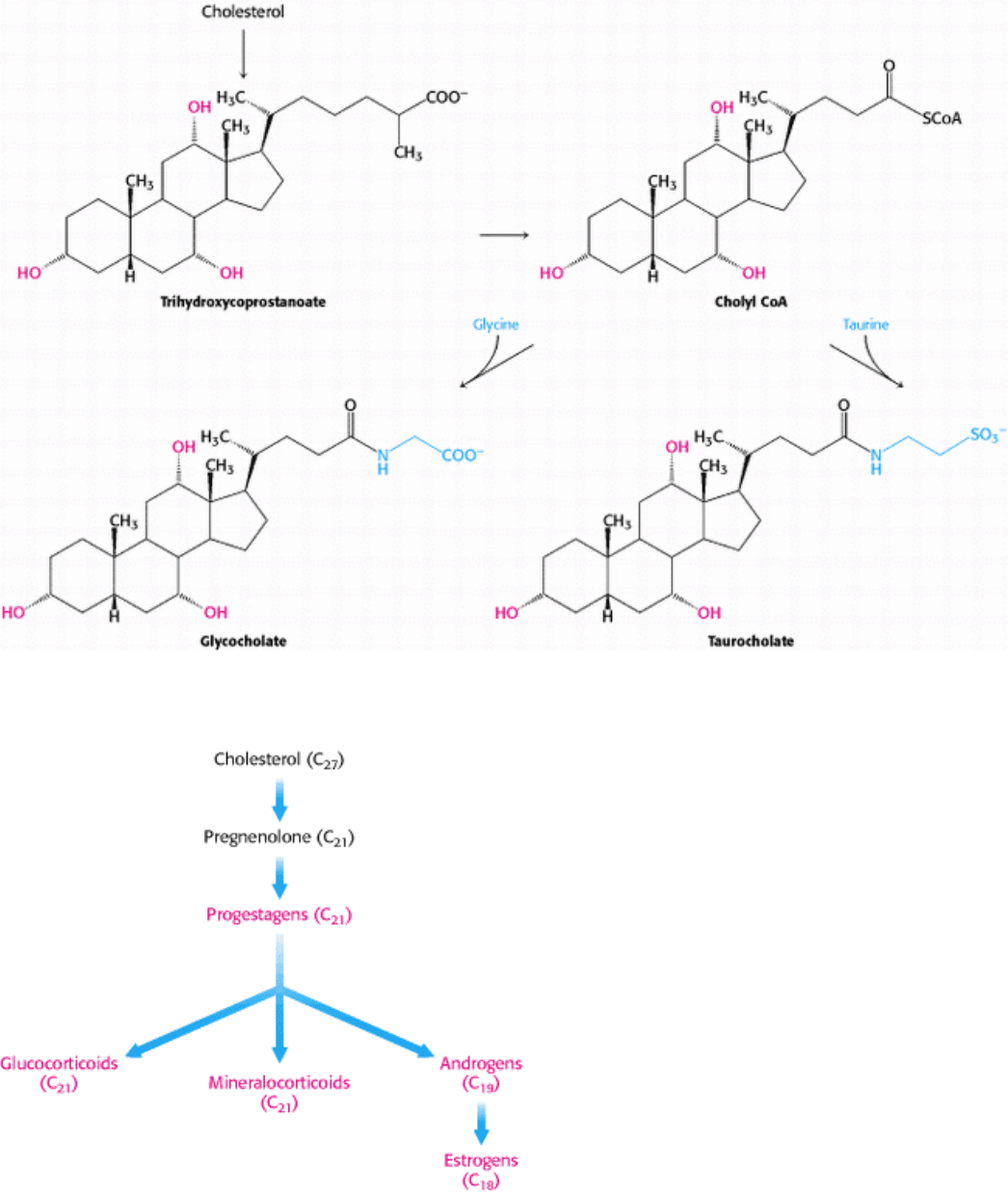
III. Synthesizing the Molecules of Life 26. The Biosynthesis of Membrane Lipids and Steroids 26.4. Important Derivatives of Cholesterol Include Bile Salts and Steroid Hormones
Figure 26.23. Synthesis of Bile Salts. Pathways for the formation of bile salts from cholesterol.
III. Synthesizing the Molecules of Life 26. The Biosynthesis of Membrane Lipids and Steroids 26.4. Important Derivatives of Cholesterol Include Bile Salts and Steroid Hormones
Figure 26.24. Biosynthetic Relations of Classes of Steroid Hormones and Cholesterol.
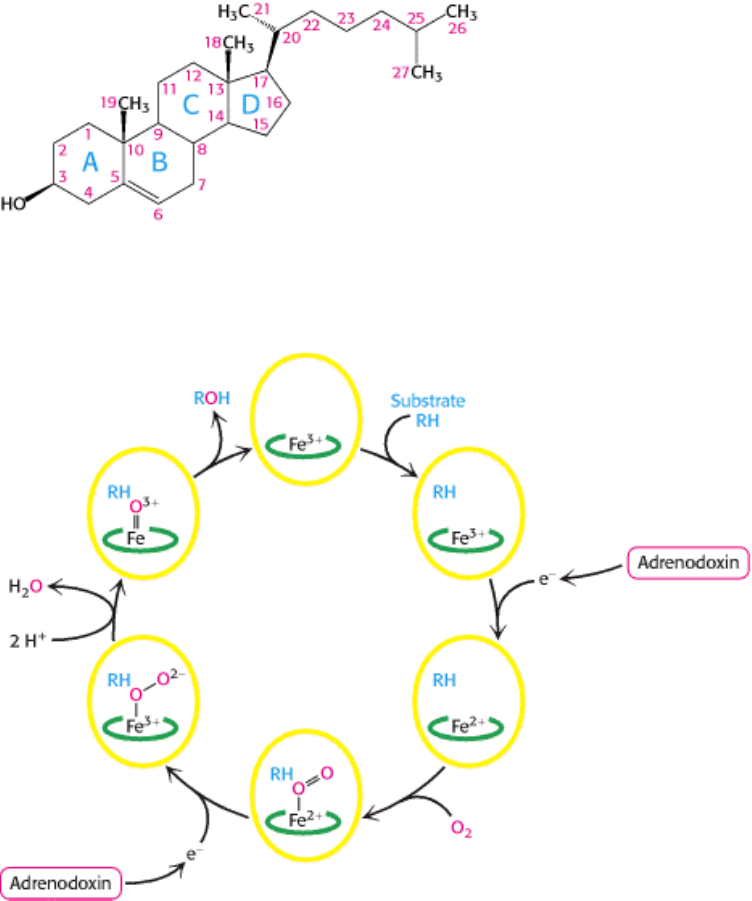
III. Synthesizing the Molecules of Life 26. The Biosynthesis of Membrane Lipids and Steroids 26.4. Important Derivatives of Cholesterol Include Bile Salts and Steroid Hormones
Figure 26.25. Cholesterol Carbon Numbering. The numbering scheme for the carbon atoms in cholesterol and other
steroids.
III. Synthesizing the Molecules of Life 26. The Biosynthesis of Membrane Lipids and Steroids 26.4. Important Derivatives of Cholesterol Include Bile Salts and Steroid Hormones
Figure 26.26. Cytochrome P450 Mechanism. These enzyme-bind O
2
and use one oxygen atom to hydroxylate their
substrates.
III. Synthesizing the Molecules of Life 26. The Biosynthesis of Membrane Lipids and Steroids 26.4. Important Derivatives of Cholesterol Include Bile Salts and Steroid Hormones
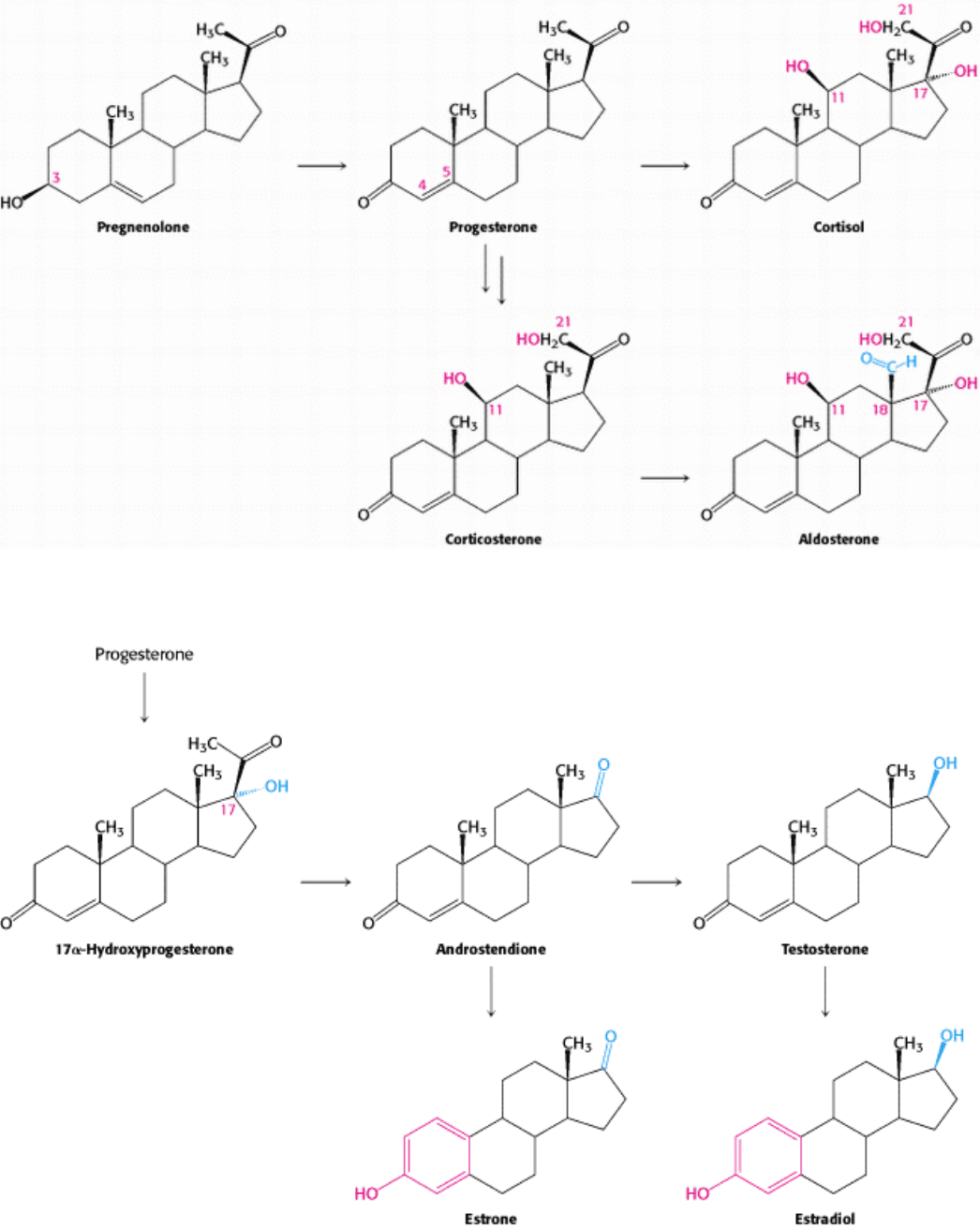
Figure 26.27. Pathways for the Formation of Progesterone, Cortisol, and Aldosterone.
III. Synthesizing the Molecules of Life 26. The Biosynthesis of Membrane Lipids and Steroids 26.4. Important Derivatives of Cholesterol Include Bile Salts and Steroid Hormones
Figure 26.28. Pathways for the Formation for Androgens and Estrogens.
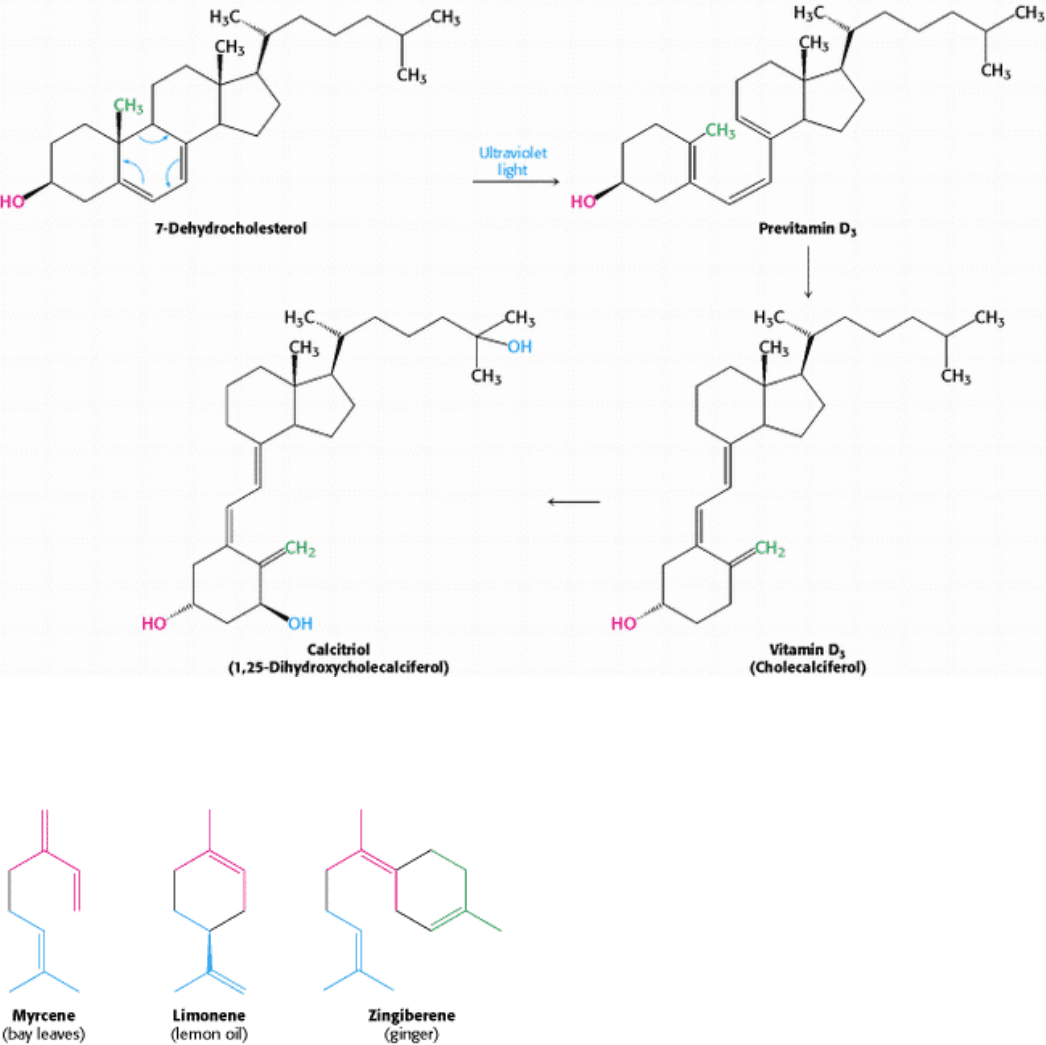
III. Synthesizing the Molecules of Life 26. The Biosynthesis of Membrane Lipids and Steroids 26.4. Important Derivatives of Cholesterol Include Bile Salts and Steroid Hormones
Figure 26.29. Vitamin D Synthesis. The pathway for the conversion of 7-dehydrocholesterol into vitamin D
3
and then
into calcitriol, the active hormone.
III. Synthesizing the Molecules of Life 26. The Biosynthesis of Membrane Lipids and Steroids 26.4. Important Derivatives of Cholesterol Include Bile Salts and Steroid Hormones
Figure 26.30. Three Isoprenoids from Familiar Sources.
III. Synthesizing the Molecules of Life 26. The Biosynthesis of Membrane Lipids and Steroids
Summary
Phosphatidate Is a Common Intermediate in the Synthesis of Phospholipids and
Triacylglycerols
Phosphatidate is formed by successive acylations of glycerol 3-phosphate by acyl CoA. Hydrolysis of its phosphoryl
group followed by acyl- ation yields a triacylglycerol. CDP-diacylglycerol, the activated intermediate in the de novo
synthesis of several phosphoglycerides, is formed from phosphatidate and CTP. The activated phosphatidyl unit is then
transferred to the hydroxyl group of a polar alcohol, such as serine, to form a phospholipid such as phosphatidyl serine.
In bacteria, decarboxylation of this phosphoglyceride yields phosphatidyl ethanolamine, which is methylated by S-
adenosylmethionine to form phosphatidyl choline. In mammals, this phosphoglyceride is synthesized by a pathway that
utilizes dietary choline. CDP-choline is the activated intermediate in this route.
Sphingolipids are synthesized from ceramide, which is formed by the acylation of sphingosine. Gangliosides are
sphingolipids that contain an oligosaccharide unit having at least one residue of N-acetylneuraminate or a related sialic
acid. They are synthesized by the step-by-step addition of activated sugars, such as UDP-glucose, to ceramide.
Cholesterol Is Synthesized from Acetyl Coenzyme A in Three Stages
Cholesterol is a steroid component of eukaryotic membranes and a precursor of steroid hormones. The committed step in
its synthesis is the formation of mevalonate from 3-hydroxy-3-methylglutaryl CoA (derived from acetyl CoA and
acetoacetyl CoA). Mevalonate is converted into isopentenyl pyrophosphate (C
5
), which condenses with its isomer,
dimethylallyl pyrophosphate (C
5
), to form geranyl pyrophosphate (C
10
). The addition of a second molecule of
isopentenyl pyrophosphate yields farnesyl pyrophosphate (C
15
), which condenses with itself to form squalene (C
30
).
This intermediate cyclizes to lanosterol (C
30
), which is modified to yield cholesterol (C
27
).
The Complex Regulation of Cholesterol Biosynthesis Takes Place at Several Levels
In the liver, cholesterol synthesis is regulated by changes in the amount and activity of 3-hydroxy-3-methylglutaryl CoA
reductase. Transcription of the gene, translation of the mRNA, and degradation of the enzyme are stringently controlled.
In addition, the activity of the reductase is regulated by phosphorylation.
Triacylglycerols exported by the intestine are carried by chylomicrons and then hydrolyzed by lipases lining the
capillaries of target tissues. Cholesterol and other lipids in excess of those needed by the liver are exported in the form of
very low density lipoprotein. After delivering its content of triacylglycerols to adipose tissue and other peripheral tissue,
VLDL is converted into intermediate-density lipoprotein and then into low-density lipoprotein. IDL and LDL carry
cholesteryl esters, primarily cholesteryl linoleate. Liver and peripheral tissue cells take up LDL by receptor-mediated
endocytosis. The LDL receptor, a protein spanning the plasma membrane of the target cell, binds LDL and mediates its
entry into the cell. Absence of the LDL receptor in the homozygous form of familial hypercholesterolemia leads to a
markedly elevated plasma level of LDL-cholesterol, the deposition of cholesterol on blood-vessel walls, and heart
attacks in childhood. Apolipoprotein B, a very large protein, is a key structural component of chylomicrons, VLDL, and
LDL.
Important Derivatives of Cholesterol Include Bile Salts and Steroid Hormones
In addition to bile salts, which facilitate the digestion of lipids, five major classes of steroid hormones are derived from
cholesterol: progestagens, glucocorticoids, mineralocorticoids, androgens, and estrogens. Hydroxylations by P450
monooxygenases that use NADPH and O
2
play an important role in the synthesis of steroid hormones and bile salts from
cholesterol. P450 enzymes, a large superfamily, also participate in the detoxification of drugs and other foreign
substances.
Pregnenolone (C
21
) is an essential intermediate in the synthesis of steroids. This steroid is formed by scission of the side
chain of cholesterol. Progesterone (C
21
), synthesized from pregnenolone, is the precursor of cortisol and aldosterone.
Hydroxylation of progesterone and cleavage of its side chain yields androstenedione, an androgen (C
19
). Estrogens (C
18
)
are synthesized from androgens by the loss of an angular methyl group and the formation of an aromatic A ring. Vitamin
D, which is important in the control of calcium and phosphorus metabolism, is formed from a derivative of cholesterol
by the action of light.
In addition to cholesterol and its derivatives, a remarkable array of biomolecules are synthesized from isopentenyl
pyrophosphate, the basic five-carbon building block. The hydrocarbon side chains of vitamin K
2
, coenzyme Q
10
, and
chlorophyll are extended chains constructed from this activated C
5
unit. Prenyl groups that are derived from this
activated intermediate target many proteins to membranes.
Key Terms
phosphatidate
triacylglycerol
phospholipid
cytidine diphosphodiacylglycerol (CDP-diacylglycerol)
glyceryl ether phospholipid
sphingolipid
ceramide (N-acyl sphingosine)
cerebroside
ganglioside
cholesterol
mevalonate
3-hydroxy-3-methylglutaryl CoA reductase (HMG-CoA reductase)
3-isopentenyl pyrophosphate
sterol regulatory element binding protein (SREBP)
lipoprotein particles
low-density lipoprotein (LDL)
high-density lipoprotein (HDL)

receptor-mediated endocytosis
bile salts
steroid hormones
cytochrome P450 monooxygenase
pregnenolone
III. Synthesizing the Molecules of Life 26. The Biosynthesis of Membrane Lipids and Steroids
Problems
1.
Making fat. Write a balanced equation for the synthesis of a triacylglycerol, starting from glycerol and fatty acids.
See answer
2.
Making phospholipid. Write a balanced equation for the synthesis of phosphatidyl serine by the de novo pathway,
starting from serine, glycerol, and fatty acids.
See answer
3.
Activated donors. What is the activated reactant in each of the following biosyntheses?
(a) Phosphatidyl serine from serine
(b) Phosphatidyl ethanolamine from ethanolamine
(c) Ceramide from sphingosine
(d) Sphingomyelin from ceramide
(e) Cerebroside from ceramide
(f) Ganglioside G
M
1
from ganglioside G
M
2
(g) Farnesyl pyrophosphate from geranyl pyrophosphate
See answer
4.
Telltale labels. What is the distribution of isotopic labeling in cholesterol synthesized from each of the following
precursors?
(a) Mevalonate labeled with
14
C in its carboxyl carbon atom
(b) Malonyl CoA labeled with
14
C in its carboxyl carbon atom
See answer
5.
Familial hypercholesterolemia. Several classes of LDL-receptor mutations have been identified as causes of this
disease. Suppose that you have been given cells from patients with different mutations, an antibody specific for the
LDL receptor that can be seen with an electron microscope, and access to an electron microscope. What differences
in antibody distribution might you expect to find in the cells from different patients?
See answer
6.
RNA editing. A shortened version (apo B-48) of apolipoprotein B is formed by the intestine, whereas the full-length
protein (apo B-100) is synthesized by the liver. A glutamine codon (CAA) is changed into a stop codon. Propose a
simple mechanism for this change.
See answer
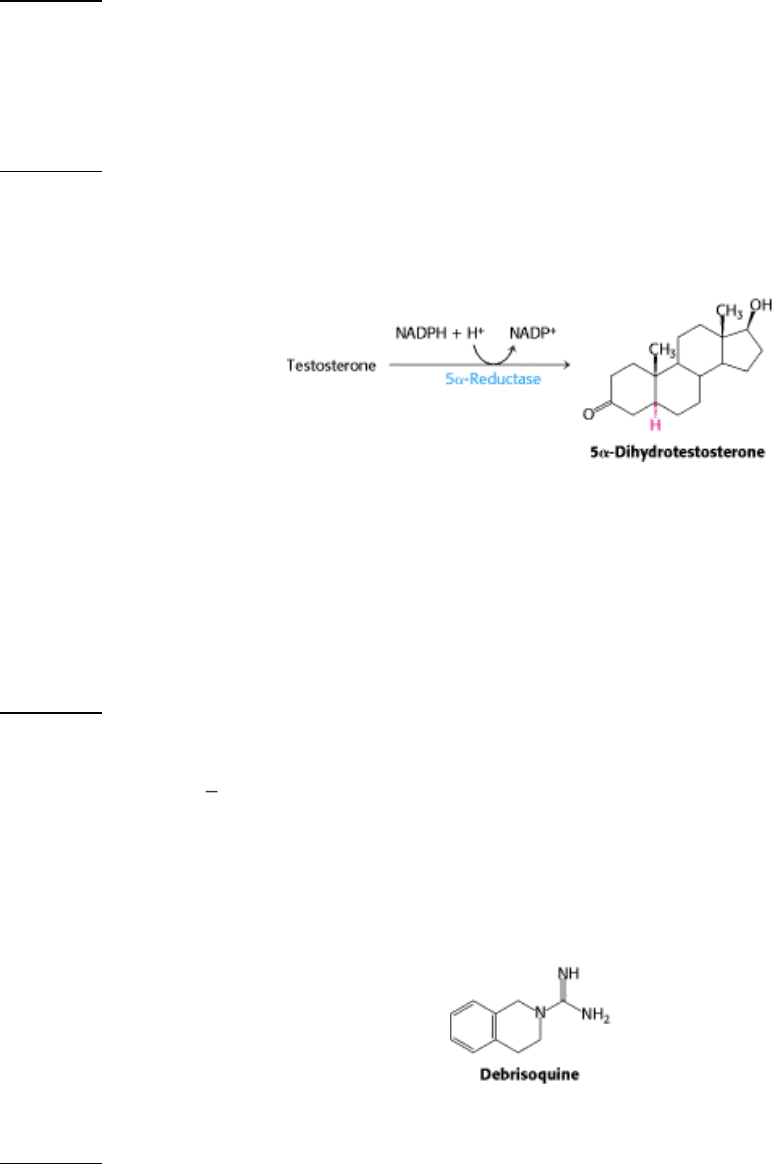
7.
Inspiration for drug design. Some actions of androgens are mediated by dihydrotestosterone, which is formed by the
reduction of testosterone. This finishing touch is catalyzed by an NADPH-dependent 5α-reductase.
Chromosomal XY males with a genetic deficiency of this reductase are born with a male internal urogenital tract but
predominantly female external genitalia. These people are usually reared as girls. At puberty, they masculinize
because the testosterone level rises. The testes of these reductase-deficient men are normal, whereas their prostate
glands remain small. How might this information be used to design a drug to treat benign prostatic hypertrophy, a
common consequence of the normal aging process in men? A majority of men older than age 55 have some degree
of prostatic enlargement, which often leads to urinary obstruction.
See answer
8.
Drug idiosyncrasies. Debrisoquine, a β-adrenergic blocking agent, has been used to treat hypertension. The optimal
dose varies greatly (20 400 mg daily) in a population of patients. The urine of most patients taking the drug contains
a high level of 4-hydroxydebrisoquine. However, those most sensitive to the drug (about 8% of the group studied)
excrete debrisoquine and very little of the 4-hydroxy derivative. Propose a molecular basis for this drug
idiosyncrasy. Why should caution be exercised in giving other drugs to patients who are very sensitive to
debrisoquine?
See answer
9.
Removal of odorants. Many odorant molecules are highly hydrophobic and concentrate within the olfactory
epithelium. They would give a persistent signal independent of their concentration in the environment if they were
not rapidly modified. Propose a mechanism for converting hydrophobic odorants into water-soluble derivatives that
can be rapidly eliminated.
See answer
10.
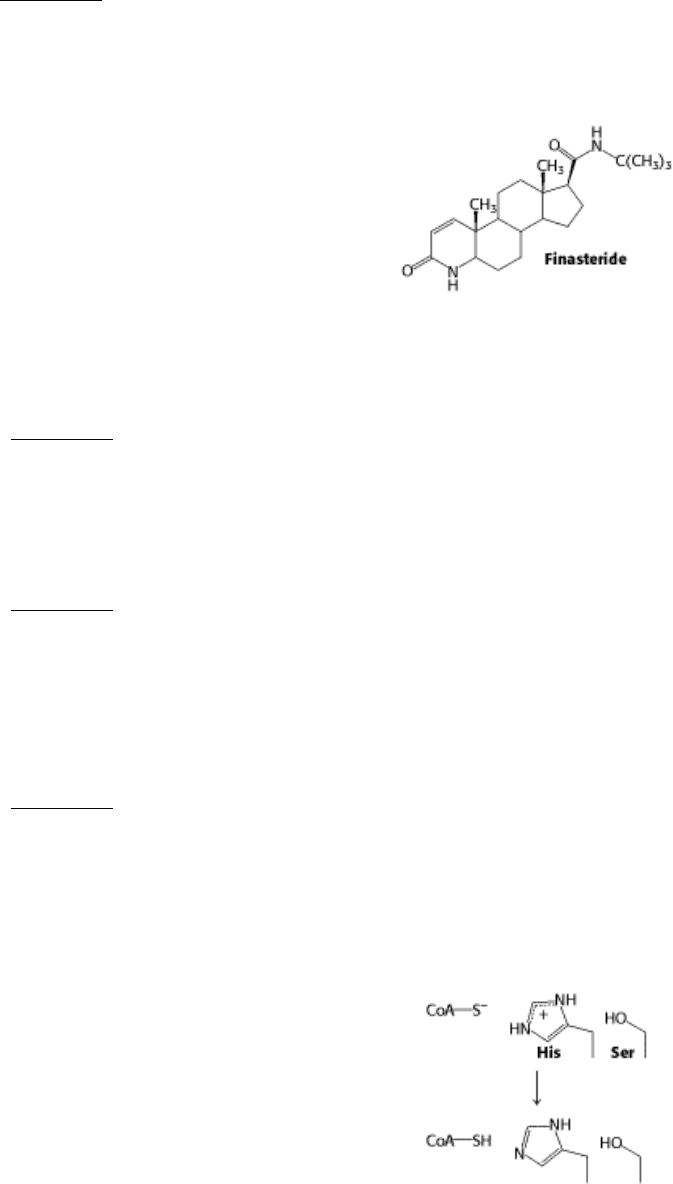
Development difficulties. Propecia (finasteride) is a synthetic steroid that functions as a competitive and specific
inhibitor of 5α-reductase, the enzyme responsible for the synthesis of dihydrotestosterone from testosterone.
It is now widely used to retard the development of male pattern hair loss. Pregnant women are advised to avoid
handling this drug. Why is it vitally important that pregnant women avoid contact with Propecia?
See answer
11.
Life-style consequences. Human beings and the plant Arabidopsis evolved from the same distant ancestor
possessing a small number of cytochrome P450 genes. Human beings have approximately 50 such genes, whereas
Arabidopsis has more than 250 of them. Propose a role for the large number of P450 isozymes in plants.
See answer
12.
Personalized medicine. The cytochrome P450 system metabolizes many medicinally useful drugs. Although all
human beings have the same number of P450 genes, individual polymorphisms exist that alter the specificity and
efficiency of the proteins encoded by the genes. How could knowledge of individual polymorphisms be useful
clinically?
See answer
Mechanism Problem
13.
An interfering phosphate. In the course of the overall reaction catalyzed by HMG-CoA reductase, a histidine
residue protonates a coenzyme A thiolate, CoAS
-
, generated in a previous step.
The nearby serine residue can be phosphorylated by AMPdependent kinase, which results in a loss of activity.
Propose an explanation for why phosphorylation of the serine residue inhibits enzyme activity.