Caballero B. (ed.) Encyclopaedia of Food Science, Food Technology and Nutrition. Ten-Volume Set
Подождите немного. Документ загружается.


glucose administered intravenously. This ability of
the gut to stimulate insulin secretion has been termed
the enteroinsular axis. Gut peptides potentiate insulin
secretin via both neural and endocrine pathways but,
in humans, the endocrine role of the gut hormones
(collectively termed incretins) is considered to be
quantitatively the more important. Although most
members of the secretion family of gut hormones
have the ability to stimulate insulin in pharmaco-
logical doses, only GIP and GLP-1 are both secreted
in response to glucose ingestion and stimulate insulin
secretion at physiological levels. Together they
account for the entire endocrine arm of the entero-
insular axis.
0016 GIP and GLP-1 are both potent stimulators of
glucose-induced insulin secretion. On a molar basis,
GLP-1 is the stronger stimulator, but as it circulates
at lower concentrations than GIP, the contribution
of the two hormones to the enteroinsular axis are
approximately equivalent. These hormones only
stimulate insulin secretion in the presence of hyper-
glycemia. This can be looked upon as providing a
safeguard against inappropriate insulin secretion, as
both hormones are also stimulated by oral fat, when
insulin secretion would be undesirable unless its
hypoglycemic properties were counteracted by simul-
taneous ingestion of carbohydrate.
0017 The neural arm of the enteroinsular axis is more
poorly defined. Both CCK- and VIP-containing
neurons have been implicated as neurotransmitters
stimulating insulin secretion. The smaller forms of
CCK (CCK-4 and CCK-8) also appear to be biologic-
ally active, although their precise role in insulin
stimulation is not yet fully known.
0018 Consistent with the anabolic role that gut hor-
mones play in carbohydrate metabolism, by stimulat-
ing insulin secretion, is a parallel anabolic role in lipid
metabolism. GIP has been shown to have an anabolic
effect on lipid metabolism, stimulating lipoprotein
lipase and enhancing postprandial triglyceride clear-
ance from the plasma. GIP and GLP-1 have also been
shown to stimulate de novo fatty acid synthesis in
adipose tissue. These two GI hormones, which po-
tentiate the secretion of insulin, also exhibit insulin-
like actions systemically and provide a direct link
between the absorption and subsequent metabolic
fate of ingested nutrients.
Appetite Regulation
0019 The regulation of hunger and satiety is a complex
process, involving the interaction of both physio-
logical and psychological factors. There is good ex-
perimental evidence for the involvement of several of
the gut hormones in the short-term regulation of
appetite, although the precise mechanisms whereby
they exert their effects in human appetite control are
less clear. CCK and GLP-1 have been found to medi-
ate satiety in both animal and human studies. The
inhibitory effects of CCK on food intake are stimu-
lated by fat ingestion, and appear to be mediated by a
local gastrointestinal action, although CCK is par-
tially dependent upon an intact vagal nerve supply
in order to exert its satiety effects. Peripheral adminis-
tration of GLP-1 has been shown to suppress appetite
in most, if not all human studies; circulating GLP-1
can theoretically pass the blood–brain barrier to act
centrally. Neuropeptide Y is distributed in the central
nervous system as well as within the gut. It acts in the
brain upon appetite centers in the hypothalamus to
stimulate food intake, and it may also be implicated
in the regulation of leptin, the peptide secreted from
adipose tissue which is involved with the longer-term
mediation of satiety.
Regulation of Secretion
0020Ingestion and absorption of food are the major stim-
uli to gut hormone secretion, especially the endocrine
gut hormones. The major nutrient stimuli for these
hormones are listed in Table 3. The type of nutrient
that stimulates the secretion of an individual gut hor-
mone usually relates to its physiological role. GIP and
GLP-1, for example, the potentiators of insulin secre-
tion, are stimulated by carbohydrate, which in itself
stimulates insulin secretion by raising circulating glu-
cose levels. Fat ingestion is a major stimulus to secre-
tin and CCK whose primary function is concerned
with stimulating pancreatic and biliary secretion,
thus aiding the digestion of fat. GIP secretion is also
stimulated by fat absorption, consistent with the role
of GIP in aiding the clearance of circulating lipids
after a meal, and aiding their uptake into adipose
tissue. The secretory response of a gut hormone to
nutrients is a very precise one. For example, carbohy-
drate-induced GIP secretion is stimulated specifically
by the absorption of actively transported sugars such
as glucose and galactose. The mere presence of nutri-
ents in the gut lumen is insufficient to stimulate GIP
secretion, and sugars such as fructose, whose absorp-
tion does not require active transport mechanism, do
not stimulate GIP secretion. In contrast, GLP-1 secre-
tion is not dependent upon the absorption of nutri-
ents, a property which is consistent with its role as a
regulator of gastric emptying and the smooth delivery
of nutrients to the upper part of the gut. Chronic
dietary changes can also affect gut hormone secre-
tion; a high-fat diet, for example, has been shown to
increase GIP secretion in response to both carbohy-
drate and fat ingestion.
HORMONES/Gut Hormones 3149

0021 Interactions between the various gut hormones
play an important part in the regulation of gut
hormone secretion, particularly in an inhibitory
capacity (Table 3). Thus somatostatin is a physio-
logical paracrine inhibitor of gastrin secretion and
probably plays an endocrine role in the regulation of
other gut hormones, e.g., secretin and neurotensin.
GRP is so named because of its role in the stimulation
of gastric secretion. GRP infusion will also cause
increased plasma concentrations of many other GI
hormones, although the physiological relevance of
this is unclear.
0022 Less is known about the regulation of gut neuro-
peptide secretion. These peptides are synthesized by
intramural neurons and released from nerve terminals
by physiologically appropriate stimuli (e.g., chemical
or mechanically induced reflexes). Vagal stimulation
has been implicated in the release of several gut
neurotransmitter peptides, e.g., VIP and GRP, but its
regulatory role has yet to be fully defined.
See also: Diabetes Mellitus: Etiology; Hormones:
Pancreatic Hormones
Further Reading
Holst JJ, Fahenkrug J, Stadil F and Rehfeld JF (1996) Gas-
trointestinal endocrinology. Scandinavian Journal of
Gastroenterology 216: 27–38.
Morgan LM (1998) The role of gastrointestinal hormones
in carbohydrate and lipid metabolism and homeostasis:
effects of gastric inhibitory polypeptide and glucagon-
like peptide-1. Biochemical Society Transactions 26:
216–222.
Rehfeld JF (1998) The new biology of the gastrointestinal
hormones. Physiological Reviews 78: 1087–1103.
Schultz H, Makhlouf GM and Rauner BB (eds) (1989) The
gastrointestinal system. Neural and endocrine biology.
In: Handbook of Physiology, vol. II, Section 6. Bethesda:
American Physiological Society.
Wilding JPH, Ghatei MA and Bloom SR (1995) Hormones
of the gastrointestinal tract. In: DeGroot LJ (ed.)
Endocrinology, pp. 2870–2894. Philadelpia, London:
Saunders.
Pancreatic Hormones
N H McClenaghan and P R Flatt, University of Ulster,
Coleraine, UK
Copyright 2003, Elsevier Science Ltd. All Rights Reserved.
Pancreatic Secretions
0001The pancreas is a secretory gland playing a key
role in the metabolism of carbohydrates, proteins,
and lipids. Cells of the exocrine pancreas (acini)
secrete pancreatic juice into the duodenum, which
contains enzymes for the digestion of major food
tbl0003 Table 3 Nutritional and other regulation of endocrine gastraintestinal hormone secretion
Hormone/peptide Majornutritional stimulants Other modulators of secretion
Secretin Fat. Exit of meal from stomach causing
duodenal acidification
Stimulation: bile salts
Inhibition: somatostatin
Gastric-inhibitory peptide (GIP) Fat, carbohydrate. Absorption from upper
small intestine
Inhibition: insulin
Glucagon-like peptides Carbohydrate, fat. Presence of food in upper
small intestine
Stimulation: gastrin-releasing peptide (GRP)
Inhibition: somatostatin
Gastrin Amino acids and protein in stomach lumen;
meal-induced gastric distension
Stimulation: GRP, acetylcholine,
norepinephrine (noradrenaline)
Inhibition: acid (pH<3), GIP, secretin, GLP-1,
somatostatin
Chdecystokinin Fat (hydrolysis of triglyceride), phenylalanine
and tryptophan
Stimulation: GRP
Inhibition: trypsin, bile salts, somatostatin
Pancreatic polypeptide Ingestion of meal, especially carbohydrate Stimulation: GRP, motilin, vagal stimulation
Inhibition: somatostatin
Peptide YY Ingestion of meal, especially fat Inhibition: GLP-1
Somatostatin Ingestion of meal, especially fat and protein.
Meal-induced lowering of gastric pH
Stimulation: GIP, CCK, secretin
Inhibition: opioid peptides
Neurotensin Fat ingestion Stimulation: GRP
Inhibition: somatostatin
Motilin Stimulated by both oral and intravenous fat
(inhibited by glucose, amino acids)
Stimulation: duodenal acidification
Inhibition: secretin, GRP, somatostatin
GLP-1, glucagon-like peptide 1.
3150 HORMONES/Pancreatic Hormones

components. The endocrine pancreas synthesizes
and secretes peptide hormones into the blood,
thereby controlling nutrient homeostasis by deter-
mining the metabolic fate of absorbed glucose,
amino acids, and fatty acids, and their mobilization
in times of need from body stores. The following
sections focus specifically on the endocrine pancreas,
and the synthesis, secretion, and action of pancreatic
hormones.
Organization of the Endocrine Pancreas
0002 The endocrine function of the pancreas resides within
the islet of Langerhans, which constitute 1–3% of
total pancreatic mass. The normal adult pancreas
contains about 1 10
6
pancreatic islets scattered
throughout the body of this elongated organ. Islets
are spherical structures less than the size of a pinhead,
each comprising a cluster of 2000–6000 individual
islet cells surrounded by a connective tissue capsule.
The diameter of the islet ranges from 20 to 300 mm,
depending on cellular composition.
Pancreatic Islet Cells
0003 Pancreatic islets consist of four major types of secre-
tory cell producing insulin (b cells), glucagon (a
2
cells), somatostatin (a
1
cells; also referred to as d
cells) and pancreatic polypeptide (PP cells) (Table 1).
These are sometimes also referred to as B, A, and D
cells, respectively. The products of several other
minor secretory cell types located in the islets remain
unknown. Islets also contain small amounts of add-
itional peptides colocalized with classical islet hor-
mones or present in nerve fibers. These agents have
various effects on insulin secretion, as indicated in
Table 1.
0004 Each islet is composed of 60–70% b cells located
primarily as a central core surrounded by an outer
mantle of a
2
cells (5–25%), a
1
cells (5–10%) and PP
cells (15–20%). The proportion of a
2
and PP cells
varies considerably, depending on islet embryonic
origin and location in the pancreas. Islets distributed
the tail and body of the pancreas are generally rich in
a
2
cells and contain relatively few PP cells, whereas
the converse applies for islets located the ventral head
region of the pancreas.
Islet Vasculature
0005 Islets are well vascularized and receive a considerable
flow of blood from afferent arterioles arising from the
splenic and pancreaticoduodenal arteries. Within the
islet, the afferent vessels appear to perforate the cellu-
lar mantle, supplying a dense network of capillaries
that traverse the b cell core, before draining into a
small number of venules that ultimately feed into the
pancreatic vein. The vasculature of individual islets is
believed to constitute a tiny intraislet ‘portal’ system,
with blood flowing progressively past b, a
2
, and a
1
cells. This arrangement provides effective delivery to
islet cells of bloodborne stimuli, such as glucose, and
ensures rapid circulation of the secreted islet hor-
mones to peripheral sites of action. There is also
recent evidence that elevation of blood glucose spe-
cifically increases islet blood flow, via a cholinergic
mechanism.
Islet Innervation
0006Islets receive an abundant supply of cholinergic, adre-
nergic, and peptidergic (nonadrenergic, noncholiner-
gic (NANC)) nerves that innervate islet cells and the
surrounding capillaries. In addition to the classical
neurotransmitters – acetylcholine and noradrenaline
– the islets contain a number of neuropeptides that
exert stimulatory or inhibitory effects on islet cell
secretions (Tables 1 and 2). Parasympathetic innerv-
ation is activated by feeding, whereas sympathetic
nerves are activated in situations of need such as
starvation or fright.
Islet Cell Interactions
0007Glucagon stimulates the secretion of insulin and
somatostatin, whereas both insulin and somatostatin
tbl0001Table 1 Hormones, peptides colocalized in hormone-
producing cells and neurotransmitters in islets of Langerhans
Cell type Effect oninsulin
secretion
Hormones
Insulin b cells #
Glucagon a
2
cells "
Somatostatin a
1
cells #
Pancreatic polypeptide PP cells #
Peptides colocalized
DBI a
2
, a
1
cells #
IAPP b cells #
Pancreastatin a
2
, b, a
1
cells #
TRH a
2
, b cells "
Neurotransmitters
Acetylcholine Nerve fibers "
Noradrenaline Nerve fibers #
CCK Nerve fibers "
Galanin Nerve fibers #
GRP Nerve fibers "
VIP Nerve fibers "
", stimulatory effect; #, inhibitory effect; CCK, cholecystokinin;
DBI, diazepam-binding inhibitor; GRP, gastrin-releasing polypeptide;
IAPP, islet amyloid polypeptide; TRH, thyrotropin-releasing hormone;
VIP, vasoactive intestinal polypeptide.
HORMONES/Pancreatic Hormones 3151
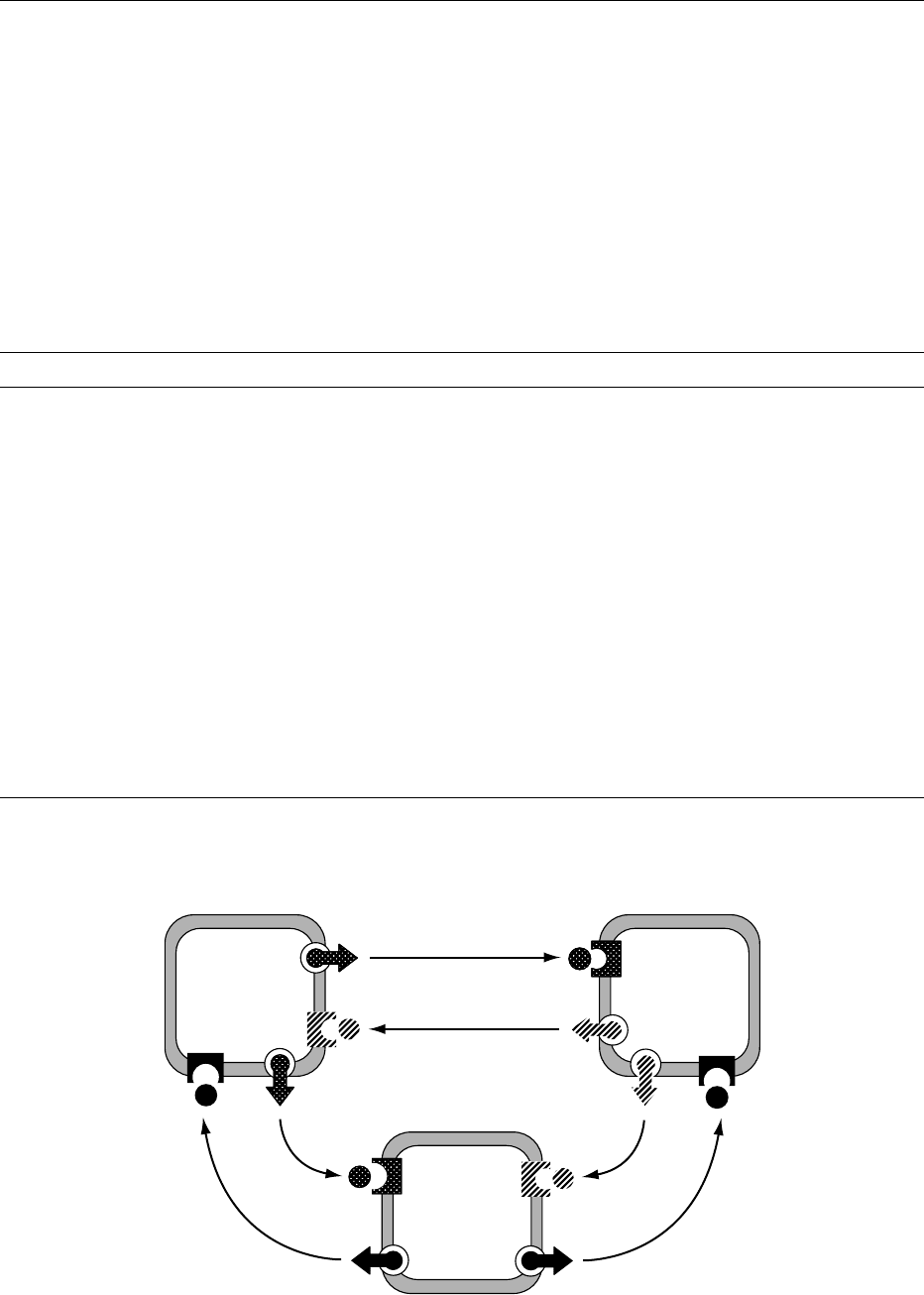
are general inhibitors of islet hormone secretions
(Table 2). This provides the basis for the possible
involvement cellular interactions in the integration
of islet secretory function (Figure 1). Such inter-
actions may be restricted to adjacent cells within the
islet, termed paracrine interactions, or may be more
widely mediated through the intraislet portal system,
which delivers insulin from b cells downstream to
a
2
cells, followed by onward passage of both insulin
and secreted glucagon to the somatostatin-containing
a
1
cells.
Hormone Biosynthesis and Secretion
0008Biosynthesis and secretion of islet hormones occur
through a similar sequence, involving the production
of a precursor polypeptide in the rough endoplasmic
reticulum followed by posttranslational enzymatic
conversion to the active hormone. The active hor-
mone is stored in secretory granules before ultimate
release from the stimulated cell by a regulated process
termed ‘exocytosis.’ This involves movement, aided
by microtubules and microfilaments of the granule to
α
2
cell
(glucagon)
β cell
(insulin)
α
1
cell
(somatostatin)
(−)
(−) (−)
(−)
(+)
(+)
fig0001 Figure 1 Hormonal interactions of major islet cells. Blood flows from b to a
2
to a
1
cells draining into the portal vein.
tbl0002 Table 2 Control of insulin, glucagon, somatostatin, and pancreatic polypeptide secretion
Agent Insulin Glucagon Somatostatin Pancreatic polypeptide
Nutrients
Glucose "# " #
Amino acids "" " "
Fatty acids "# " "
Pancreatic hormones
Insulin ## # "
Glucagon " ? " –
Somatostatin ## # #
Pancreatic polypeptide # – # ?
Intestinal hormones
CCK
a
"" " "
Gastrin "" " –
GIP "" " "
GLP-1[7-36]-amide "# " ?
Secretin "# " –
VIP
a
"" ? "
Neurotransmitters
Acetylcholine "" # "
a
2
-Adrenergic agonists ## # #
b-Adrenergic agonists "" " "
a
Also present in nerve fibers innervating islet cells.
", stimulatory effect; #, inhibitory effect; – no effect; ?, not established; CCK, cholecystokinin; GIP, gastric inhibitory polypeptide; GLP-1[7-36]-amide,
glucagon-like peptide 1[7-36]amide (sometimes referred to as ‘insulinoptropin’); VIP, vasoactive intestinal polypeptide.
3152 HORMONES/Pancreatic Hormones

the periphery, followed by fusion of the granule mem-
brane with the plasma membrane and liberation of
the granule contents into the blood.
0009 More specific details of events concerning biosyn-
thesis and secretion are best known for insulin. In the
b cell, transcription of messenger ribonucleic acid
(mRNA) gives rise to production of preproinsulin,
an 11.5-kDa polypeptide. This is rapidly cleaved
within 1 min by proteolytic enzymes to proinsulin
(86 amino acid residues, 9 kDa) which is transported
by microvesicles to the Golgi apparatus. Proinsulin is
then packaged into secretory granules, where it is
converted by proteases to equimolar amounts of
insulin (51 residues, 6 kDa) and proinsulin C-peptide.
A number of second messengers are involved in
coupling stimulus recognition to the secretion of
insulin by exocytosis. These include cytoplasmic
calcium ion concentration ([Ca
2þ
]
i
), cyclic adenosine
monophosphate (cAMP), diacylglycerol (DAG), and
inositol 1,4,5-trisphosphate (IP
3
).
0010 In pancreatic a
2
cells, the initial product of the
proglucagon is a 160-residue polypeptide (18 kDa)
that contains the amino acid sequences of several
biologically active peptides in addition to glucagon.
Posttranslational processing of proglucagon yields
glucagon (29 residues, 3.5 kDa) as the main product,
plus a proglucagon fragment and, possibly, small
amounts of glicentin-related pancreatic peptide
(GRPP). In pancreatic a
1
cells, preprosomatostatin
(116 residues) is similarly processed to prosomato-
statin (92 residues) before conversion to the active,
14-amino-acid form of somatostatin (SRIF-14,
1.5 kDa). In PP cells, pancreatic polypeptide (36 resi-
dues, 4.3 kDa) also appears to be produced from a
larger, 95-amino-acid, prohormone form. In the in-
testine, the posttranslational processing of progluca-
gon and prosomatostatin differs, yielding as major
products glucagon-like peptide-1[7-36]-amide (GLP-
1[7-36]-amide) and the 28-amino-acid form of soma-
tostatin (SRIF-28), respectively.
Regulation of Secretion
0011 A wide range of nutrients, hormones, and neurotrans-
mitters are capable of influencing the secretion of
insulin, glucagon, somatostatin, and pancreatic poly-
peptide (Tables 1 and 2). In the case of insulin, the
major physiological regulator of b cell function is
the prevailing plasma glucose concentration. Other
metabolizable nutrients can mimic the stimulatory
action of glucose, but these other agents basically
act as modulators of the glucose response.
0012 Insulin acting locally within the islet is an influen-
tial inhibitor of glucagon secretion. Glucose inhib-
ition of glucagon secretion is thus attributable in
large part to concomitant stimulation of the b cell
with the release of insulin. g-Aminobutyric acid
(GABA), cosecreted with insulin from the b cell, has
also been suggested to contribute to inhibition of
a
2
cells. The paradoxical ability of amino acids and
food-activated elements of the enteroinsular axis to
stimulate glucagon as well as insulin secretion is often
considered to offer protection from hypoglycemia
that might result from the marked stimulation of
insulin release. An alternative interpretation is that
the stimulation of glucagon release serves to promote
insulin secretion by paracrine interaction within
the islet. Other situations associated with secretion
of glucagon include hypoglycemia, starvation, and
stress. In these circumstances, glucagon serves to
mobilize nutrient fuels from body stores to meet
immediate energy requirements.
Nutrient Regulation of Insulin Secretion
0013Although many factors influence the secretion of
pancreatic hormones, the regulation of insulin secre-
tion is particularly critical for normal metabolism
and maintenance of good health. b cell dysregulation
leading to deficient insulin secretion results in
impaired glucose tolerance, hyperglycemia and type
2 diabetes (also known as noninsulin-dependent dia-
betes mellitus). Hypersecretion of insulin may result
in life-threatening hypoglycemia, as observed in cer-
tain individuals with an islet cell tumor (insulinoma).
0014Nutrients exert direct and indirect effects on insulin
secretion. Thus, in addition to the direct stimulatory
actions of glucose, amino acids, and fatty acids on the
b cell, the ingestion of food and absorption of nutri-
ents trigger neural and hormonal elements of the
enteroinsular axis (Figure 2). Activation of the para-
sympathetic nerve innervating the islets liberates
acetylcholine from nerve terminals close to b cells.
Stimulation of intestinal endocrine cells leads to the
secretion of GLP-1[7–36]-amide, gastric-inhibitory
polypeptide (GIP) and cholecystokinin (CCK). These
agents are powerful insulin secretagogs at raised
glucose concentrations, and they markedly augment
the direct stimulatory actions of glucose and other
nutrients.
0015The mechanisms through which nutrients and
enteroinsular stimuli trigger insulin secretion from b
cells involve elevation of [Ca
2þ
]
i
and sensitization of
the exocytotic process to normal stimulatory action
of [Ca
2þ
]
i
(Figure 3). Metabolizable nutrients, includ-
ing glucose, certain amino acids, and fatty acids, act
by virtue of the ability of the b cell to metabolize these
substances with the generation of adenosine triphos-
phate (ATP). ATP generation results in closure of
ATP-sensitive potassium channels (K
ATP
) channels in
HORMONES/Pancreatic Hormones 3153
the plasma membrane, leading to membrane de-
polarization. This results in the opening of voltage-
dependent calcium channels (VDCCs) allowing Ca
2þ
to enter the b cell down its concentration gradient.
The subsequent increase of [Ca
2þ
]
i
triggers secretion
of insulin by exocytosis. Elevation of [Ca
2þ
]
i
helps to
restore the membrane potential through opening of
Ca
2þ
-sensitive K
þ
channels in the plasma membrane.
Conditions of elevated [Ca
2þ
]
i
increase the sensitivity
of the b cell to the stimulatory actions of phospho-
lipase C and adenylate cyclase. Acetylcholine, CCK,
GLP-1[7-36]-amide and GIP are particularly potent
activators of these enzymes. Generation of IP
3
in-
hibits Ca
2þ
sequestration by the endoplasmic reticu-
lum, whereas protein kinase C (PKC) and cAMP
sensitize the secretory process to the stimulatory
action of Ca
2þ
. cAMP also activates protein kinase
(PK) responsible for phosphorylation of functionally
important proteins.
Physiological Actions
0016 Within the islet, insulin is the principal hormone that
appears to directly modulate the secretory activity of
other islet cell types. Outside the islet, insulin is the
anabolic hormone and driving force behind the regu-
lation of carbohydrate, protein and lipid metabolism
(Figure 4). Glucagon is the main catabolic counter-
regulatory hormone, but the net effect of glucagon is
more dependent on the insulin:glucagon molar ratio
rather than the absolute level of glucagon per se. The
minute-to-minute control of insulin and glucagon
secretion from the islets is therefore of paramount
importance in the overall regulation of nutrient
homeostasis.
0017Insulin and glucagon exert major effects on the
metabolism of carbohydrate, protein and lipid
(Table 3). The principal actions of insulin are to
lower circulating concentrations of glucose, amino
acids and fatty acids, to increase fat storage in adipose
tissue, and to promote the formation of muscle pro-
tein. Insulin achieves this by stimulating glucose and
amino acid uptake and by alteration of the activities
of enzymes in liver, muscle, and adipose tissue. These
enzymes mediate the effects of insulin on glycogen-
esis, glycogenolysis, gluconeogenesis, glycolysis, lipo-
genesis, lipolysis, ketogenesis, and both the synthesis
and degradation of protein. The actions of insulin on
these metabolic pathways are generally opposed by
the action of glucagon. Secretion of this counter-
regulatory hormone in conjunction with a decrease
in the insulin:glucagon molar ratio serves to generate
a catabolic response in times of need, involving mo-
bilization of nutrients from various tissue stores.
0018Somatostatin and PP secreted from the islets are
less significant in physiological terms than the other
two pancreatic hormones. Somatostatin and PP are
also found in endocrine cells of the intestine, and
they are both released into the circulation following
feeding.
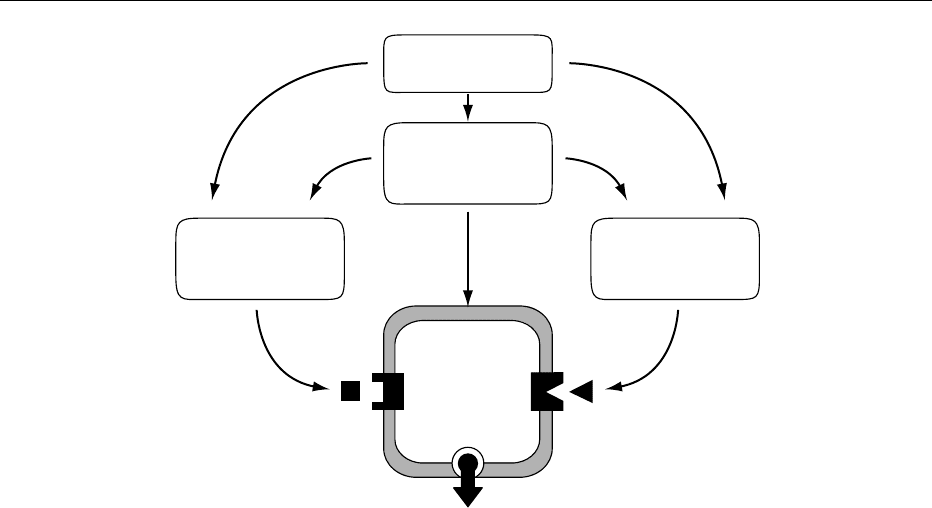
Feeding
Digestion and
absorption of
nutrients
Gastrointestinal
peptide
secretions
Parasympathetic
nervous system
activation
β cell
(insulin)
Insulin exocytosis
(+)
(+)
(+)
(+)
(+) (+)
fig0002 Figure 2 Major neural and hormonal pathways regulating the insulin secretory response to feeding.
3154 HORMONES/Pancreatic Hormones
0019 Somatostatin is a potent inhibitor of secretion
from endocrine (pancreas, intestine, pituitary) and
exocrine cells, partly acting through paracrine
effects, thus contributing to the fine tuning of insulin
and glucagon secretion. Somatostatin also inhibits
the following aspects of gastrointestinal function,
largely as a result of inhibition of gut hormone
secretions: gastric acid secretion; secretion of
bile and pancreatic juice; gall bladder contraction;
intestinal motility; intestinal absorption; splanchnic
blood flow.
0020PP has been shown to exert various actions at
pharmacological concentrations. These include the
following: inhibiting exocrine and endocrine pancre-
atic secretions; gall bladder relaxation; increasing
gastrointestinal motility; inducing satiety. However,
there is presently little convincing evidence for a
physiological role of PP.
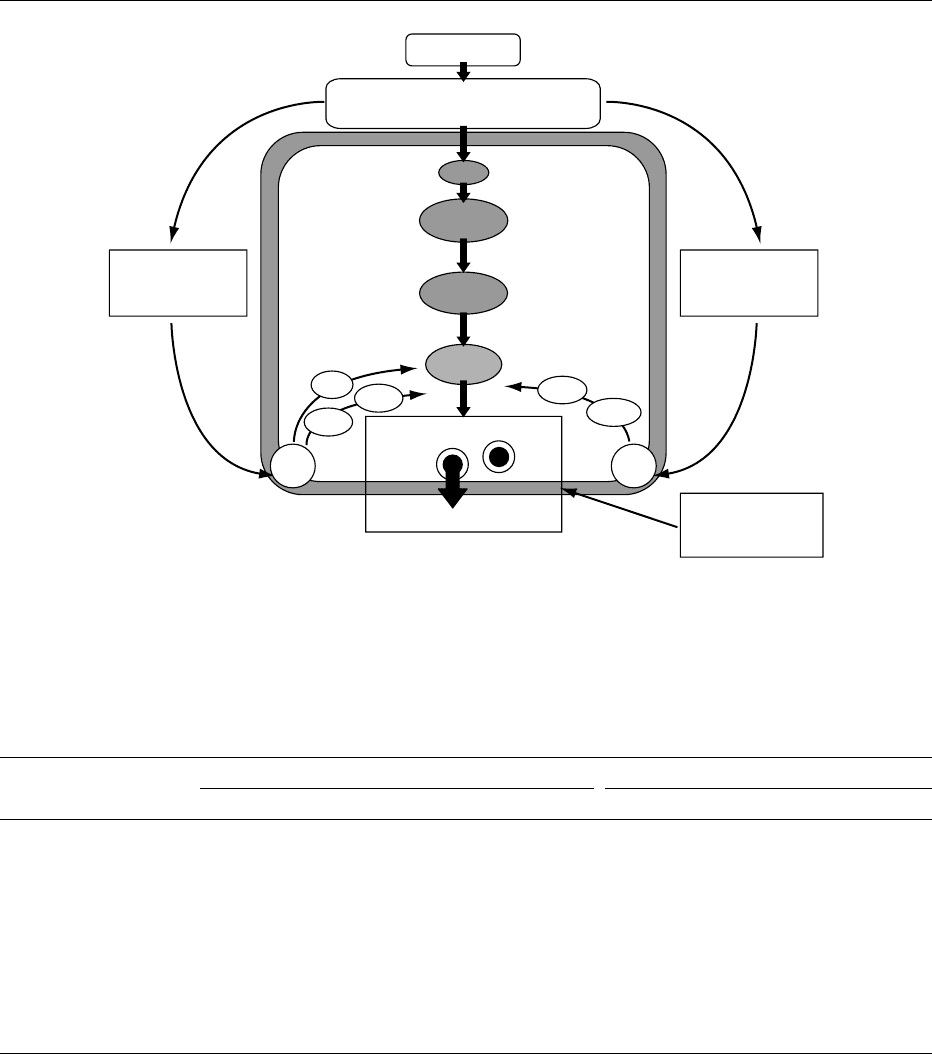
Feeding
Metabolism
Closure
Depolarization
Absorbed nutrients
(glucose, amino acids, fatty acids)
Secretory granule docking
Insulin exocytosis
GIP (+)
GLP-1 (+)
Epinephrine (−)
Norepinephrine (−)
Somatostatin (−)
Acetylcholine (+)
CCK (+)
IP
3
DAG
PKC
[Ca
2
+
]
i
ATP
K
ATP
channel
Ca
2
+
influx
cAMP
PKA
ACPLC
fig0003 Figure 3 Direct and indirect actions of nutrients in the stimulation of insulin secretion from pancreatic b cells. AC, adenylate cyclase;
ATP, adenosine triphosphate; cAMP, cyclic AMP; [Ca
2þ
]
i
], cytoplasmic calcium ion concentration; CCK, cholecystokinin; DAG,
diacylglycerol; GIP, gastric inhibitory polypeptide; GLP-1[7-36]-amide, glucagon-like peptide-1[7-36]-amide; IP
3
, inositol 1,4,5-
triphosphate; K
ATP
channel, ATP-sensitive K
þ
channel; PLC, phospholipase C; PKA, protein kinase A; PKC, protein kinase C.
tbl0003 Table 3 Major metabolic effects of insulin and glucagon on liver, muscle, and adipose tissue
Carbohydrate metabolism Proteinmetabolism Lipidmetabolism
Insulin glucagon Insulin glucagon Insulin glucagon
Blood glucose #"Blood amino acids # – Blood fatty acids #"
Glucose uptake (muscle,
adipose tissue)
" – Protein (muscle) "#Fat storage (adipose tissue) "#
Glycogenesis (liver,
muscle)
"#Amino acid uptake
(muscle, liver)
""Lipogenesis
(liver, adipose tissue)
"#
Glycogenolysis (liver,
muscle)
#"Protein synthesis
(liver, muscle)
" – Lipolysis
(adipose tissue)
#"
Gluconeogenesis (liver) #"Protein degradation
(liver, muscle)
#"Ketogenesis (liver) #"
Glycolysis (liver,
muscle, adipose tissue)
"#
", stimulatory effect; #, inhibitory effect; –, no effect.
HORMONES/Pancreatic Hormones 3155
Mechanisms of Action
0021 Insulin and the other pancreatic hormones are se-
creted into the interstitial fluid; they cross the endo-
thelial barrier and enter the circulation. Effects at the
target cells are initiated by the action and binding
of the hormone to specific cell surface receptors
(Figure 5). The binding of the hormone to these recep-
tors initiates a sequence of postreceptor events ultim-
ately culminating in the various biological actions of
the hormone. Rapid events on the secretion of other
hormones are mediated by changes of [Ca
2þ
]
i
and the
activities of adenylate cyclase and phospholipase C
(see above). Stimulation of glucose uptake by muscle
and adipose tissue results from insulin-induced trans-
location of glucose transporter proteins (GLUT4)
from cytoplasmic vesicles to the plasma membrane.
Classical metabolic effects of insulin and glucagon at
target cells are achieved by activation and suppres-
sion of enzyme activity (so redirecting cell metabol-
ism) or by altering the rate of synthesis of enzymes at
the level of transcription and translation.
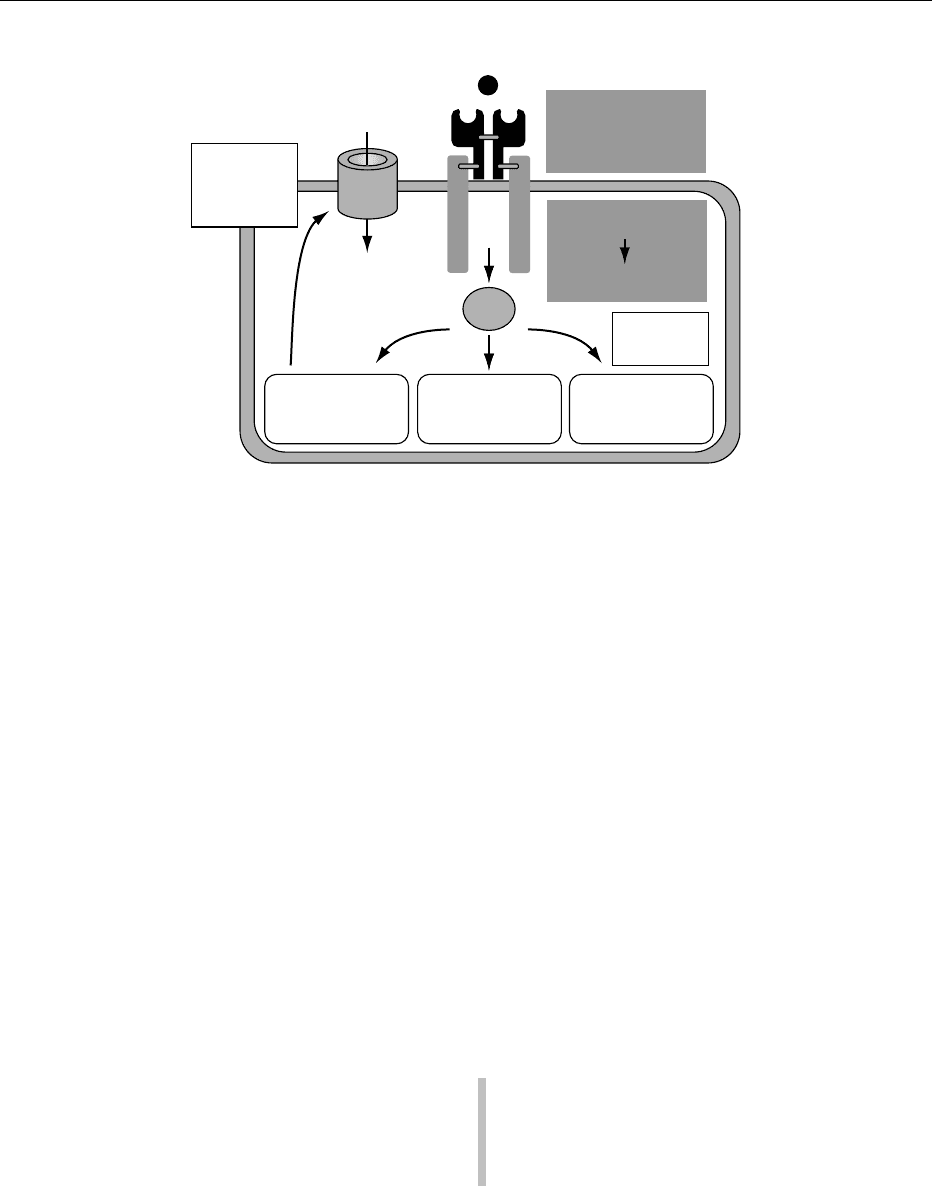
0022 Insulin binding to the external a-subunit of the
glycoprotein insulin receptor leads to a conforma-
tional change in the receptor that stimulates tyrosine
kinase activity. The activated the b-subunit auto-
phosphorylates at tyrosine sites in addition to phos-
phorylating intracellular proteins (Figure 5). Indeed it
is the tyrosine kinase activity that is essential for
insulin signaling. The best characterized postreceptor
signal transduction pathway centers around insulin-
receptor substrate 1 (IRS-1). Rapid tyrosine phos-
phorylation following insulin stimulation results in
noncovalent binding between phosphorylated sites
and specific domains on intracellular protein targets.
This ultimately results in the various important bio-
logical effects of insulin on target tissues (Figure 5).
Following initiation of postreceptor events, the insu-
lin–receptor complex is internalized by the target
cell, and whereas receptors are recycled to the cell
surface, insulin is degraded intracellularly. Elevated
insulin levels, for example in obesity or type 2 dia-
betes, results in insulin receptor ‘downregulation’
through decreasing numbers on the cell surface
and also by reducing tyrosine kinase activity of the
receptor.
Endocrine Pancreas in Diabetes
0023The two major forms of diabetes, type 1 insulin-
dependent diabetes mellitus and type 2 diabetes, are
associated with marked changes in the structure of
the endocrine pancreas. In long-standing type 1 dia-
betes, the insulin deficiency is the result of total auto-
immune destruction of b cells, while the mass of a
2
,
a
1
and PP cells remains virtually unchanged. How-
ever, in type 2 diabetes, the b cell mass is marginally
decreased in association with increases in the a
2
cell
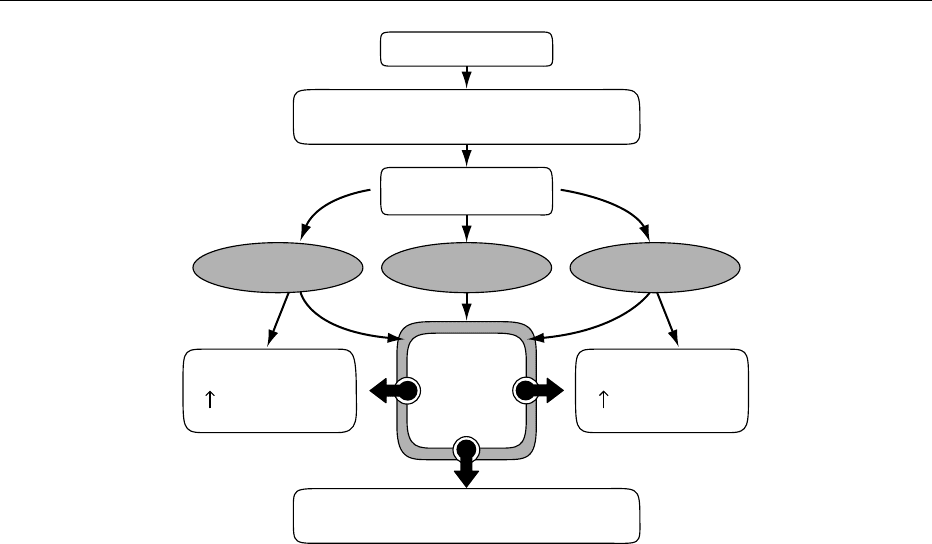
Feeding
Absorbed nutrients
(glucose, amino acids, fatty acids)
Intestine
Muscle
Glycogenesis
Adipose tissue
Lipogenesis
β cell
(insulin)
Anabolic metabolic actions
of insulin
GLP-1[7-36]amide CCK GIP
fig0004 Figure 4 Insulin-mediated anabolic responses to feeding. CCK, cholecystokinin; GIP, gastric inhibitory polypeptide; GLP-1[7-36]-
amide, glucagon-like peptide-1[7-36]-amide.
3156 HORMONES/Pancreatic Hormones
mass and a
2
:b cell ratio. Hyperglucagonemia and
inappropriate a
2
cell function, both owing to a
lack of inhibition of glucagon secretion by insulin,
contribute to glucose intolerance and associated
metabolic disarray generally observed in diabetes.
0024 Type 2 diabetes is classically associated with de-
fective b cell function, normal or moderately raised
insulin concentrations, and insulin resistance. Insulin
secretory responses to glucose and other stimuli are
impaired in magnitude and kinetics. The b cells in
type 2 diabetics are also known to release excessive
amounts of proinsulin rather than insulin. Since
proinsulin cross-reacts with most antisera used to
measure insulin, actual concentrations of insulin in
type 2 diabetes are likely to be much lower than
previously estimated. Proinsulin and derived frag-
ments exhibit only 10–50% of the biological activity
of insulin, and therefore may contribute to the condi-
tion of insulin resistance.
See also: Diabetes Mellitus: Etiology; Chemical
Pathology; Treatment and Management; Problems in
Treatment; Secondary Complications
Further Reading
Alberti KGMM, DeFronzo RA and Zimmet P (eds) (1997)
International Textbook of Diabetes Mellitus, 2nd edn.
Chichester, UK: John Wiley & Sons.
Ashcroft F and Ashcroft SJ (eds) (1992) Insulin. Oxford:
Oxford University Press.
Bailey CJ and Flatt PR (eds) (1990) New Antidiabetic
Drugs. London: Smith-Gordon.
Flatt PR (ed.) (1992) Nutrient Regulation of Insulin Secre-
tion. London: Portland Press.
Flatt PR and Lenzen S (eds) (1994) Frontiers of Insulin
Secretion and Pancreatic B-cell Research. London:
Smith-Gordon.
Frayn KN (1996) Metabolic Regulation: A Human Perspec-
tive. London: Portland Press.
McClenaghan NH and Flatt PR (1999) Physiological and
pharmacological regulation of insulin release: insights
offered through exploitation of insulin secreting cell
lines. Diabetes, Obesity and Metabolism 1: 137–150.
Montague W (1983) Diabetes and the Endocrine Pancreas:
A Biochemical Approach. London: Croom Helm.
Pickup JC and Williams G (eds) (2002) Textbook of Dia-
betes, 3rd edn. Oxford: Blackwell Scientific Publications.
Porte D, Jr and Sherwin RS (eds) (1997) Ellenberg and
Rifkin’s Diabetes Mellitus: Theory and Practice, 5th
edn. Stamford, Connecticut, CT: Appleton & Lange.
Williams G and Pickup JC (eds) (1999) Handbook of Dia-
betes, 2nd edn. Oxford: BlackwellScientific Publications.
Pituitary Hormones
M Wallis, The University of Sussex, Falmer,
Brighton, UK
Copyright 2003, Elsevier Science Ltd. All Rights Reserved.
The Pituitary Gland
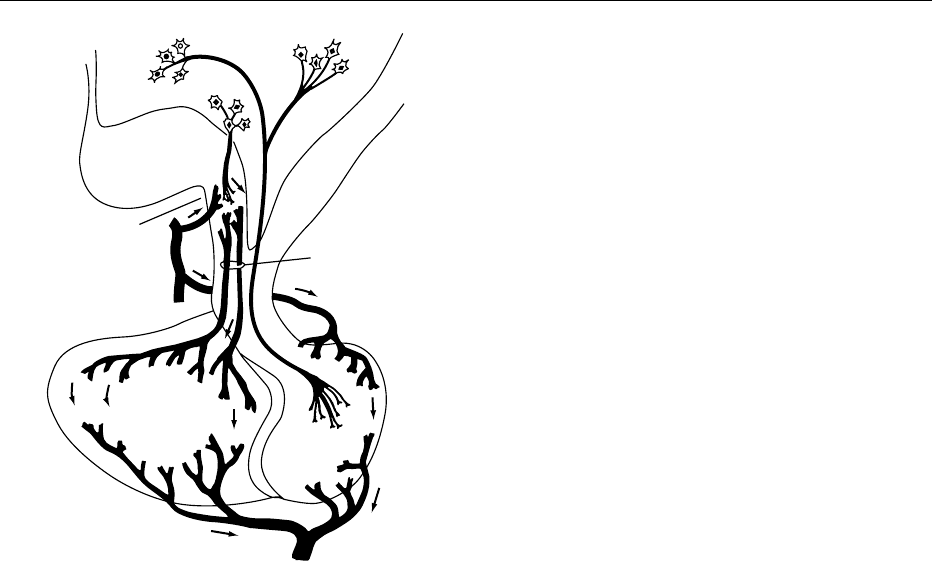
0001In humans the pituitary gland (Figure 1) is a small
organ about the size of a pea which occupies a bony
pocket in the skull (the sella turcica), situated below
Target tissue
Adipose
Liver
Muscle
Insulin
(in bloodstream)
αα
Insulin receptor
·
Glycoprotein
·
400 kDa
·
Divalent
GLUT4
Glucose
transport
Protein, lipid
and glycogen
synthesis
Growth and gene
expression
ββ
IRS-1
Autophosphorylation
of β receptor subunit
Tryosine kinase
activity
Signal
transduction
fig0005 Figure 5 Insulin signalling in target cells. a, a-subunit of insulin receptor; b, b-subunit of insulin receptor; GLUT4, glucose transport
protein; IRS-1, insulin receptor substrate-1.
HORMONES/Pituitary Hormones 3157
the midbrain. It is attached to the floor of the third
ventricle of the brain (the hypothalamus) by a stalk
and this close association between the brain and the
pituitary is of general importance to the functioning
of the latter. The gland comprises two main parts,
with different embryological origins – the neurohy-
pophysis and the adenohypophysis. The neurohypo-
physis (posterior lobe) originates as a downgrowth
from the floor of the brain, the connection with the
brain being retained as a major component of the
pituitary stalk. The bulk of the neurohypophysis is
made up of the swollen nerve terminals of nerve cells
which have their cell bodies in the hypothalamus
(particularly the supraoptic and paraventricular
nuclei), and whose axons pass through the stalk to
the neurohypophysis. The adenohypophysis is ecto-
dermal in origin, being formed during development
from a pouch (Rathke’s pouch) derived from an in-
growing of the cells that will form the roof of the
mouth. The adenohypophysis comprises several
regions, including the anterior lobe (the source of
most adenohypophysial hormones) and the inter-
mediate lobe. The anterior lobe contains about six
different hormone-secreting cell types, secreting 6–
10 different hormones (Table 1); the main function
of several of these is to control the activity of other
endocrine glands. Pituitary growth hormone has a
particularly important role in regulating the nutri-
tional balance of the organism, and this hormone is
emphasized in the second half of this article.
Neurohypophysial Hormones
0002The neurohypophysis in mammals secretes two pep-
tide hormones, oxytocin and vasopressin (antidiuretic
hormone, ADH). Each of these is a nonapeptide,
including a disulfide bridge and an amidated C-
terminus. Oxytocin and vasopressin are very similar,
differing only at two positions. Peptides with slightly
different structures but substantially different bio-
logical actions are found in lower vertebrates; some
invertebrates possess related peptides.
0003In mammals oxytocin functions mainly to regulate
contraction of smooth muscle, especially in mam-
mary gland (leading to milk ejection) and uterus
(stimulating uterine contraction in parturition). Sev-
eral lines of evidence, including recent studies with
transgenic (knockout) mice, suggest that the effects
on the uterus may be of relatively little physiological
importance, although the hormone is widely used
clinically to induce birth. The significance of the
effects on milk ejection is clearly established – if
secretion of oxytocin is prevented (for example, by
anesthetizing a suckling animal) the young are unable
to obtain milk from the mammary gland. The mech-
anism of action of oxytocin involves binding to a G-
protein-linked receptor on the target cells, followed
by activation of intracellular signalling systems
leading to elevation of Ca
2þ
levels and myosin light
chain phosphorylation.
0004The main physiological action of vasopressin in
mammals is regulation of water resorption by the
kidney. A small elevation of blood osmotic pressure
leads to increased vasopressin secretion and increased
water retention at the kidney. Again, the hormone
binds to a specific G-protein-linked receptor on its
target cells, followed by activation of intracellular
signaling systems, including elevation of cyclic adeno-
sine monophosphate (AMP) levels, leading to inser-
tion of water channels into the luminal membrane of
cells of the loop of Henle in the kidney. Vasopressin
has various other effects, including actions on blood
pressure, hepatic metabolism, and adrenocortico-
tropic hormone (ACTH) secretion; at least some of
these are mediated by a different type of receptor
from that found in the kidney.
Optic
chiasma
Median
eminence
Portal
vessels
Artery
Posterior
lobe
Efferent
vein
Anterior
lobe
fig0001 Figure 1 The pituitary gland. Factors secreted at nerve termin-
als in the median eminence of the hypothalamus are carried via
the hypophysial portal vessels to the anterior lobe of the pituitary
gland where they regulate the secretion of the various pituitary
hormones. Precursors of oxytocin and vasopressin are synthe-
sized in neurons in the hypothalamus and transported via axons
(with concomitant processing to the active peptides) to the pos-
terior lobe of the pituitary gland. Modified from Wallis M (1988)
The molecular basis of growth hormone deficiency. Molecular
Aspects of Medicine 10: 429–509.
3158 HORMONES/Pituitary Hormones