Kenneth E. Gonsalves, Craig R. Halberstadt, Cato T. Laurencin, Lakshmi S. Nair. Biomedical Nanostructures
Подождите немного. Документ загружается.

23. Pufe T, et al. Endostatin/collagen XVIIIan inhibitor of angiogenesisis
expressed in cartilage and fibrocartilage. Matrix Biol 2004;23:267276.
24. Brammer RD, Bramhall SR, Eggo MC. Endostatin expression in a pancreatic cell
line is modulated by a TNFalpha-dependent elastase. Br J Cancer 2005;93:
10241028.
25. Brammer RD, Bramhall SR, Eggo MC. Endostatin expression in pancreatic tissue
is modulated by elastase. Br J Cancer 2005;92:8993.
26. Carlson MA and Longaker MT. The fibroblast-populated collagen matrix as a
model of wound healing: a review of the evidence. Wound Repair Regen
2004;12:134147.
27. Lackie JM, Chaabane N, Crocket KV. A critique of the methods used to assess
leucocyte behaviour. Biomed Pharmacother 1987;41:265278.
28. Presta M, Rusnati M, Urbinati C, Sommer A, Ragnotti G. Biologically active
synthetic fragments of human basic fibroblast growth factor (BFGF): Identification
of two Asp-Gly-Arg-containing domains involved in the mitogenic activity of
BFGF in endothelial cells. J Cell Physiol 1991;149:512524.
29. Nagayoshi T, et al. Human nidogen: complete amino acid sequence and structural
domains deduced from cDNAs, and evidence for polymorphism of the gene. DNA
1989;8:581594.
30. Thompson MT, et al. Biochemical functionalization of polymeric cell substrata can
alter mechanical compliance. Biomacromolecules 2006;7:19901995.
31. Engler A, et al. Substrate compliance versus ligand density in cell on gel responses.
Biophys J 2004;86:617628.
32. Pelham RJ Jr and Wang Y. Cell locomotion and focal adhesions are regulated by
substrate flexibility. Proc Natl Acad Sci USA 1997;94:1366113665.
33. Wozniak MA, Desai R, Solski PA, Der CJ, Keely PJ. Rock-generated contractility
regulates breast epithelial cell differentiation in response to the physical properties
of a three-dimensional collagen matrix. J Cell Biol 2003;163:583595.
34. Deroanne CF, Lapiere CM, Nusgens BV. In vitro tubulogenesis of endothelial cells
by relaxation of the coupling extracellular matrixcytoskeleton. Cardiovasc Res
2001;49:647658.
35. Engler AJ, et al. Myotubes differentiate optimally on substrates with tissue-like
stiffness: pathological implications for soft or stiff microenvironments. J Cell Biol
2004;166:877887.
36. Polte TR, Eichler GS, Wang N, Ingber DE. Extracellular matrix controls
myosin light chain phosphorylation and cell contractility through modulation of
cell shape and cytoskeletal prestress. Am J Physiol Cell Physiol 2004;286:
C518528.
37. Wong GA, Tang V, El-Sabeawy F, Weiss RH. BMP-2 inhibits proliferation of
human aortic smooth muscle cells via p21Cip1/Waf1. Am J Physiol Endocrinol
Metab 2003;284:E972979.
38. Damljanovic V, Lagerholm BC, Jacobson K. Fibroblast traction assays on modified
polyacrylamide substrates. Mol Biol Cell 2004;15:160A.
39. Doyle AD and Lee J. Cyclic changes in keratocyte speed and traction stress arise
from Ca2
+
-dependent regulation of cell adhesiveness. J Cell Sci 2005;118:369379.
316
BIOMEDICAL NANOSTRUCTURES
40. Georges PC, Miller WJ, Meaney DF, Sawyer ES, Janmey PA. Matrices with
compliance comparable to that of brain tissue select neuronal over glial growth in
mixed cortical cultures. Biophys J 2006;90:30123018.
41. Woerly S, Doan VD, Sosa N, de Vellis J, Espinosa-Jeffrey A. Prevention of gliotic
scar formation by neurogel allows partial endogenous repair of transected cat spinal
cord. J Neurosci Res 2004;75:262272.
42. Engler AJ, Sen S, Sweeney HL, Discher DE. Matrix elasticity directs stem cell
lineage specification. Cell 2006;126:677689.
43. Abrams GA, Murphy CJ, Wang ZY, Nealey PF, Bjorling DE. Ultrastructural
basement membrane topography of the bladder epithelium. Urol Res 2003;31:
341346.
44. Abrams GA, Bentley E, Nealey PF, Murphy CJ. Electron microscopy of the canine
corneal basement membranes. Cells Tissues Organs 2002;170:251257.
45. Harrison R. On the stereotropism of embryonic cells. Science 1911;34:279.
46. Harrison R. The reaction of the embryonic cells to solid structures. J Exp Zool
1921;17:521.
47. Curtis A and Wilkinson C. Topographical control of cells. Biomaterials 1997;18:
15731583.
48. Washburn NR, Yamada KM, Simon CG Jr, Kennedy SB, Amis EJ. High-
throughput investigation of osteoblast response to polymer crystallinity: influence
of nanometer-scale roughness on proliferation. Biomaterials 2004;25:12151224.
49. Teixeira AI, Abrams GA, Bertics PJ, Murphy CJ, Nealey PF. Epithelial contact
guidance on well-defined micro- and nanostructured substrates. J Cell Sci
2003;116:18811892.
50. Fan YW, et al. Culture of neural cells on silicon wafers with nano-scale surface
topograph. J Neurosci Methods 2002;120:1723.
51. Clark P, Connolly P, Curtis AS, Dow JA, Wilkinson CD. Cell guidance by ultrafine
topography in vitro. J Cell Sci 1991;99: (Pt 1):7377.
52. Dalby MJ, Riehle MO, Johnstone HJ, Affrossman S, Curtis AS. Polymer-demixed
nanotopography: control of fibroblast spreading and proliferation. Tissue Eng
2002;8:10991108.
53. Dalby MJ, Yarwood SJ, Johnstone HJ, Affrossman S, Riehle MO. Fibroblast
signaling events in response to nanotopography: a gene array study. IEEE Trans
Nanobioscience 2002;1:1217.
54. Bystrom B, Virtanen I, Rousselle P, Gullberg D, Pedrosa-Domellof F. Distribution
of laminins in the developing human eye. Invest Ophthalmol Vis Sci 2006;47:
777785.
55. Ziebarth NM, Manns F, Uhlhorn SR, Venkatraman AS, Parel JM. Noncontact
optical measurement of lens capsule thickness in human, monkey, and rabbit
postmortem eyes. Invest Ophthalmol Vis Sci 2005;46:16901697.
56. Futaki S, et al. Molecular basis of constitutive production of basement membrane
components. Gene expression profiles of Engelbreth Holm Swarm tumor and f9
embryonal carcinoma cells. J Biol Chem 2003;278:5069150701.
57. Orkin RW, et al. A murine tumor producing a matrix of basement membrane. J
Exp Med 1977;145:204220.
CELLULAR BEHAVIOR ON BASEMENT 317
58. Kleinman HK, et al. Basement membrane complexes with biological activity.
Biochemistry 1986;25:312318.
59. Kubota Y, Kleinman HK, Martin GR, Lawley TJ. Role of laminin and basement
membrane in the morphological differentiation of human endothelial cells into
capillary-like structures. J Cell Biol 1988;107:15891598.
60. Dalby MJ, Riehle MO, Sutherland DS, Agheli H, Curtis AS. Changes in fibroblast
morphology in response to nano-columns produced by colloidal lithography.
Biomaterials 25:2004;54155422.
61. Price RL, Ellison K, Haberstroh KM, Webster TJ. Nanometer surface roughness
increases select osteoblast adhesion on carbon nanofiber compacts. J Biomed Mater
Res A 2004;70:129138.
62. Stevens MM and George JH. Exploring and engineering the cell surface interface.
Science 2005;310:11351138.
63. Ahmed I, et al. Three-dimensional nanofibrillar surfaces covalently modified with
tenascin-c-derived peptides enhance neuronal growth in vitro. J Biomed Mater Res
A 2006;76:851860.
64. Nur EKA, Ahmed I, Kamal J, Schindler M, Meiners S. Three dimensional
nanofibrillar surfaces induce activation of Rac. Biochem Biophys Res Commun
2005;331:428434.
65. Brunette DM. Spreading and orientation of epithelial cells on grooved substrata.
Exp Cell Res 1986;167:203217.
66. Chehroudi B, Gould TR, Brunette DM. Titanium-coated micromachined grooves
of different dimensions affect epithelial and connective-tissue cells differently in vivo.
J Biomed Mater Res 1990;24:12031219.
67. Chehroudi B, McDonnell D, Brunette DM. The effects of micromachined surfaces
on formation of bonelike tissue on subcutaneous implants as assessed by
radiography and computer image processing. J Biomed Mater Res 1997;34:
279290.
68. Dalby MJ, Riehle MO, Yarwood SJ, Wilkinson CD, Curtis AS. Nucleus alignment
and cell signaling in fibroblasts: response to a micro-grooved topography. Exp Cell
Res 2003;284:274282.
69. den Braber ET, et al. Effect of parallel surface microgrooves and surface energy on
cell growth. J Biomed Mater Res 1995;29:511518.
70. Kim SO, et al. Epitaxial self-assembly of block copolymers on lithographically
defined nanopatterned substrates. Nature 2003;424:411414.
71. Pins GD, Bush KA, Cunningham LP, Campagnola PJ. Multiphoton excited
fabricated nano and micropatterned extracellular matrix proteins direct cellular
morphology. J Biomed Mater Res A 2006;78:194204.
72. Wojciak-Stothard B, Curtis A, Monaghan W, MacDonald K, Wilkinson C.
Guidance and activation of murine macrophages by nanometric scale topography.
Exp Cell Res 1996;223:426435.
73. Whitesides GM, Ostuni E, Takayama S, Jiang X, Ingber DE. Soft lithography in
biology and biochemistry. Annu Rev Biomed Eng 2001;3:335373.
74. Kane RS, Takayama S, Ostuni E, Ingber DE, Whitesides GM. Patterning proteins
and cells using soft lithography. Biomaterials 1999;20:23632376.
318
BIOMEDICAL NANOSTRUCTURES
75. Liliensiek SJ, Campbell S, Nealey PF, Murphy CJ. The scale of substratum
topographic features modulates proliferation of corneal epithelial cells and corneal
fibroblasts. J Biomed Mater Res A 2006;79:185192.
76. Araki-Sasaki K, et al. An SV40-immortalized human corneal epithelial cell line and
its characterization. Invest Ophthalmol Vis Sci 1995;36:614621.
77. DeCaprio JA, et al. Sv40 large tumor antigen forms a specific complex with the
product of the retinoblastoma susceptibility gene. Cell 1988;54:275283.
78. Fatt I. Architecture of the lid-cornea juncture. CLAO J 1992;18:187192.
79. King-Smith PE, et al. The thickness of the human precorneal tear film: evidence
from reflection spectra. Invest Ophthalmol Vis Sci 2000;41:33483359.
80. Karuri NW, et al. Biological length scale topography enhances cell-substratum
adhesion of human corneal epithelial cells. J Cell Sci 2004;117:31533164.
81. Chung TW, Liu DZ, Wang SY, Wang SS. Enhancement of the growth of human
endothelial cells by surface roughness at nanometer scale. Biomaterials
2003;24:46554661.
82. Diehl KA, Foley JD, Nealey PF, Murphy CJ. Nanoscale topography modulates
corneal epithelial cell migration. J Biomed Mater Res A 2005;75:603611.
83. Sastry SK and Burridge K. Focal adhesions: a nexus for intracellular signaling and
cytoskeletal dynamics. Exp Cell Res 2000;261:2536.
84. Meyle J, Gultig K, Wolburg H, von Recum AF. Fibroblast anchorage to
microtextured surfaces. J Biomed Mater Res 1993;27:15531557.
85. Uttayarat P, Toworfe GK, Dietrich F, Lelkes PI, Composto RJ. Topographic
guidance of endothelial cells on silicone surfaces with micro- to nanogrooves:
orientation of actin filaments and focal adhesions. J Biomed Mater Res A 2005;75:
668680.
86. Curtis AS, et al. Cells react to nanoscale order and symmetry in their surroundings.
IEEE Trans Nanobioscience 2004;3:6165.
87. Dalby MJ, et al. Nanotopographical stimulation of mechanotransduction and
changes in interphase centromere positioning. J Cell Biochem 2006;100:326338.
88. Tan KS, Qian L, Rosado R, Flood PM, Cooper LF. The role of titanium surface
topography on J774A. 1 macrophage inflammatory cytokines and nitric oxide
production. Biomaterials 2006;27:51705177.
89. Zhu B, Lu Q, Yin J, Hu J, Wang Z. Alignment of osteoblast-like cells and cell-
produced collagen matrix induced by nanogrooves. Tissue Eng 2005;11:825834.
90. Zinger O, et al. Differential regulation of osteoblasts by substrate microstructural
features. Biomaterials 2005;26:18371847.
91. Teixeira AI, et al. The effect of environmental factors on the response of human
corneal epithelial cells to nanoscale substrate topography. Biomaterials 2006;27:
39453954.
92. Foley JD, Grunwald EW, Nealey PF, Murphy CJ. Cooperative modulation of
neuritogenesis by PC12 cells by topography and nerve growth factor. Biomaterials
2005;26:36393644.
93. King MR. Anisotropic Brownian diffusion near a nanostructured surface. J Colloid
Interface Sci 2006;296:374376.
CELLULAR BEHAVIOR ON BASEMENT 319

CHAPTER 12
Focal Adhesions: Self-Assembling
Nanoscale Mechanochemical
Machines that Control Cell Function
TANMAY LELE and DONALD E. INGBER
12.1 INTRODUCTION
Living cells and organisms are constructed through hierarchical self-assembly
of nanoscale molecular components. In the past, the primary focus in biology
was on analysis of the composition and structure of individual biochemical
constituents. However, it has become increasingly clear that the organic
properties of living cells, including their ability to change shape, move, and
grow, are a function of biological architecture how these molecular
components are positioned in space and connected to each other so as to
produce struc tures with specialized forms and biochemical functions [1, 2]. This
is true whether at the scale of a few molecules that come together to form a
single multicomponent enzyme or at the level of the whole cell where different
types of multimolecular, nanoscale filaments, bundles, and tubes join together
to form the molecular framework of the cell, known as the ‘‘cytoskeleton’’.
Because certain molecular filaments in the cytoskeleton actively generate
mechanical (e.g., contractile) forces through an actomyosin filament sliding
mechanism (like in muscle), all of the components of the cytoskeleton must
physically resist these forces, as well as external mechanical stresses, to maintain
cell shape stability [1]. At the same time, the cell possesses a mechanism to
respond to these physical stresses by remodel ing these very same structures (e.g.,
adding new components where they are needed and removing them where they
are not). This ability to convert mechanical signals into biochemical signals and
vice versa is one form of ‘‘mechanotransduction,’’ a fundamental property of
BiomedicalNanostructures, Editedby KennethE.Gonsalves,CraigR.Halberstadt,CatoT.Laurencin,
and Lakshmi S. Nair
Copyright # 2008 John Wiley & Sons, Inc.
321
virtually all living materials [3]. If we could understand how structures are
constructed at the nanometer scale so as to provide these linked mechanical and
chemical functions, we might be able to revolutionize materials design by
creating multifunctional ‘‘biomimetic’’ materials that mimic these properties for
medical, industrial, and military applications.
Another key feature of living materials that makes it possible for them to
interconvert mechanical and chemical energy is that the molecules that
comprise many of structural scaffolds in cells, including the cytoskeleton, also
carry out biochemical functions, or physically associate with chemically active
molecules in the cytoplasm. This structure-based form of metabolism has been
termed ‘‘solid-state biochemistry’’ [2]. Thus, for example, mechanical stresses
that are exerted on the surface of the cell and transferred to the internal
cytoskeleton can influence biochemical activities of some of its load-bearing
components. Biochemical signaling may be triggered through stress-dependent
alterations of molecular shape, which change biophysical parameters such as
molecular binding affinities or opening and closing rates of ion channels [4].
Thus, to understand biological nanostr uctures and to mimic their novel
properties in futur e man-made materials, it is necessary to understand how
living cells are able to build and organize macromolecular scaffolds that
provide critical structural and biochemical functions, as well as trans duce
mechanical and chemical signals at the nanometer scale.
In this chapter, we focus on specialized cell anchoring complexes, known as
‘‘focal adhesions,’’ as an example of self-assembling nanostructures that carry
out all of these functions. These are particularly interesting because they
physically bridge load-bearing extracellular matrix (ECM) scaffolds on the
outside of the cell with the intracellular cytoskeleton as a result of binding
interactions with transmembrane integrin receptors [5–7]. These multimolecular
linkages provide a mechanism to channel mechanical stresses from ECM
molecules to intracellular focal adhesion proteins that, in turn, anchor to the
actin cytoskeleton [6]. In addition, the macromolecular scaffolds that comprise
the focal adhesion grow and shrink when stresses applied to integrins are
increased or decreased, respectively [8]; and numerous signal transducing
molecules are concentrated within these structures [9, 10]. Thus, focal adhesions
are a major site for mechanochemical conversion inside the cell, and the dynamic
assembly of these structures is critical for whole cell behaviors, including growth,
differentiation, and movement. In this chapter, we therefore review the concepts
of solid-state biochemistry, macromolecular assembly, cellular mechanotrans-
duction, and mechanoregulation of cell behavior using the focal adhesion as the
paradigm for a self-assembling nanoscale mechanochemical machine. We also
discuss the implications of these findings for future biomaterials design.
12.2 SOLID-STATE BIOCHEMISTRY IN FOCAL ADHESIONS
The control of cell behavior has been traditionally viewed as being triggered
primarily by soluble stimuli (e.g., hormones, growth factors). These factors
322 BIOMEDICAL NANOSTRUCTURES
bind to surface receptors on the outside of the cell and alter their
conformation. This change in molecular shape triggers a cascade of
biochemical events a process known as signal transduction that ultimately
results in control of cell behavior. However, over the last decade, it has become
clear that many of the molecules that mediate signal transduction are not
floating free in the cytoplasm or lipid bilayer, and instead function when
physically immobilized on insoluble molecular scaffolds inside the cytoplasm
and nucleus, such as the cytoskeleton and nuclear matrix [11].
Signaling proceeds on these self-assembled, multimolecular scaffolds
through binding interactions between several constituent macromolecules.
Furthermore, there is an even higher level of complexity because the
multiprotein complexes that form these insoluble scaffolds are constantly
assembling and disassembling insi de living cells, and this assembly may be
controlled by physical cues like mechanical forces [12, 13]. Thus, the focal
adhesion is an outstanding example of this type of insoluble multimolecular,
mechanochemical scaffold that serves to mediate cell adhesion, spreading,
contraction, and movement on ECM substrates.
Unlike a water droplet spreading on a surface, cells adhere to ECM at
discrete sites. Cell substrate adhesions assemble in these locations when
transmembrane integrin receptors ligate ECM molecules (e.g., fibronectin,
laminin, vitronectin, various collagen types) in the external environment and
cluster together within the surface membrane [7]. This causes recruitment of a
variety of molecules to the inner surface of the clustered integrins, includi ng
focal adhesion scaffold proteins that physically couple integrins to tensed actin
microfilaments, such as talin, vinculin, a-actinin, paxillin, zyxin, and filamin,
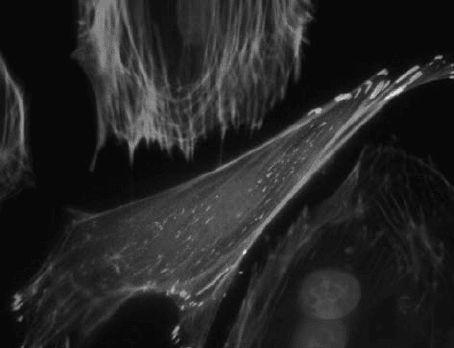
thereby mechanically anchoring the ECM to the cytoskeleton (Fig. 12.1). Other
FIGURE 12.1 Fluorescence image of a single capillary endothelial cell expressing
GFP-vinculin (green), stained for F-actin with Alexa 468-phalloidin (red), and nuclei
with DAPI (blue). Note how each actin stress fiber anchors to focal adhesions at its
distal ends (bar ¼ 10 mm).
FOCAL ADHESIONS: SELF-ASSEMBLING NANOSCALE MECHANOCHEMICAL MACHINES 323
proteins that are simultaneously recruited to the cytoskeletal backbone of the
integrin-associated focal adhesion include various signal transducing mole-
cules, such as protein kinases (tyrosine, serine, and threonine), protein phos-
phatases, inositol lipid kinases, ion channels, G proteins, and certain growth
factor receptors [14, 15]. In fact , it is now clear that focal adhesions are
extremely complex macromolecular assemblies consisting of at least 50
different proteins [10]. These various proteins play unique functional roles in
mediating focal adhesion assembly, as well as providing the crucial link
between the cellECM interface and biochemical signaling pathways that
regulate cellular behavior.
Ultrastructural studies of focal adhesion architecture have revealed that
they measure 60 nm in thickness, with areas of a few square micrometers in
the plane parallel to the ECM substrate [16]. Use of two-photon microscopy
with fluorescent membrane probes has also revealed that these are higher
ordered portions of the cell membrane, which may, in turn, influence structural
organization of associated lipid rafts [17] . These adhesion structures can vary
in size and shape within the same cell because they form, mature, and
disassemble over time [18].
A mature focal adhesion is the culmination of a complex process that begins
with the formation of small dot-like adhesions, called ‘‘focal complexes,’’ at the
leading edge of a motile cell [18], which differ in their chemical composition
from mature focal adhesions [14]. Nascent focal complexes have significantly
low concentrations of certain focal adhesion proteins, such as a -actinin,
vinculin, paxillin, FAK, and VASP [14]. Interestingly, zyxin, a protein presen t
abundantly in mature focal adhesions, is completely absent in focal complexes
in certain cells. Many focal complexes are fated to dissolve away, but some
survive and grow in size to form mature focal adhesions in a tension-dependen t
manner [14].
Exertion of sustained, polarized tensional forces by or on cells causes some
focal adhesions to reorganize and move toward the nucleus, resulting into
linear, streak-like shaped anchoring structures called fibrillar adhesions [19].
While focal complexes and focal adhesions are predominantly present close to
the cell periphery, fibrillar adh esions are more localized in the central regions of
the cell. Fibrillar adhesions are enriched in a5b1 integrin as opposed to aVb3
integrin, which is primarily found in focal adhesions [20]. Fibrillar adhesions
also have elevated levels of tensin and low levels of phosphotyrosine compared
to focal adhesions [20]. Thus, focal complexes, focal adhesions, and fibrillar
adhesions differ not only in their shape, size, and subcellular localization, but
also in their molecular composition.
12.3 FOCAL ADHESION AS A MECHANOTRANSDUCTION MACHINE
Because focal adhesions form a molecular bridge between ECM, integrins, and
the contractile actin cytoskeleton, they provide a preferential path for the
324 BIOMEDICAL NANOSTRUCTURES
transfer of mechanical forces across the cell surfa ce [6]. This is true whether it
involves transmission of internal cytoskeletal traction forces to the ECM
substrate, which are critical for cell migration, muscle contraction, and cell
shape changes, or for transfer of external mechanical stresses to the
cytoskeleton and nucleus [21].
Importantly, the focal adhesion does not just passively transmit force to
associated biochemical signal transducing molec ules, it also changes its own
assembly in response to forces that impinge on it [22, 23]. For example, focal
adhesion area directly correlates with the level of traction force exerted on
these sites at the substrate interface in stationary (nonmotile) cells [24, 25].
When tension in contractile microfilaments is relaxed by chemically inhibiting
myosin II-mediated contractility, time-dependent disassembly of preexisting
focal adhesions occurs and new adhesions do not form [26, 27]. Application of
fluid shear stresses to the apical cell surface or mechanical strain to whole cells
causes remodeling of focal adhesions at the base, in part, through stress-
dependent activation of additional integrin receptors [28, 29]. Forces applied to
the apical surface of the cell through a micropipette can also induce adhesion
assembly at the cell base in a myosin II-independent manner [8]. This appears
to be mediated by force channeling through the cytoskeleton [5, 6, 21,30 33].
When cells are cultured on ECMs that vary in stiffness, focal adhesions also
have a smaller average size on soft versus rigid substrates; this is because cells
are not able to exert significant traction forces on the softer matrices [34].
Taken together, these observations support the hypothesis that mechanical
forces control focal adhesion assembly.
Various stimuli including externally applied mechanical forces, ECM
ligand affinity [35], ECM mechanics [36], nanoscale ligand distribut ion, and
substrate topology [37, 38] regulate the composition and the concentration of
macromolecules that localize within focal adhesions. Focal adhesion assembly
also regulates soluble signaling pathways, including Erk signaling that
controls cell growth [39]. Integrin ligation triggers a variety of signal
transduction pathways that modulate cell shape, gene expression, differentia-
tion, and apoptosis [39–45]. However, the cell’s response to solubl e growth,
motility, and survival factors also depends on the ability of integrins to
transfer transmembrane mechanical signals across the cell surface and to the
focal adhesion. For example, force transfer to the focal adhesion mediated by
integrincytoskeletal connections induces a variety of responses including
cAMP signaling [46, 47], Ca
2+
influx (through mechanosensitive ion
channels), cytoskeletal remodeling [48], alterations of cell shape [49, 50],
and changes in nuclear morphology [21]. These latter effects on cell and
nuclear shape are equally important regulators of cell growth, differentiation,
contractility, motility, and survival as soluble cytokines and hormones
[51–56]. Thus, the focal adhesion is really a nanoscale mechanoch emical
machine that transduces mechanical forces into intracellular bioch emical
signals, and therefore mediates both chemical and physical control of cellular
physiology by ECM and mechanical forces.
FOCAL ADHESIONS: SELF-ASSEMBLING NANOSCALE MECHANOCHEMICAL MACHINES 325