Keyfitz N., Caswell H. Applied Mathematical Demography
Подождите немного. Документ загружается.


13.1. Eigenvalue Sensitivity 305
13.1 Eigenvalue Sensitivity
13.1.1 Perturbations of Matrix Elements
We begin with the sensitivity of an eigenvalue λ to changes in the ma-
trix elements a
ij
. The functional dependence of λ on the a
ij
is expressed
implicitly in the characteristic equation
det(A − λI)=0.
When the form of the characteristic equation is known, implicit differenti-
ation can be used to find ∂λ/∂a
ij
(Hamilton 1966, Demetrius 1969, Emlen
1970, Goodman 1971, Keyfitz 1971a, Mertz 1971a). We will use this ap-
proach in Section 13.1.6, but it is limited to cases in which the characteristic
equation is sufficiently simple that implicit differentiation yields a simple
solution.
A general approach, applicable to matrices of any structure, was intro-
duced by Caswell (1978). The formula dates back at least to Jacobi (1846),
and has been rediscovered many times (e.g., Faddeev 1959, Faddeev and
Faddeeva 1963, Desoer 1967). For a complete and rigorous discussion, see
Kato (1982). Cohen (1978, p. 186) presents an alternative approach to
eigenvalue sensitivity.
We begin with the equations defining the eigenvalues and the right and
left eigenvectors:
Aw
i
= λ
i
w
i
(13.1.1)
v
∗
i
A = λ
i
v
∗
i
, (13.1.2)
where v
∗
i
is the complex conjugate transpose of v
i
. For the moment, let
us suppress the subscript i; the following formulae apply to any of the
eigenvalues and their corresponding right and left eigenvectors. Taking the
differential of both sides of (13.1.1) yields
A(dw)+(dA)w = λ(dw)+(dλ)w, (13.1.3)
where dA =(da
ij
) is a matrix whose elements are the differentials da
ij
.
Form the scalar product
∗
of both sides with the left eigenvector v,
A(dw), v + (dA)w, v = λ(dw), v + (dλ)w, v. (13.1.4)
Expanding the scalar products and cancelling terms leaves
dλ =
(dA)w, v
w, v
(13.1.5)
=
v
∗
dAw
v
∗
w
. (13.1.6)
∗
The scalar product of the vectors x and y is x, y = y
∗
x.
306 13. Perturbation Analysis of Matrix Models
Suppose that only one element, a
ij
, is changed, while all the others are
held constant. Then dA contains only one nonzero entry, da
ij
,inrowi and
column j, and (13.1.6) reduces to
dλ =
¯v
i
w
j
da
ij
w, v
, (13.1.7)
where ¯v
i
is the complex conjugate of v
i
. Dividing both sides by da
ij
and
rewriting the differentials as partial derivatives (since all but one of the
variables of which λ is a function are being held constant), we get the
formula
†
∂λ
∂a
ij
=
¯v
i
w
j
w, v
. (13.1.8)
That is, the sensitivity of λ to changes in a
ij
is proportional to the
product of the ith element of the reproductive value vector and the jth
element of the stable stage distribution. The scalar product term in the
denominator is independent of i and j, and can be ignored when considering
the relative sensitivities of λ to different elements in the same matrix. Or,
the eigenvectors may be scaled so that w, v = 1, so that the term can be
ignored.
It is easy to calculate a sensitivity matrix S whose entries give the
sensitivities of λ to the corresponding entries of A:
S =
∂λ
∂a
ij
(13.1.9)
=
¯
vw
T
w, v
. (13.1.10)
The sensitivity is the local slope of λ, considered as a function of a
ij
.
13.1.2 Sensitivity and Age
The sensitivity of λ to changes in age-specific survival and fertility plays
an important role in the theory of senescence (Medawar 1952, Williams
1957, Hamilton 1966, Rose 1984, Wachter and Finch 1997, Carey 2003).
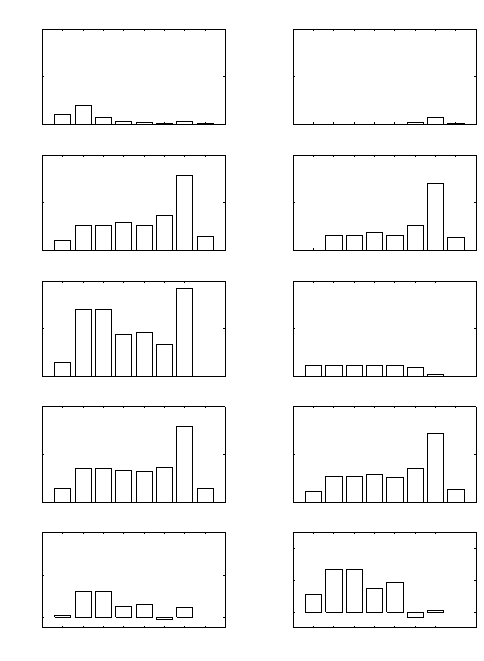
Figure 13.1 shows these sensitivities for four populations (a flour beetle,
a vole, a whale, and humans). Sensitivities of λ to fertility decline nearly
exponentially with age. At early ages, λ is more sensitive to changes in
survival than to changes in fertility. At older ages, the pattern is reversed.
The sensitivities vary by as much as eight orders of magnitude.
†
This formula is often written with v
i
in place of ¯v
i
. This makes no difference in the
analysis of λ
1
, since v
1
is real, but could cause errors in computing sensitivities of other
eigenvalues, and of eigenvectors.

13.1. Eigenvalue Sensitivity 307
0 10 20
10
−8
10
−7
10
−6
10
−5
10
−4
10
−3
10
−2
10
−1
10
0
Age
Sensitivity of λ
C. oryzae
F
i
P
i
0 10 20
Age
M. orcadensis
0 25 50
Age
O. orca
0 25 50
Age
H. sapiens
Figure 13.1. The sensitivity of λ to changes in age-specific fertility F
i
and survival
probability P
i
for (from left to right) a laboratory population of the flour beetle
Calandra oryzae (Birch 1948), a laboratory population of the vole Microtus orca-
densis (Leslie et al. 1955), the killer whale Orcinus orca (Olesiuk et al. 1990), and
the human population of the United States in 1965 (Keyfitz and Flieger 1968).
Some of these properties can be derived directly from the sensitivity
formula. The eigenvectors w and v for age-classified populations are
w
1
= 1 (13.1.11)
w
i
= P
1
P
2
···P
i−1
λ
−i+1
for i>1, (13.1.12)
and
v
1
= 1 (13.1.13)
v
i
= F
i
λ
−1
+ P
i
λ
−1
v
i+1
for i>1. (13.1.14)
We can use w and v to show how the sensitivities change with age
(Demetrius 1969, Caswell 1978, 1982c). From (13.1.8) and (13.1.12) it
follows that
∂λ/∂F
j
∂λ/∂F
j+1
=
w
j
w
j+1
(13.1.15)
=
λ
P
j
. (13.1.16)
Thus the sensitivity of λ to fertility is a strictly decreasing function of
age as long as λ>1. If the P
j
are constant, then the decrease will be
308 13. Perturbation Analysis of Matrix Models
exponential, as is approximately true in Figure 13.1. Other things being
equal, the sensitivity of λ to changes in fertility falls off with age more
rapidly the greater the value of λ. In a decreasing population, however, the
sensitivity will actually increase from age j to j +1ifλ<P
j
.
The sensitivities of λ to changes in survival at successive ages satisfy
∂λ/∂P
j
∂λ/∂P
j+1
=
w
j
v
j+1
w
j+1
v
j+2
(13.1.17)
=
λ
P
j
F
j+1
λ
−1
+ P
j+1
v
j+2
v
j+2
(13.1.18)
= λ
P
j+1
P
j
+
F
j+1
P
j
v
j+2
(13.1.19)
≥
P
j+1
P
j
if λ ≥ 1. (13.1.20)
Thus the sensitivity of λ to survival decreases monotonically with age pro-
vided λ ≥ 1andP
j+1
≥ P
j
. If survival were age-independent, then the
sensitivity of λ to changes in survival would decrease monotonically with
age provided λ>1.
The relative sensitivities of λ to fertility and survival satisfy
∂λ/∂P
j
∂λ/∂F
j
=
v
j+1
w
j
v
1
w
j
(13.1.21)
=
v
j+1
v
1
. (13.1.22)
Thus λ is more sensitive to survival than to fertility if v
j+1
>v
1
,whichis
true at least up to the age of first reproduction. In Figure 13.1, ∂λ/∂P
j
>
∂λ/∂F
j
for young ages; the inequality is reversed at older ages.
13.1.3 Sensitivities in Stage- and Size-Classified Models
The patterns of sensitivity in size- or stage-classified models can be quite
different from those in age-classified models. Because the stable size distri-
bution may exhibit peaks (as is commonly observed in fish and tree size
distributions), sensitivities need not be monotonic functions of size or stage.
Example 13.1 Size-specific sensitivity in the desert tortoise
Figure 13.2 shows the sensitivity of λ to changes in the F
i
, P
i
,and
G
i
. None of the sensitivities shows a monotonic trend with size. Pop-
ulation growth rate is very sensitive to changes in the vital rates of
size classes 2 and 3, and also size class 7.
Example 13.2 Sensitivity analysis of teasel
In stage-classified models, there may be no variable, like size, against
which sensitivities can be plotted. In this case, it is useful to display
13.1. Eigenvalue Sensitivity 309
0 1 2 3 4 5 6 7 8 9
0
0.2
0.4
Sensitivity of λ ...
to F
i
1 2 3 4 5 6 7 8
0
0.2
0.4
to P
i
1 2 3 4 5 6 7
0
0.2
0.4
to G
i
1 2 3 4 5 6 7 8
0
0.2
0.4
to σ
i
0 1 2 3 4 5 6 7 8 9
0
0.2
0.4
to γ
i
Size class i
0 1 2 3 4 5 6 7 8 9
0
0.2
0.4
Elasticity of λ ...
1 2 3 4 5 6 7 8
0
0.2
0.4
1 2 3 4 5 6 7
0
0.2
0.4
1 2 3 4 5 6 7 8
0
0.2
0.4
1 2 3 4 5 6 7
0
0.02
0.04
Size class i
Figure 13.2. Perturbation analysis of population growth rate for the desert tor-
toise. Left column shows sensitivities, right column elasticities, of λ to F
i
, P
i
, G
i
,
σ
i
, and γ
i
. Calculated from data of Doak et al. (1994), medium-high fertility.
the entire sensitivity matrix S calculated from (13.1.10):
S =
⎛
⎜
⎜
⎜
⎜
⎜
⎜
⎝
0.067 0.028 0.001 0.007 0.001 0.001
0.002 0.001 0.000 0.000 0.000 0.000
0.421 0.174 0.008 0.046 0.008 0.003
2.442 1.011 0.046 0.265 0.047 0.018
16.432 6.801 0.313 1.786 0.315 0.119
47.475 19.650 0.905 5.160 0.911 0.344
⎞
⎟
⎟
⎟
⎟
⎟
⎟
⎠
. (13.1.23)
310 13. Perturbation Analysis of Matrix Models
The matrix can be displayed graphically as a surface plot or an image
plot, or by plotting the entries of vec S (Figure 13.3).
‡
These plots
all show that λ is most sensitive to changes in entries in the lower-
left corner of the matrix. These entries correspond to growth directly
from seeds to large rosettes and flowering plants. Such growth doesn’t
happen in this species (i.e., these entries of A are all zero), but that
does not change the fact that if those probabilities were changed, the
impact on λ would be considerable.
13.1.4 What About Those Zeros?
The sensitivity (13.1.7) gives the effect on λ of changes in any entry of A,
including those that may, in a given context, be regarded as fixed at zero or
some other value. This is exactly as it should be. The derivative tells what
would happen to λ if a
ij
were to change, not whether, or in what direction,
or how much, a
ij
actually changes (Caswell et al. 2000). The results of
such impossible perturbations may or may not be of interest, but they are
not zero. It is up to you to decide whether they are useful and whether to
display them.
For example, in the projection matrix for teasel, a
66
= 0 because flow-
ering plants neither survive nor produce new flowering plants in one year.
But from the sensitivity matrix (13.1.23) we see that
∂λ
∂a
66
=0.344.
If a
66
were changed by a small amount ∆a
66
, the resulting change in λ
would be approximately 0.344∆a
66
. This bit of information would be of
no interest to someone concerned with environmental perturbations, but
very interesting indeed to someone studying the evolution of annual and
biennial life histories.
13.1.5 Total Derivatives and Multiple Perturbations
The sensitivity (13.1.8) is written as the partial derivative of λ with respect
to a
ij
, holding all other parameters constant. If more than one parameter
is perturbed simultaneously, the net effect on λ is given by the differential
dλ =
k,l
∂λ
∂a
kl
da
kl
. (13.1.24)
This result can be used in several ways.
‡
The vec operator produces a vector from a matrix by stacking the columns one
above the other.
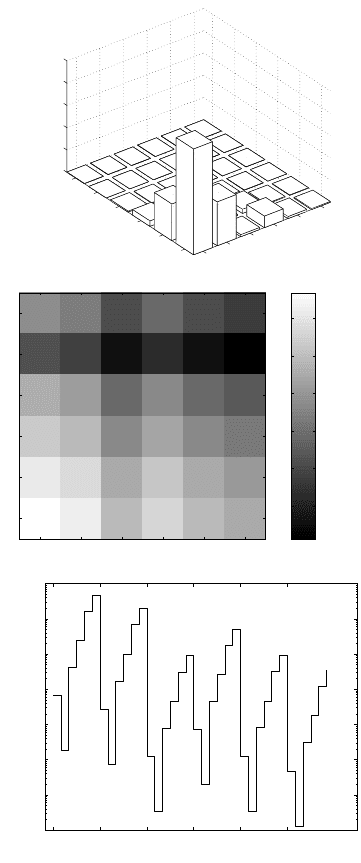
13.1. Eigenvalue Sensitivity 311
1
2
3
4
5
6
1
2
3
4
5
6
0
10
20
30
40
50
Sensitivity of λ
Log
10
sensitivity of λ
−4
−3
−2
−1
0
1
1 2 3 4 5 6
1
2
3
4
5
6
1 2 3 4 5 6
10
−5
10
−4
10
−3
10
−2
10
−1
10
0
10
1
10
2
Column
Sensitivity of λ
11
31
21
41
51
61
12
22
32
42
52
62
13
23
33
43
53
63
14
24
34
44
54
64
15
25
35
45
55
65
16
26
36
46
56
66
Figure 13.3. Three ways of displaying the sensitivity matrix S for teasel (Field
A). The upper graph plots sensitivities as a surface, the middle graph as an
image, and the lower graph as a plot of vec S. In the lower graph, the numbers
are coordinates (i, j) of the entry of S.
312 13. Perturbation Analysis of Matrix Models
1. Lower-level parameters. Suppose that one or more of the matrix
entries are functions a
ij
(x) of some “lower-level” variable x (e.g.,
allocation of energy to reproduction, which affects both survival and
fertility). Then
da
kl
=
∂a
kl
∂x
dx. (13.1.25)
Substituting this expression into (13.1.24) yields the chain rule
dλ
dx
=
k,l
∂λ
∂a
kl
.
∂a
kl
∂x
(13.1.26)
2. Physiological or genetic constraints. One vital rate may be function-
ally related to another, for many reasons. Increased fertility may
result in decreased somatic growth, because of allocation of resources.
Increased survival early in life may result in decreased survival later
in life due to pleiotropic effects. We suppose that perturbing one vital
rate a
ij
will affect some or all of the other matrix entries, so that
da
kl
=
∂a
kl
∂a
ij
da
ij
. (13.1.27)
Substituting into (13.1.24), we obtain
dλ
da
ij
=
∂λ
∂a
ij
+
kl=ij
∂λ
∂a
kl
∂a
kl
∂a
ij
. (13.1.28)
3. A special case of constraints arises when individuals face mutually
exclusive options. Consider seeds that may either germinate or remain
dormant. Clearly, P [germination] = 1 − P [dormancy], and a change
in one must produce a compensatory change in the other.
Example 13.3 Sensitivity to survival and growth in size-classified
models
The parameters G
i
and P
i
in the standard size-classified model for
the desert tortoise are functions of the survival probability (σ
i
)and
the growth probability (γ
i
) of size class i. From (13.1.26) we have
∂λ
∂σ
i
=
∂λ
∂G
i
∂G
i
∂σ
i
+
∂λ
∂P
i
∂P
i
∂σ
i
(13.1.29)
=
∂λ
∂G
i
γ
i
+
∂λ
∂P
i
(1 − γ
i
) (13.1.30)
=
w
i
(v
i
+ γ
i
∆v
i
)
w, v
, (13.1.31)
13.1. Eigenvalue Sensitivity 313
where ∆v
i
= v
i+1
− v
i
is the change in reproductive value from size
class i to i + 1. Similarly,
∂λ
∂γ
i
=
∂λ
∂G
i
∂G
i
∂γ
i
+
∂λ
∂P
i
∂P
i
∂γ
i
(13.1.32)
= σ
i
∂λ
∂G
i
−
∂λ
∂P
i
(13.1.33)
=
σ
i
w
i
∆v
i
w, v
. (13.1.34)
The sensitivity of λ to growth rate (13.1.34) is negative if v
i+1
<v
i
.
That is, λ is reduced by increasing the growth rate from N
i
to N
i+1
if stage i + 1 has a lower reproductive value than stage i.
Applying (13.1.31) and (13.1.34) to the size-classified model for the
desert tortoise yields the results shown in Figure 13.2. This untangles
the survival and growth components of the sensitivities to P
i
and G
i
.
Changes in the survival of stage 7 would have a major impact on
λ. The sensitivity of λ to γ
6
is slightly negative because of a slight
decline in reproductive value from stage 6 to stage 7.
In the desert tortoise, the fertilities F
i
were considered independent of
survival and growth. More detailed descriptions of the F
i
, however, usually
involve both the σ
i
and γ
i
. In such cases, the sensitivity of λ to the lower-
level parameters must also include their effects on fertility.
13.1.6 Sensitivity to Changes in Development Rate
Population growth rate is sensitive to changes in the timing of events in
the life cycle (e.g., Lewontin 1965, Mertz 1971, Caswell and Hastings 1980,
Caswell 1982c, Hoogendyk and Estabrook 1984, Ebert 1985). In Section
6.3 we saw the effect on the intrinsic rate of increase r =logλ of changes
in the mean µ and variance σ
2
of the net maternity function. Increasing µ
corresponds to a delay in development, and an analysis using age-classified
matrices yields the same conclusion as (6.3.5); i.e., that in an increasing
population, delayed reproduction reduces population growth rate.
However, this analysis is based on the transformation of the charac-
teristic equation into a cumulant generating function, and holds only for
age-classified models. What can we say about slowing the rate of transition
in a general stage-classified model?
To answer this question, we start with a transformation of a life cycle
graph (Section 9.1.3). The coefficient on each arrow is multiplied by λ
−α
,
where α is the number of projection intervals required for the transition.
Once transformed, the graph can be simplified by multiplying the coeffi-
cients on pathways between any two stages (see MPM Chapter 7). Figure
13.4 shows such a graph, focusing on the transition from stage j to stage

314 13. Perturbation Analysis of Matrix Models
i
j
1
c
(λ)
e
(λ)
d
(λ)
b
(λ)
a
(λ)
P
λ
−α
Figure 13.4. A life cycle graph for evaluating the sensitivity of λ to changes in α,
the time required for the transition from stage j to stage i.
i, which requires α projection intervals to complete, with a probability P
of surviving. The other arcs in this graph represent
• a(λ) for all paths between N
1
and N
j
that do not pass through N
i
.
• b(λ) for all paths between N
j
and N
1
that do not pass through N
i
.
• c(λ) for all paths between N
i
and N
1
• d(λ) for all paths from N
1
to N
i
which do not pass through N
j
• e(λ) all loops involving neither N
i
nor N
j
.
We assume that there are no disjoint loops in the graph. The following
method can be extended to life cycles containing such loops, but the results
are more complicated. Since the time (α) required for transition from N
j
to N
i
appears explicitly in the transformed life cycle graph, it is possible to
evaluate the sensitivity of λ to changes in development rate between these
two stages.
The characteristic equation can be written down from the graph
§
as
1=a(λ)c(λ)Pλ
−α
+Φ(λ) (13.1.35)
= F(λ), (13.1.36)
where Φ(λ)=a(λ)b(λ)+c(λ)d(λ)+e(λ). Implicit differentiation of λ with
respect to α gives
∂λ
∂α
=
−∂F/∂α
∂F/∂λ
. (13.1.37)
§
By setting equal to 1 the product of the transmissions around all loops in the graph;
see MPM Chapter 7.