Меннинг О. Поведение животных. Вводный курс
Подождите немного. Документ загружается.

12
Глава 1
УТОМЛЕНИЕ
Если мышца сокращается под влиянием регулярно повторяю-
щейся стимуляции двигательного нерва, то она продолжает отвечать
в течение очень длительного времени — нескольких часов или более.
По-другому обстоит дело, если та же самая мышца возбуждается
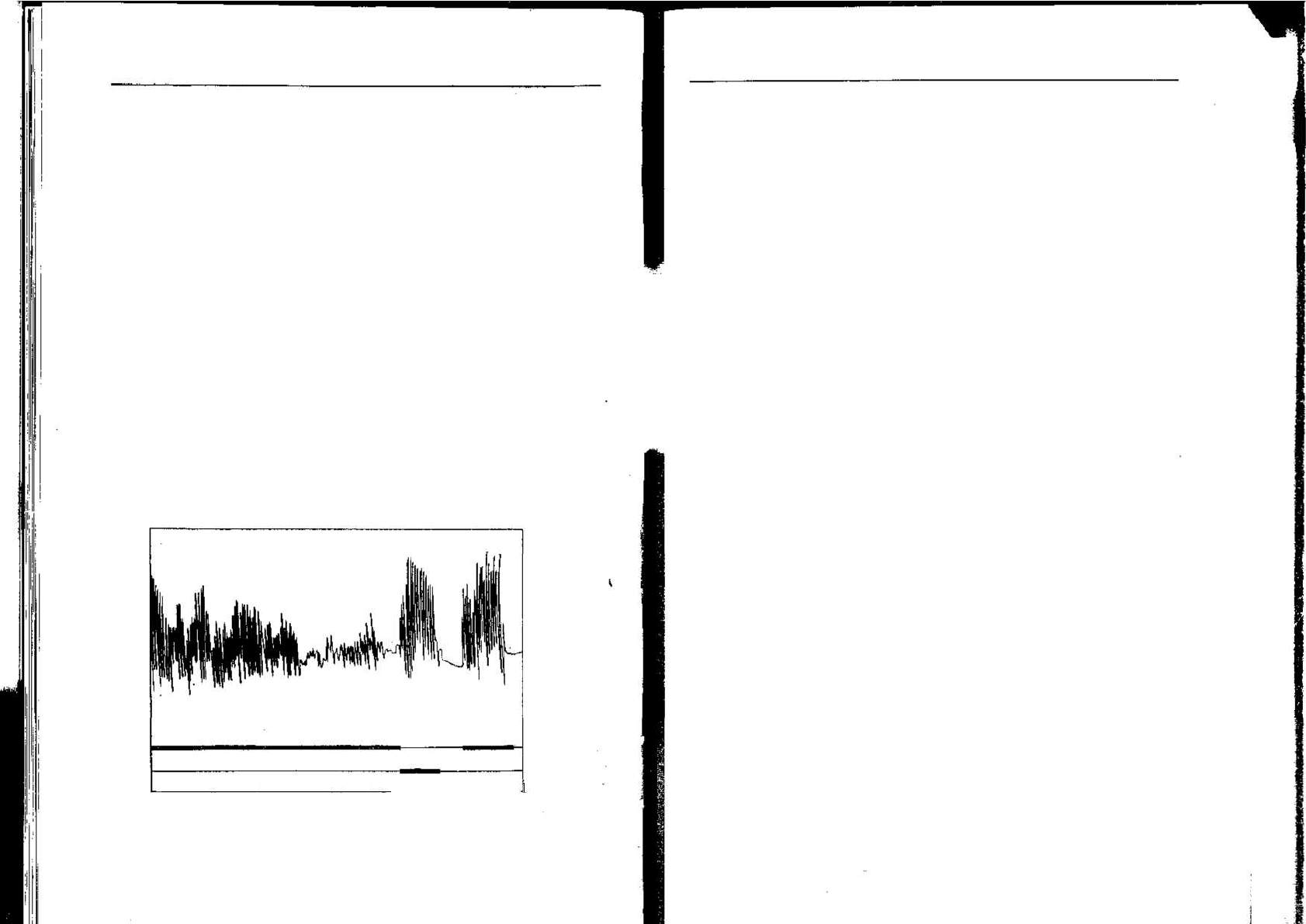
через рефлекторную дугу. Шеррингтон обнаружил, что чесатель-
ный рефлекс у собаки начинает ослабевать приблизительно через
20 с непрерывной механической или электрической стимуляции в
одной и той же точке кожи. Движения лапы становятся слабее и
теряют ритм. Утомление обусловлено, разумеется, не мышцей,
потому что если в это же время вызвать сгибательный рефлекс, ко-
торый обеспечивается теми же мышцами, то он проявится в полной
мере. Если начать стимуляцию кожи в области, отстоящей от пер-
вой в нескольких сантиметрах, то чесательный рефлекс восстано-
вится, а после короткого перерыва стимуляция первой области снова
станет эффективной (рис. 1.4).
Это означает, что утомление должно возникать где-то между
сенсорными органами кожи и началом двигательного нерва. Шер-
рингтон считал, что оно связано с торможением проведения через
синапсы между вставочными нейронами, которые проводят импуль-
сы вниз по спинному мозгу, и мотонейронами.
Францискет [154] показал, что вставочные нейроны являются
местом развития утомления в случае чесания лапой у лягушки.
Если слегка коснуться кожи спины лягушки, то она поднимает
Включение
Выкл.
Вял.
вб/лл/телие Включение
1 1 1 1 М 1 1 1 1 i 1 1 1 1 1 1 1 1 1 1 1, 111111< ЦД 1 1 Ц 1 1 I И.
Точка
А
Ъчка
В
1
Время, с
Рис. 1.4. «Утомление» чесательного рефлекса ([435] с изменениями). Продолжи-
тельная стимуляция точки А в конце концов перестает быть эффективной, но сти-
муляция точки Б немедленно возобновляет рефлекс. Стимуляция точки А снова
вызывает отгет только через 9 с.
РесЬлекш и сложное поведение
21
ногу и один раз потирает спину. После нескольких десятков сти-
мулов наступает утомление, но, как и в примере с собакой, при пере-
мене места стимуляции работа конечности восстанавливается.
Францискет отметил, что легкие судороги в коже лягушки вокруг
места, где ее касались, продолжаются и после того, как конечность
перестала реагировать. Это свидетельствует о том, что сенсорные
нервы кожи все еще работают и к двигательным нервам, которые
управляют отдельными мышцами кожи, передаются импульсы.
Лягушка перестает потирать ногой спину, потому что нарушается
проведение импульсов во вставочных нейронах, связывающих сен-
сорные нервы с двигательными нервами ноги.
Утомление всегда является признаком сложных форм поведения.
Сравним рис. 1.3, где показано, как убывает число криков зябли-
ков при окрикивании совы, с левой частью рис. 1.4, показываю-
щего, как ослабевает чесательный рефлекс собаки. В примере с
зябликом, однако, утомление — это просто обозначение того факта,
что ответ ослабевает, даже если стимул сохраняется. Рассматривае-
мые отдельно подобные наблюдения ничего не говорят нам о ме-
ханизмах, лежащих в основе этого явления, которые, конечно,
различны в этих двух случаях.
Иногда, как и в случае рефлексов, изменение стимула приводит
к возобновлению ответа, ослабленного утомлением. Молодые во-
робьиные птицы выпрашивают пищу у прилетающих к гнезду ро-
дителей, вытягивая шеи и широко раскрывая клювы. Прехтль
[383] обнаружил, что реакцию выпрашивания пищи вызывает лю-
бой темный предмет, появляющийся над краем гнезда. Это же про-
исходит и при сотрясении гнезда, обычно свидетельствующем о
прилете родителей. Когда реакцию выпрашивания многократно вы-
зывают зрительными стимулами, она ослабевает и постепенно ис-
чезает. Если в этот момент потрясти гнездо, то реакция выпраши-
вания снова проявится с полной силой. В таком случаемы можем
назвать исходное утомление «специфичным в отношении стимула»,
так как на стимулы других типов реакция по-прежнему возникает.
Работа Хайнда [211] по реакции окрикивания хищника зябли-
ками показывает, что есть и другие типы утомления, которые мы
можем описать в поведенческих терминах. На рис. 1.5 приведены
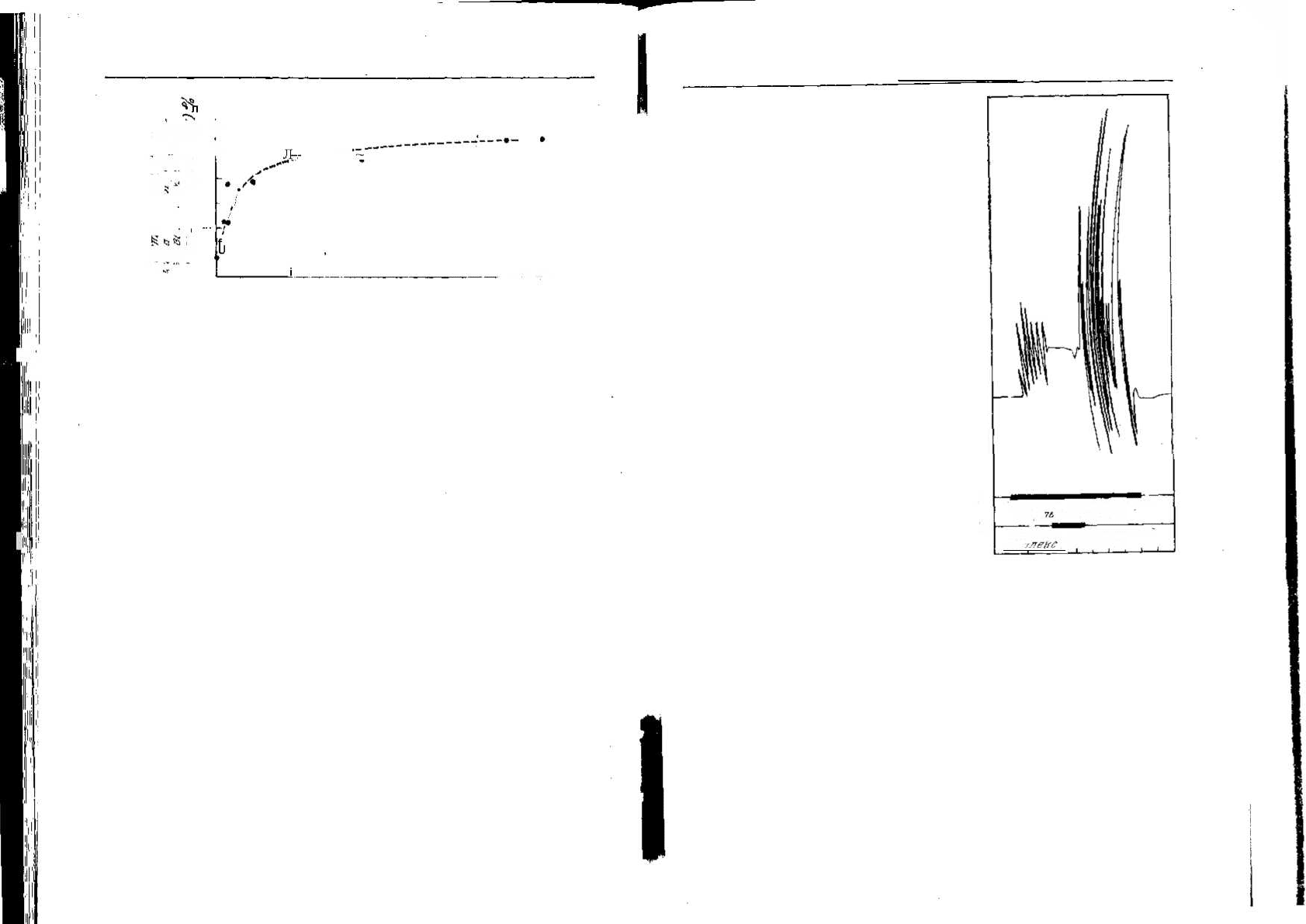
результаты одного из его опытов. Сову подсаживают к зябликам
на 30 мин; по окончании этого времени коллективное окрикивание
обычно прекращается и для оценки силы реакции Хайнд подсчиты-
вает число криков «чинк» в первые 6 мин. Затем сову удаляют из
клетки на периоды от 0,5 мин до 24 ч. При возвращении совы в
клетку для проведения второго теста снова подсчитывают крики
в первые 6 мин. Очевидно, чем короче промежуток между двумя
опытами, тем меньше мы ожидаем, что во втором случае птицы
будут реагировать на сову. Рис. 1.5 показывает, что интенсивность
криков достигает 50% начального уровня при перерыве между опы-
,1
III '!
f
и
I:
I
22
Глава \1
* ^ t 60 [•
1 р 1 5
0
Л
HM
S
,, • * 40
Ш*
30
m
s 5
S
20
«з ч.; н
20
g
^^ 10
^>5; Л
^mmim - - -
t I
_..1. _1_
120 24ч
С> 30 60мин 90
Перерыв между концом первоге опыта
а началом второго
Рис. 1.5. Восстановление реакции окрикивания после длительной экспозиции чу-
чела совы (объяснения см. в тексте) [211].
тами в 30 мин, но при перерыве в 24 ч этот показатель почти не улуч-
шается. Хайнд отмечает, что даже при более длительных перерывах
первоначальный уровень реакции окрикивания никогда не восста-
навливается.
Поскольку сова служила стимулом в обоих опытах, неполное вос-
становление отчасти можно было бы объяснить утомлением, специ-
фичным в отношении стимула, так же как в реакции выпрашивания
у молодых птиц. Однако Хайнд обнаружил, что при использовании
во втором тесте чучела горностая (которое является столь же силь-
ным раздражителем, как и сова) возникает небольшое усиление
криков, но создается почти та же картина, которая зарегистриро-
вана на рис. 1.5. Сначала утомление снимается быстро за первые
30 мин, но затем этот процесс идет чрезвычайно медленно, причем
для него характерна специфичность в отношении стимула, что сов-
падает со свойствами самой реакции «коллективного окрикивания».
Утомление такого рода не может быть результатом действия
тех же механизмов, что и в случае реакции на определенный сти-
мул. Хайнд предполагает, что в действительности здесь идут два
различных процесса: после одного восстановление происходит бы-
стро, а после другого — в очень слабой степени. Мы должны до-
бавить, что слабая степень утомления, специфичного в отношении
стимула, обнаруживается при замене совы горностаем. Это означает,
что возможны три различных вида утомления и процессы восста-
новления происходят одновременно с реакцией зябликов.
Рассмотренные явления достаточно сложны, я, хотя как в слу-
чае рефлексов, так и в случае сложных форм поведения происходит
утомление, ситуация, выявленная в опытах с зябликами, должна
заставить нас остерегаться поспешных выводов относительно его
Рефлексы и сложное поведение
23
механизмов. Каждый конкретный слу-
чай нужно изучать отдельно, только
тогда можно разобраться в лежащих
в его основе процессах с помощью
тщательно поставленных поведенче-
ских опытов.
ТОРМОЖЕНИЕ
Эта категория является не столько
свойством рефлексов, сколько целой
областью функционирования нервной
системы. Одно из величайших дости-
жений Шеррингтона состоит в сле-
дующем: он показал, что в некото-
рых случаях импульсы, проходя по
аксону, тормозят проведение в нерв-
ной клетке, с которой он образует
синапс, даже если клетка при этом
одновременно получает возбуждаю-
щие импульсы по другим аксонам.
Торможение действует на каждом
уровне центральной нервной системы.
Поскольку каждый нейрон потенци-
ально имеет контакт со всеми осталь-
ными, возникает вопрос, почему жи-
вотное не бьется в судорогах после
каждого получаемого им стимула. От-
части это объясняется задержкой и
утомлением при синаптическом про-
ведении, но свою роль играет также
активное торможение.
Мышцы обычно объединены в ан-
тагонистические пары, так что одна
сгибает конечность,а другая разгиба-
ет ее. Шеррингтон показал, что возбу-
ждение одной мышцы сопровождается
торможением ее антагониста. Это тор-
можение не абсолютное, и заторможен-
ная мышца не остается просто расслаб-
ленной. Когда, например, мышца-
разгибатель растягивается в результате сокращения мышцы-сгиба-
теля, под влиянием ее собственного «рефлекса на растяжение» (бо-
лее подробно см. с. 29) она стремится сократиться. Хотя
Л
сгибатель
мог бы преодолеть сопротивление разгибателя, последний перейдет
в расслабленное состояние, так сказать, активным способом. Если
А
В
Стимул, вызывающий,
рктлекс чеса»ия
Стимул, вызываюшии
ррсрлерс ^ czi/fy'wf*,
время> с
Рис. 1.6. Торможение чесатель-
ного рефлекса рефлексом сгиба-
ния ([435] с изменениями). Сти-
мул, обозначенный на линии А,
вызывает рефлекс чесания, но
реакция ингибируется, когда
стимул, обозначенный на линии
Б, вызывает рефлекс сгибания.
В момент прекращения действия
стимула Б рефлекс чесания во-
зобновляется в гораздо более
выраженной форме, чем рань-
ше — пример «рефлекса отдачи».
1

12
Глава 1
РесЬлекш и сложное поведение
25
таким образом удалось бы заставить работать мышцы-антагонисты
одна против другой, то был бы возможен гораздо более тонкий кон-
троль движений.
Взаимное торможение позволяет им по очереди брать верх во
время движений конечности. Шеррингтон нашел, что помимо тормо-
жения мышц-антагонистов одной конечности возможно также анта-
гонистическое влияние во время движения мышц разных конечно-
стей. Когда сгибатели одной конечности сокращаются, на другой
конечности они тормозятся. Реццпрокное торможение этого типа
является одним из основных интегративных механизмов при ходьбе.
В случае рефлексов активное торможение движения легко отли-
чить от утомления. Утомление чесательного рефлекса (рис. 1.4)
выражается в его постепенном ослаблении. Когда тот же рефлекс
тормозится благодаря антагонистическому рефлексу сгибания, он
сразу исчезает без предшествующего ослабления и возобновляется
тотчас же после прекращения действия стимула, вызвавшего сги-
бание (рис. 1.6). Торможение служит для быстрого и бесперебойного
перехода от одного рефлекторного действия к другому.
• Роль торможения в сложном поведении внешне не столь очевидна,
как роль возбуждения. Мы стимулируем животное, и наиболее за-
метным результатом является его реакция. Однако для ее осуще-
ствления должно произойти быстрое переключение, для которого
требуется предшествующее стимулу торможение, необходимое для
приспосабливания рейкций животного к новой ситуации. Шерринг-
тон рассмотрел способ «конкуренции» рефлексов за контроль над
общим конечным путем, т. е. механизмом, который управляет мыш-
цами, участвующими в осуществлении нескольких различных реф-
лексов. Подобным образом мы можем рассматривать различные
системы, которые контролируют виды сложного поведения — пита-
ние, борьбу, сон — как конкурирующие за управление животным.
Очевидно, такие виды поведения несовместимы, и только один из
них возможен в каждый данный момент. Какой именно — это за-
висит от различных внутренних и внешних причин. Когда, напри-
мер, контроль осуществляет пищевая система, все остальные сис-
темы на это время должны быть заторможены.
Иногда мы можем установить, какая часть нервной системы жи-
вотного выполняет тормозные функции, обеспечивающие угнете-
ние активности, которая иначе была бы постоянной. В недавних ис-
следованиях мозга насекомых выявлены механизмы такого рода.
Мозг отбирает подходящие ответы, снимая торможение с соответ-
ствующих отделов брюшной нервной цепочки и ганглиев, которые
затем возбуждают необходимые мышцы. Один из наиболее ярких
примеров этого явления дает работа Рёдера [396] на степном бого-
моле (рис. 1.7). Если самца богомола обезглавить, отделив мозг от
брюшной нервной цепочки, то его тело начинает непрерывно пере-
бирать ногами, двигаясь по кругу. В дополнение к этому брюшко
Третий \
торакальный ганглий [
торакальный гангли. /
Второй таракальА
ньт ганглий
Абдоминальные
ганглии /-б
'Антенна
ffoSsmmowm
ганглии
Рис. 1.7. Схема центральной нервной системы богомола. В опытах Рёдера нервная
цепочка была перерезана по линии А Б [396].
и половые придатки непрерывно совершают копуляторные движения,
и обезглавленный самец зачастую может успешно спариться. В ес-
тественных условиях самцы тоже иногда оказываются обезглавлен-
ными, так как их самки крупны и агрессивны; отсюда следует, что
такое приспособление очень адаптивно!
Показано, что у позвоночных также часто в результате разру-
шения некоторых из областей мозга значительно облегчается сти-
муляция поведения какого-либо одного типа. Как мы увидим в
следующих главах, при трактовке результатов опытов с разруше-
нием мозговых структур необходима осторожность, однако нет
сомнений в том, что одни отделы мозга позвоночных регулируют
активность других отделов с помощью торможения.
Шеррингтон обнаружил, что после прекращения торможения
рефлекс возобновляется с большей интенсивностью. Рис. 1.6 на
примере чесательного рефлекса иллюстрирует это явление, которое
Шеррингтон назвал «рефлексом отдачи». Как правило, если слож-
ное поведение какого-то специфического типа, например ухажива-
ние, некоторое время не проявляется, то, будучи в конце концов
вызвано, оно имеет сниженный порог и повышенную интенсивность.
Возможно, что система мозга, контролирующая ухаживание, была
заторможена системами, контролирующими другие виды актив-
ности, и при устранении торможения обнаруживается что-то близ-
кое к рефлексу отдачи. Кеннеди [265, 266] рассмотрел в этом плане
некоторые аспекты поведения тлей. В поведении крылатых тлей
чередуются периоды полета и оседлого кормления на листьях.
Если тля садится на непривлекательную для нее поверхность, на-
пример на старый лист, она не задерживается там и вскоре улетает,
но летит недолго и скоро снова садится на лист. Напротив, если она
I
26
Глава 1
садится на хороший молодой росток, то остается на нем на длитель-
ное время, а когда улетает, то летит быстро и долго.
В серии изящных экспериментов Кеннеди [266] удалось исклю-
чить любое объяснение этих явлений, основанное просто на физи-
ческом истощении во время полета и восстановлении после отдыха
и питания на молодом листе. Он предполагает, что между системами,
контролирующими поведение тли в полете и во время оседлости,
существует взаимное торможение. Как и в случае рефлексов, из-за
активации системы, контролирующей поведение одного типа (осед-
лость), может временно тормозиться проявление активности систе-
мы, контролирующей полет, но в то же время порог последней сис-
темы постепенно снижается. Кеннеди назвал это «антагонистической
индукцией», которая в какой-то степени сходна с рефлексом от-
дачи.
В гл. 4 и 5 мы обсудим другие доказательства того, что системы,
контролирующие сложные формы поведения, должны тормозить
друг друга и что одни из них «прорываются», когда ослабевает бло-
кирующее их торможение. Однако рефлекс отдачи не может пол-
ностью объяснить, как одни сложные формы поведения сменяют
другие. Снижение порога могут обусловить другие факторы. На-
пример, с позиций физиологии совершенно неуместно говорить о
«торможении питья», если собаке физически недоступна поилка
с водой, но потребность в воде у нее тем не менее растет.
РЕГУЛЯЦИЯ С ПОМОЩЬЮ СИСТЕМЫ ОБРАТНОЙ СВЯЗИ
Мы только что рассмотрели вопрос о том, как тормозные меха-
низмы могут привести к переключению активности от одной систе-
мы контроля на другую. Но очень часто рефлекс или сложная форма
поведения состоит не из внезапных переключений, а скорее из
какой-то постоянной активности, которая должна поддерживаться
на заданном уровне. Когда мы стоим свободно, без напряжения, наше
тело хорошо уравновешено и легко корректирует любые неболь-
шие толчки, которые мы можем получить. Чтобы обеспечить это,
мышцы ног и спины должны поддерживаться в состоянии напряже-
ния на каком-то постоянном уровне, и если этот уровень меняется,
то должна произойти коррекция, благодаря которой тело снова вы-
прямится. Подобно этому животные в нормальных условиях поддер-
живают вес тела на строго постоянном уровне за счет потребления
воды и пищи в правильных соотношениях. Они едят и пьют в соот- .
ветствии со своими потребностями через определенные интервалы.
Если даже они имеют пищу в избытке, то не переедают. Когда пищи -
мало, они тратят больше времени на ее поиски и, когда появляется
возможность, съедают больше, чтобы восполнить дефицит.
Оба примера показывают, каким образом поведение как гоме-
остатический процесс выполняет свою роль в сохранении status
Вефл
Л
сы и сложное поведение
27
quo. В первом случае это обеспечивается относительно простой
системой рефлексов, управляющих мышцами ног и туловища. Во
втором случае имеется набор более сложных систем, регулирующих
поиск пищи, процесс питания и насыщение. В обоих случаях для
функционирования систем требуется, чтобы конечный результат
(поза и равновесие в первом случае, состояние насыщения — во
втором) каким-то образом контролировался. Когда он не соответ-
ствует установленному уровню, в контролирующие механизмы по-
ступает сигнал, позволяющий исправить отклонение и приводящий
систему к заданному уровню.
Инженерам, проектирующим контролирующие системы для ма-
шиностроения, знакомы механизмы, которые функционируют сход-
ным образом. Они называют их системами, действующими по прин-
ципу обратной связи. Некоторые диаграммы, которые мы приводим,
помогут объяснить основные принципы контроля с помощью об-
ратной связи. Мы покажем также, как их можно приложить к реф-
лексам и сложным формам поведения. Выделим два типа контроли-
рующих механизмов — с открытой цепью и с замкнутой цепью
(рис. 1.8, Л и Б).
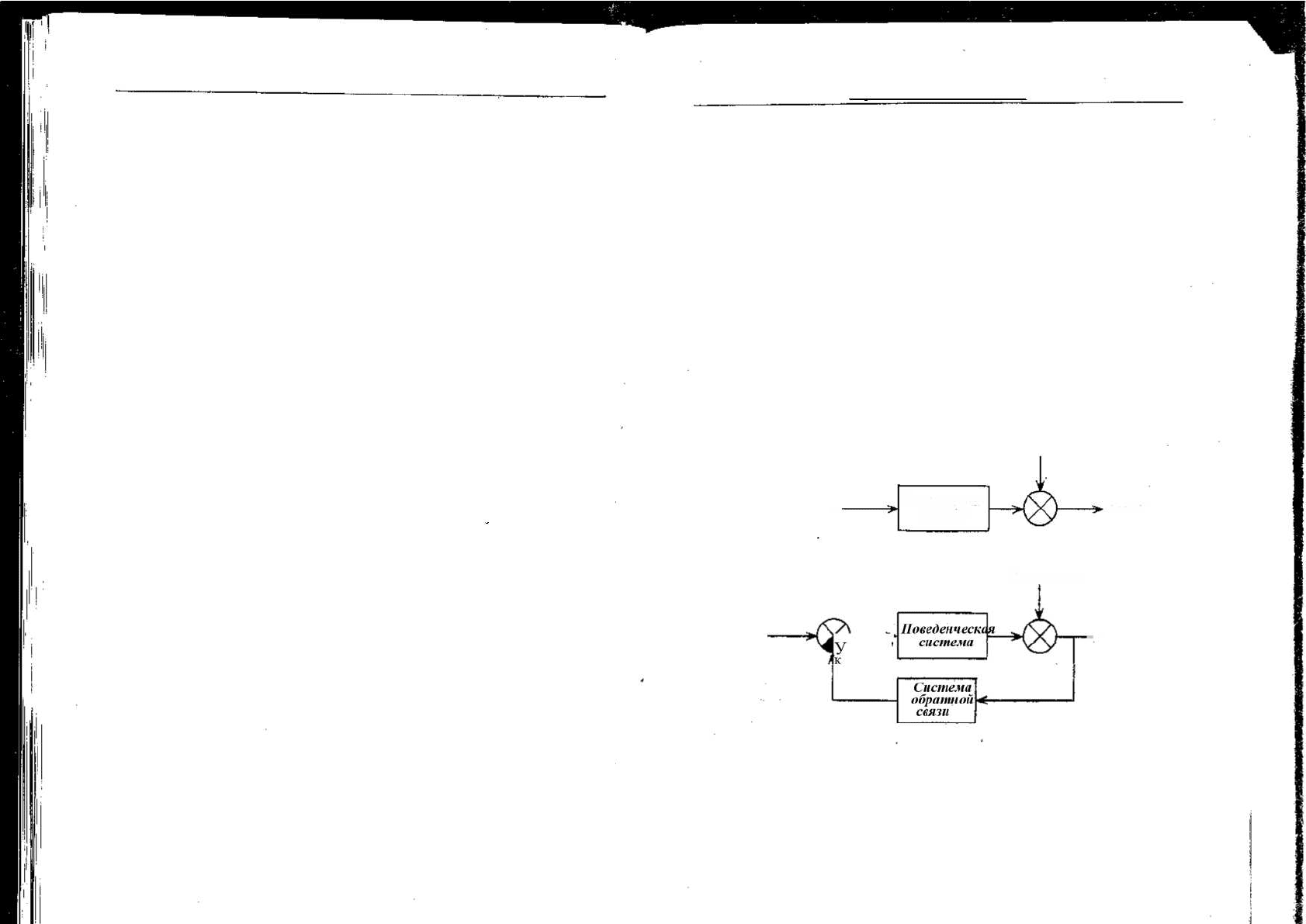
Отклонения
Вход—
Позедвнческая
система
выход
А, Контроль с помощью
открытой цепи
Вход ~
Итлонени*
-> 'Выход
Б. Контроль о помощью
замкнутой цепи
Рис. 1.8. Схема систем регуляции с помощью открытой и замкнутой цепей. На вы-
ходной сигнал поведенческой системы воздействуют факторы отклонения (их взаи-
модействие обозначено кружком с крестом). При контроле с помощью открытой
цепи (А) изменение выходного сигнала не ведет ни к каким поправкам. При конт-
роле с помощью закрытой цепи (Б) изменения на выходе передаются и влияют на
вход в поведенческую систему. Черный сегмент круга изображает взаимодействие
между механизмами обратной связи и входным сигналом, в результате которого
выходной сигнал возвращается к исходному уровню. Система обратной связи про-
дуцирует свой собственный выходной сигнал, который пропорционален отклоне-
нию и изменяет входной сигнал в нужной степени и в нужном направлении.

12
Глава 1
В обеих системах некий механизм (обозначенный на схеме пря-
моугольником) генерирует выходной сигнал в ответ на входной сиг-
нал определенного типа, причем выходной сигнал меняется при
разного рода воздействиях. Кружком, поделенным на квадранты,
обозначен процесс, благодаря взаимодействию с которым осущест-
вляется влияние отклонений.
В системе, изображенной на верхней схеме, отклонения выход-
ного сигнала не корректируются. Если лодка движется опреде-
ленным курсом с закрепленным рулем, а боковые течения сбивают
ее с курса, то нельзя изменить ее новое направление, чтобы компен-
сировать отклонение. Это пример системы с незамкнутой цепью.
В системе с замкнутой цепью (петлей), представленной на нижней
схеме, выходной сигнал измеряется, данные об отклонениях пере-
даются обратно и оказывают влияние на входной сигнал. Обычно
изменяется его степень и направление таким образом, что выход-
ной сигнал возвращается к исходному уровню. Наша лодка в этом
случае имеет автоматического рулевого, который реагирует на не-
благоприятные течения поворотом руля, благодаря чему лодка
удерживается на курсе. Эта ситуация с замкнутой цепью включает
контроль с помощью обратной связи; очевидно, она наиболее при-
менима к рефлексам или сложным формам поведения, которые
связаны с поддержанием гомеостаза. Это не означает, что все виды
поведения контролируются с помощью обратной связи. Было по-
казано, что в некоторых случаях действует система контроля с
открытой цепью. В частности, когда движение должно быть совер-
шено очень быстро с использованием системы нейронов и синапсов,
просто нет времени изменять движение в процессе его выполнения.
«Нападение» богомола, хищнические наклонности которого мы
только что обсуждали,— один из таких случаев. Богомол движется
по направлению к мухе, медленно и точно ориентируя свое тело
(этот процесс, конечно, включает контроль с помощью обратной
связи), но раз нацелившись на жертву, он совершает движение по
принципу «все или ничего». Если муха движется после того, как
нападение началось, это ничего не изменит в характере движения
богомола.
Если вновь обратиться к нашим первоначальным примерам, к
контролю позы или потребления пищи и воды, то станет ясно, что
оба они включают систему обратной связи, которая дает возмож-
ность поддерживать оба процесса на постоянном уровне. Очевидно,
что каждому из них присущ свой особый механизм, но принципы
действия механизмов сходны.
Регуляция позы — один из примеров тонкого мышечного конт-
роля, и физиологи провели полный анализ связанных с ним меха-
низмов. Действительно, в некоторых случаях мы знаем, какие ней-
ронные пути участвуют в данном процессе, и можем идентифици-
ровать структуры, которые функционируют в качестве компонентов
РесЬлекш и сложное поведение
29
Спинной
мозг •
Чувствительный нейрон
мышечного вере/пена
Тело се тарной ялетки
Капсула мышечного веретена
^нтрвфтвло -
4
мое волокно
Bwfywtfawu/m
всшаеатый
нгйрон
Тело метонеар/г/м,
Мшонейром
Рис. 1.9. Упрощенная схема некоторых нервных путей, вовлеченных в рефлекс ра-
стяжения. Объяснения см. в тексте.
системы контроля по замкнутой цепи. На рис. 1.9 схематически
изображена типичная мышца конечности млекопитающего, которая
участвует в поддержании позы. Двигательные нейроны, тела ко-
торых расположены в вентральных рогах спинного мозга, направ-
ляют к мышцам свои волокна, и именно их активность определяет
создаваемое мышцей напряжение. Параллельно каждой скелет-
ной мышце расположены мышечные веретена, уложенные среди ее
волокон таким образом, что они сокращаются и расслабляются
вместе с ней. Это специализированные сенсорные органы для реги-
страции степени натяжения мышцы. Их сенсорные нервы входят
в спинной мозг через дорсальные корешки и контактируют там с
мотонейронами, иннервирующими данную мышцу-. Так замыкается
цепь, которая служит основой для рефлекса растяжения, о котором
уже шла речь. Когда за счет сокращения антагонистов мышца растя-
гивается, мышечные веретена растягиваются вместе с ней и в их
сенсорных волокнах повышается частота разрядов; благодаря этому
мотонейроны стимулируются таким образом, что иннервируемая
ими мышца сокращается. Можно легко соотнести элементы системы
контроля с замкнутой цепью (рис. 1.8, Б) с реальными нервными и
мышечными структурами. Выходной сигнал (степень напряжения
мышцы) изменяется благодаря растяжению других мышц. Это изме-
нение воспринимается механизмом обратной связи (мышечные вере-
тена) и передается в спинной мозг, после чего изменяется входной
сигнал (посылаемый двигательным нервом) и восстанавливается
первоначальный выходной сигнал.
Это упрощенная картина реальной ситуации, которая на самом
Деле включает в себя и другие регуляторные механизмы, обеспечи-

12
Глава 1
РесЬлекш и сложное поведение
30
вающие более тонкий контроль сокращения мышц, участвующих
как в поддержании позы, так и в движениях. Однако данная упро-
щенная картина должна проиллюстрировать реальность контроля
с помощью обратной связи в регуляции рефлексов (более подробно
см. Хайнд [217], гл. 3).
Гораздо труднее идентифицировать нейроны или рецепторы,
участвующие в регуляции потребления воды или пищи, хотя это
очень важно сделать для анализа поведения. Принцип обратной
связи ясно проявляется в пищевом поведении большинства живот-
ных, которые не едят непрерывно (как это делают некоторые бес-
позвоночные, фильтрующие воду). В гл. 4 мы более подробно рас-
смотрим вопрос о том, что в мозгу млекопитающих есть совершенно
определенные отделы, которые можно отнести к системе регуляции
пищевого поведения. Обратимся снова к рис. 1.8, J5, на котором
выходным сигналом можно считать адекватный уровень питатель-
ных веществ в крови. При голоде он снижается, и это нарушение
контролируется клетками гипоталамуса (гл. 4), которые активиру-
ют нервные механизмы, управляющие питанием. Эти механизмы
перестают работать, когда уровень сахара в крови возрастает, или
даже раньше, когда другие сигналы, например от наполненного
желудка, информируют, что еда должна быть окончена. Рис. 4.5
и его обсуждение на с. 143—144 относятся к модели мотивации,
которая включает этот вид контроля с помощью обратной связи.
Принцип обратной связи несомненно помогает нам понять, как
организовано поведение различных типов. Как мы только что ви-
дели, он приложим к рефлексам и сложным формам поведения от-
дельных особей. До некоторой степени он может быть применен
также и к группам животных. Например, мы можем рассматривать
способ реакции животных на так называемые факторы плотности
популяции как форму регуляции с помощью обратной связи. Когда
размер популяции превышает нормальный уровень, число столкно-
вений между животными увеличивается, пищи становится меньше,
а болезни распространяются быстрее — эффективность комбина-
ции этих и некоторых других факторов будет возрастать с повыше-'
нием плотности популяции. В ответ на эти изменения ухудшается
размножение животных, увеличивается эмиграция и т. д., и в ко-
нечном счете размер, популяции возвращается на прежний уровень.
Поскольку принцип обратной связи имеет чрезвычайно широкое
применение, мы будем возвращаться к нему еще не раз в различных
частях этой книги, например при обсуждении развития пения
птиц (гл. 2). В некоторых книгах по физиологии или поведению,
рассчитанных на достаточно осведомленного читателя, он рассмо-
трен более подробно, а Мак-Фар ланд [315] сделал полный обзор
вопросов, касающихся этого принципа.
Л
В этой главе мы пытались показать, что рефлексы и сложные
формы поведения имеют много общих свойств. Как мы уже указы-
вали, сложные комплексы поведения можно расчленить на более
мелкие элементы, часть из которых непосредственно приравнивается
к рефлексам. Поведение часто бывает организовано иерархически;
мы можем, например, рассматривать простые акты поведения, та-
кие, как ходьба, стояние, кусание или глотание как подчиненные
более высоким системам контроля, которые обозначают как нападе-
ние, бегство, питание и т. д. Иногда рефлексы низшего уровня могут
быть специфичны для единственного вида контроля более высокого
уровня — различные рефлексы, с помощью которых насекомое
чистит туловище, ножки, вытирает глаза, антенны и т. д., в норме
проявляются только в периоды умывания. Но обычно одни и те же
рефлексы должны быть «пригодны» для нескольких видов сложного
поведения. Например, рефлексы, которые принимают участие в по-
лете и ходьбе, должны быть использованы при миграции насеко-
мых, при поиске пищи, при размножении и т. д. Таким образом,
сложные системы регуляции поведения должны конкурировать
между собой за контроль над рефлексами, подобно тому как опи-
санные Шеррингтоном рефлексы конкурируют за управление мыш-
цами.
Конечно, объяснить наблюдаемые нами акты поведения в тер-
минах рефлекторной теории можно далеко не всегда. Такая попытка
часто бессмысленна, потому что существуют разные уровни слож-
ности и поведение необходимо анализировать в его собственных
терминах, а не просто с позиций нейрофизиологии. Тем не менее,
если при изучении поведения физиологические процессы, лежащие
в его основе, окажутся вне поля зрения исследователя, то это
будет большим упущением, потому что между различными уровнями
анализа существуют непрерывные переходы. Даже если оставить фи-
зиологию на заднем плане, всегда необходимо иметь в виду не-
прерывную активность нервной системы. Она не остается дремлю-
щей в ожидании стимулов, которые побудили бы ее к ответам. Ор-
ганы чувств и проприорецепторы в мышцах обеспечивают непре-
рывный фоновый поток импульсов, уровень которого зависит как
от внешних воздействий, так и от эфферентной импульсации мозга.
Информация из внешнего мира и от мышц объединяется в нервной
системе с информацией о различных аспектах обмена веществ в
организме, в том числе о температуре, концентрации углекислоты
и водном балансе. Любое отклонение от оптимального уровня бы-
стро корректируется, и большинство проявлений поведения, ко-
торые мы наблюдаем как результат всей этой нервной активности,
имеет непосредственное приспособительное значение для выживания
животного. Как вы видели, поведение может действовать как чув-
ствительный и мощный механизм поддержания гомеостаза.

р
ГЛАВА 2
РАЗВИТИЕ ПОВЕДЕНИЯ
Раввитие поведения
33
Почти все формы поведения, которые мы наблюдаем у животных,
имеют приспособительное значение. Животные адекватно отвечают
на соответствующие стимулы и благодаря этому обеспечивают себя
пищей, находят убежище, размножаются, заботятся о потомстве.
Их действия, конечно, не безошибочны, но зачастую причиной оши-
бок служит неестественность условий, в которые животные были
перенесены. Так, например, когда птицы впервые попадают в клет-
ку, они делают тщетные попытки вырваться из нее.
ИНСТИНКТ И ОБУЧЕНИЕ
Каким образом животные так хорошо приспосабливаются к сво-
ей обычной среде обитания? Возможно, это обусловлено адекватны-
ми реакциями, запрограммированными в нервной системе животного
при рождении как часть его наследственной конституции. Пчелы,
например, наследуют способность иметь крылья и мышцы для полета,
а также способность подлетать к цветкам за пыльцой и нектаром.
Такие реакции принято называть «инстинктивными» — термин, ко-
торый часто критиковали, но который остается полезным. Инстинк-
тивное поведение развивается постепенно, как и морфологические
признаки, и естественный отбор изменяет его, чтобы лучше приспо-
собить животное к среде обитания. Инстинкт представляет собой
своего рода «видовую память», передаваемую от поколения к поко-
лению по наследству.
Возможен и альтернативный путь приспособления к среде оби-
тания: изменение животным своего поведения в процессе обучения
и накопления опыта. Животные усваивают, какие ответы дают наи-
лучшие результаты, и в соответствии с этим изменяют свое поведе-
ние.
Как инстинкт, так и обучение обеспечивают адаптивность пове-
дения за счет отбора, производимого в первом случае на протя-
жении истории вида, а во втором — на протяжении истории особи;
Однако такое четкое дихотомическое деление поведения оказыва-
ется невозможным, когда дело доходит до реальных примеров. Но
прежде чем переходить к этому, рассмотрим сначала общее значе-
ние инстинкта и обучения для животного царства.
БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ ИНСТИНКТА И ОБУЧЕНИЯ
Инстинкт наделяет животное набором адаптивных реакций, ко-
торые оказываются «готовыми» к проявлению при первой же необ-
ходимости в них. Это создает явные преимущества для животных
с коротким сроком жизни или лишенных родительской заботы.
Членистоногие, например, характеризуются замечательным разви-
тием инстинкта, потому что другие способы поведения для них не-
возможны. Самка роющей осы появляется из своей подземной ку-
колки весной. Ее родители умерли прошлым летом. Она должна
встретиться с самцом и затем выполнить сложную серию действий,
связанных с рытьем гнездовой норки, постройкой в ней камер,
охотой и умерщвлением добычи, такой, например, как гусеница.
Далее, она должна снабдить каждую камеру добычей, отложить
яйца и, наконец, «запечатать» ячейки. Все это нужно сделать в те-
чение нескольких недель, после чего она погибает. Совершенно не-
постижимо, как она могла бы выполнить всю эту плотную програм-
му, если бы должна была учиться всему от случая к случаю методом
проб и ошибок.
Ситуация с роющей осой противоположна той, в которой оказы-
вается львенок. Он рождается совершенно беспомощным, и мать обе-
регает и выкармливает его до тех пор, пока он сможет сам передви-
гаться. Постепенно он переходит на твердую пищу, приобретает
ловкость в игре с другими львятами. Он имеет постоянную возмож-
ность наблюдать родителей и других сородичей и подражать им
в преследовании и ловле добычи. Впервые поймать живую добычу
львенок может в возрасте 6 мес, но пройдет не менее 2 лет, пока он
вырастет достаточно, чтобы питаться самостоятельно. Его поведение,
и особенно методы и уловки, используемые им во время охоты,
могут изменяться в соответствии с обстоятельствами на протяжении
всей его жизни.
Роющая оса, для которой характерно предустановленное ин-
стинктивное поведение, и лев, который относительно свободно обу-
чается, представляют две крайние точки на шкале поведенческих
актов. На самом деле в приведенных описаниях действительный ха-
рактер развития поведения этих животных чрезвычайно упрощен.
Роющая оса за свою короткую жизнь может и должна обучиться
многому, например запомнить точное расположение каждой из
своих норок, потому что она должна возвращаться к ним после вы-
ходов на охоту. Молодой лев обладает некоторыми хищническими
наклонностями, которые несомненно инстинктивны, хотя лев должен
учиться, как их применять.
У всех животных более высокого уровня развития, чем кольча-
тые черви, обнаруживаются оба этих вида поведения, и каждый имеет
свои собственные особые преимущества. Это положение хорошо
иллюстрируется примерами в исследованиях криков птиц. У птиц
I
2 № 147
68
Глава 2
часто обнаруживается сильное развитие как инстинктов, так и спо-
собности к обучению. Как мы рассмотрим далее, для того чтобы песня
самца окончательно сформировалась, ему необходимо не только
петь самому, но и слушать других самцов. Что же касается криков
тревоги, то оказалось, что у всех изученных видов они полностью
воспринимаются и воспроизводятся с первого же раза. Естественный
отбор благоприятствовал врожденным реакциям там, где задержка
на обучение могла бы оказаться фатальной. Такие контрасты на-
водят на мысль, что сам по себе тип развития может эволюциониро-
вать путем отбора (с. 73—74).
Одно из преимуществ обучения перед инстинктом — его большая
потенциальная способность изменять поведение при столкновении
с изменившимися обстоятельствами. Оно несомненно более важно
для животных с долгим сроком жизни, чем для насекомых, которые
живут лишь несколько недель. Другим существенным фактором
может быть размер тела, так как высокоразвитая способность к
обучению сопряжена с относительно большим количеством мозговой
ткани, что невозможно для очень маленьких животных. Обычно
размеры тела и срок жизни в некоторых пределах положительно кор-
релируют: крупные животные живут дольше, чем мелкие. Не говоря
о физических факторах, ясно, что благодаря естественному отбору
возможны разные степени способности к обучению в соответствии
с историей вида. Два наиболее развитых отряда насекомых, пере-
пончатокрылые (муравьи, пчелы и осы) и двукрылые, сопоставимы
по размерам и продолжительности жизни. Для перепончатокрылых
в дополнение к их богатому набору инстинктивных действий харак-
терна чрезвычайная легкость обучения, хотя и специализирован-
ного типа, и это играет важную роль в их жизни. В течение коротких
трех недель, когда рабочая пчела летает за взятком, она запоминает
точное расположение своего улья и цветущих культур, на которых
кормится. В течение дня она может перелетать от одних культур
к другим, потому что запоминает, в какое время на какой из них
выделяется больше всего нектара. После трех посещений кормушки,
помеченной определенным цветом, пчела-сборщица запоминает этот
цвет на весь оставшийся период жизни. Даже после единственного
посещения кормушки пчела реагирует на соответствующий цвет,
и лишь через 5—6 дней ее реакции возвращаются к среднему уровню
[342].
Двукрылые тоже могут обучаться. Журчалки, например, запо-
минают положение цветов, которые они посещают, а комнатные мухи
стремятся садиться в комнате на одни и те же места. Доказано
также, что с помощью специальных приемов можно выработать у
дрозофилы некоторые простые дифференцировки. Но у большей
части двукрылых память коротка, а возможности обучения очень
ограниченны. В отличие от перепончатокрылых они не строят по-
стоянных гнезд, к которым должны были бы возвращаться регуляр-
Развитие поведения
57
но. Для их короткой жизни им вполне достаточно наследственно
обусловленных реакций на стимулы, которые сигнализируют о
присутствии пищи, убежища и партнера.
ХАРАКТЕРИСТИКИ ИНСТИНКТА И ОБУЧЕНИЯ
Рассмотрим более подробно характеристики инстинкта и обу-
чения. Две особенности инстинктивных действий на первый взгляд
могут показаться уникальными. Во-первых, эти действия представ-
ляют собой закрепленные стереотипные комплексы движений, очень
сходные у всех особей данного вида. Например, все роющие осы
одного вида строят свои гнезда одинаковым способом; все домаш-
ние петухи совершают одну и ту же серию движений, когда ухажива-
ют за курами, и т. д. Во-вторых, инстинктивные действия часто
Легко вызываются очень простыми стимулами. Сталкиваясь со слож-
ной ситуацией, животное реагирует на одну ее часть и фактически
игнорирует остальные. Зарянка, например, гораздо агрессивнее
при виде пучка красных перьев из грудки самца-соперника, чем
при виде птицы, у которой отсутствуют только эти перья.
Поразительно, однако, что эти характеристики совершенно не
годятся для того, чтобы отличать инстинкт от обучения. Послед-
нее часто описывается довольно неопределенно как «более лабиль-
ное» поведение, но на деле набор связанных с ним движений может
быть столь же стереотипным, как и при инстинкте. Крысы, поме-
щенные в камеру, где они должны научиться нажимать на выступаю-
щий из стенки камеры рычаг, чтобы получить пищевую таблетку
(рис. 2.1), применяют для этого разные способы. Некоторые ис-
пользуют всегда левую лапу, другие нажимают подбородком, причем
обычно крысы проявляют консерватизм в использовании избранного
ими метода. В своей очаровательной книге «Кольцо царя Соломона»
Лоренц [306] описывает в интереснейших деталях, как куторы изу-
чают географию окрестностей гнезда. Если в каком-то месте своего
пути они должны перепрыгивать через небольшой брусок, они зау-
чивают это движение так прочно, что продолжают совершать пры-
жок точно таким же образом даже после того, как препятствие
устранено. Есть масса других примеров, когда животные испыты-
вают сходные трудности, если им нужно разучиться делать что-то.
Оказывается, движения после обучения становятся почти «автома-
тическими» и могут сохраняться, даже если перестают быть эффек-
тивными. Выученные двигательные акты почти всегда вызываются
какими-то специфическими признаками окружающей среды, тогда
как, так же как и в случае инстинктов, другие ее признаки могут
игнорироваться. В сложной меняющейся обстановке животное от-
носительно легко обучается зыделять один ключевой стимул.
Если мы хотим выявить какое-то кардинальное различие между
инстинктивным и приобретенным поведением, то должны основы-
2*

П'1
60
Г лава 2
Рис. 2.1. Крыса в скиннеровской камере. При нажиме на рычаг в кормушке появ-
ляются маленькие пищевые таблетки.
I ;
ваться не на их xapaKTepncTHKax
v
а на способе развития того и
другого у данной особи. Сопоставим поведение крысы при обучении
нажатию рычага для получения пищи с инстинктивным поведением
при спаривании, Каждая крыса нажимает на рычаг стереотипным
способом, но эти способы различны у разных крыс. В отличие от
этого готовые к размножению самки, когда самец делает садку,
принимают одну и ту же позу подставления (рис. 2.2). Далее, если
мы поместим голодную крысу в камеру с рычагом, то могут пройти
часы, прежде чем она случайно толкнет его. Но если девственную
самку подготовить к размножению путем гормональных инъекций,
то она примет позу подставления после короткого опыта общения с
самцом.
Основой критерия инстинктивного поведения является способ
его развития, а наиболее общий метод тестирования — так называе-
мый опыт с изоляцией, Животных содержат индивидуально вне кон-
Развитие поведения 37
Рис. 2.2. Спаривание у крыс [28]. Поза самки с поднятым газом и отогнутым хво-
стом очень стереотипна. Давление передних лап на ее .бока — один из стимулов,
необходимых для того, чтобы самка реагировала.
такта с сородичами со столь раннего возраста, насколько это воз-
можно. Когда живбтное вырастает, проверяют его реакции на
разные стимулы и сравнивают их с реакциями нормально выращен-
ных животных. В действительно строгих условиях изоляции было
исследовано довольно мало животных, однако показано, что неко-
торые рыбы и птицы совершенно нормально выполняют многие ком-
плексы реакций полового, пищевого и ориентировочного поведения
после выращивания в изоляции. Жизненные циклы многих насеко-
мых представляют собой естественные опыты с изоляцией. Они ярко
свидетельствуют о том, что в формировании поведения взрослых
насекомых ни тренировка, ни обучение не играют роли. Хотя та-
кие опыты бесспорно позволяют нам исключить возможность того,
что животные учатся, как сделать что-то, они дают лишь очень
ограниченное представление о факторах, которые могут действовать
в период развития,— обучение и тренировка только две из большого
набора возможностей.
Дело в том, что в опытах с изоляцией можно показать только,
какие факторы не важны для развития поведения, но при этом ни-
чего нельзя сказать о том, какие факторы в нем участвуют. Напри-
мер, молодые кряквы, выведенные в инкубаторе, первое время реа-
гируют предпочтительно на крики крякв, а не других близких им
видов. Они явно инстинктивно предпочитают крики своих сороди-
чей. Однако Готтлиб [171, 172] показал, что один из факторов,
участвующих в развитии этого предпочтения,— это способность
утиных эмбрионов воспринимать звуки, совершенно не похожие
, на крики матери, которые они сами и другие утята производят,
пока еще находятся в скорлупе.
Нет сомнения в том, что развитие многих форм поведения должно
быть генетически обусловлено и наследственно детерминировано

38
Глава 2
Развитие поведения
39
потенциями центральной нервной системы. Это положение необхо-
димо иметь в виду при изучении развития поведения. Разумеется,
мы не должны довольствоваться лишь отнесением таких форм по-
ведения к «инстинктивным». Гены могут контролировать развитие
поведения, но для этого они должны взаимодействовать с окружаю-
щей животное средой. Утята в опытах Готтлиба, прежде чем начать
реагировать на призывы матери, нуждались в некоторой звуковой
стимуляции. Наша задача будет заключаться в том, чтобы найти те
факторы в окружении животного, которые влияют на развитие его
инстинктивного поведения. Очевидно, мы должны избегать любой
простой дихотомии, которая приписывает инстинкт действию генов,
а обучение влиянию среды. В развитии всех форм поведения долж-
ны участвовать оба этих фактора. Казалось бы, нет нужды особо
подчеркивать это очевидное положение, но в действительности до
совсем недавнего времени именно непонимание этого положения
служило причиной больших разногласий среди исследователей пове-
дения животных.
ЭТОЛОГИ и психологи
После второй мировой войны на Западе сложились две основные
школы исследователей поведения животных. Первую составляли в
основном американские специалисты по экспериментальной пси-
хологии. Их родоначальником был Уотсон [488], чья книга «Бихе-
виоризм», опубликованная в 1924 г., явилась поворотным пунктом
в развитии современного экспериментального подхода к изучению
поведения. Вторая школа — преимущественно европейская — была
основана в конце 30-х годов Конрадом Лоренцем, австрийским ис-
следователем, работавшим в Германии. Он был связан с Нико Тин-
бергеном (Голландия), с которым сотрудничал еще в ранних рабо-
тах. После окончания войны Лоренц и Тинберген организовали
исследовате 1ьские группы, куда вошли различные ученые, большей
частью зоологи. Они называли себя «этологами» и этологию опреде-
ляли просто как «научное изучение поведения».
Этологи и американские психологи подходили к изучению пове-
дения с противоположных сторон. Этологи изучали широкий круг
видов животных в условиях, близких к естественным, часто в поле
с использованием укрытий, Они находились под впечатлением бо-
гатого разнообразия форм адаптивного поведения у насекомых,
рыб и птиц и сосредоточились на их изучении. В основном они изу-
чали поведение животных в период размножения, поскольку за ним
удобно наблюдать. Кроме того, этологи часто достигали больших
успехов в содержании животных в неволе и добивались их размно-
жения.
Первоначальный подход этологов описан Тинбергеном [460] в.
его книге «Изучение инстинкта». Позднее он развил свои идеи в
замечательном обзоре [466], где рассматриваются цели этологии.
В основном эти идеи касаются способов, с помощью которых по-
ведение животных обеспечивает их выживание. Этологи изучили
стимулы, которые вызывают инстинктивное поведение, и мотива-
цию, которая его контролирует, и впоследствии часто изучали в
деталях развитие такого поведения. Центральной проблемой для
них всегда была функция и эволюция поведения. Этологи изучают
эволюцию поведения путем сопоставления поведения близкородст-
венных видов и устанавливают происхождение отдельных поведен-
ческих актов подобно тому, как сравнительные анатомы делают
это в отношении признаков строения. Исследователи, занимающиеся
этологией, не очень интересуются проблемами, связанными с обу-
чением, и хотя они признают, что животные способны к обучению,
обычно они не рассматривают этот факт или исключают его из
своих теорий и экспериментов.
Вместе с тем экспериментальные психологи интересуются почти
исключительно обучением. Они в значительной степени пренебрега-
ют поведением животных в естественных условиях и намеренно
ограничивают их в условиях эксперимента. Поведение животного
в лабиринте и в «челночной камере», в которой его учат переходить
из одной половины камеры в другую, чтобы избежать электрического
раздражения, в «скиннеровской камере», в которой его учат нажи-
мать на рычаг или клевать ключ для получения пищи или питья
в качестве вознаграждения,— таковы ситуации, которые интересу-
ют психологов. Их цель заключается, во-первых, в составлении «за-
конов поведения», которые описали бы, как поведение изменяется
при данном уровне тренировки, подкрепления или наказания, и,
во-вторых, в составлении прогнозов относительно, например, той
или другой экспериментальной ситуации для осуществления обу-
чения.
Неудивительно, что экспериментальные психологи использова-
ли более ограниченный набор животных, чем этологи. Самый лучший
объект в исследованиях по обучению — млекопитающие, которых
психологи преимущественно и использовали, в основном отдавая
предпочтение белым лабораторным крысам.
Сначала обе школы мало замечали работы друг друга, но через
несколько лет возник перекрестный огонь и каждая старалась за-
нять довольно крайние позиции. Работа Лермана [286], например,
может служить крайним вариантом выступления против концепции
этологов того периода
1
. Психологи обвиняли этологов в недооценке
1
Упоминая здесь Лермана, мы как бы причисляем его к лагерю психологов,
но это совершенно неверно. Он имел хорошее биологическое образование и, хотя
и выступал с критикой многих положений ранней этологической теории, симпати-
зировал их подходу и весьма критически относился к большей части современной
экспериментальной психологии. Описание работ Лермана, имевших в этом плане
большое влияние, можно найти у Бира [44].