Меннинг О. Поведение животных. Вводный курс
Подождите немного. Документ загружается.

J
Глава 3
В то же время хотя лысухи имеют ярко-белый клюв, их молодые
птицы все же предпочитают красное. Фазанята и утята нескольких
видов клюют сильнее всего зеленое, тогда как этот цвет совершенно
непривлекателен для других исследованных птиц. Возможно, что
предпочтение зеленого может быть просто реакцией на яркость,
потому что глаза уток и фазанов имеют повышенную чувствитель-
ность к зеленой части спектра. Кир [263] обнаружила, что при
варьировании яркости цветовых пятен относительная популяр-
ность некоторых цветов изменяется. Однако кажется наиболее
вероятным, что утята выбирают зеленое именно по цвету. Возмож-
но, это связано с тем, что фазанята и утята должны кормиться сами
как только покинут гнезда, причем едят они преимущественно раз-
нообразные зеленые растения.
Оппенгейм [374] недавно подробно изучил предпочтение цвета
у утят кряквы. Подобно Кир, он обнаружил сильное предпочтение
зеленого, вероятно, потому, что утят привлекают зеленые предметы,
контрастирующие с фоном. Оппенгейм обнаружил, что исходное
предпочтение зеленого не зависит от того, находились ли яйца в
инкубаторе в полной темноте или же в их скорлупе были проделаны
окошки и глаза эмбрионов освещались белым или желтым светом
в течение многих часов перед вылуплением. Оппенгейм не смог
отнести предпочтение зеленого цвета только за счет свойств сетчатки
глаза уток.
Все эти наблюдения приводят к предположению, что в некото-
рых случаях предпочтение цвета имеет наследственную основу,
которой обладает центральная нервная система. В следующем при-
мере мы рассмотрим избирательную реактивность на другие аспек-
ты зрительного стимула — форму и движение.
Реакция тревоги при виде летящего хищника
Многие гнездящиеся на земле птицы, такие, как утки, гуси,
фазаны и индейки, издают крик тревоги и затаиваются, когда над
ними пролетает хищная птица. Эта реакция особенно выражена у
самок с выводком. Индейки при этом расправляют хвосты, и ин-
дюшата бегут прятаться под них.
Тинберген [460] описывает некоторые опыты, поставленные сов-
местно с Лоренцем в 1937 г. Они изготовили множество простых
картонных силуэтов летящих птиц и передвигали их по проволоке
над загоном с гусями и различной дичью.
Регистрируя наличие и силу реакции тревоги, они обнаружили,
что наиболее действенны все модели с «короткой шеей», характер-
ной для большинства хищных птиц. В наиболее поразительном эк-
сперименте, ставшем классическим, несмотря на его противоречи-
вость, использовали силуэт, показанный на рис. 3.10. Он не вызы-
вал беспокойства, если летел влево, длинной шеей вперед (изобра-
Стимулы и коммуникация
99
-fcu. "Хищник"
"Гусь~
Рис. 3.10. Силуэт «гусь — хищник», использованный при изучении реакции тре-
воги у гусей и других птиц [460].
жая гуся), и был очень эффективен, если двигался короткой шеей
вперед (изображая ястреба). В данной ситуации действенна не про-
сто форма знакового стимула, но ее соотношение с движением. Ма-
ловероятно, чтобы такая избирательная реактивность могла быть
обусловлена только свойствами сетчатки птиц.
Многие критики скептически отнеслись к предположению, что
ответ на стимул столь сложной конфигурации может быть наслед-
ственно обусловленным. Шнейрла [424] предположил, что существу-
ет гораздо более общее объяснение для таких реакций. Он считает,
что все животные выше простейших имеют тенденцию приближаться
к любым источникам умеренной стимуляции —звуковым, световым
или химическим — и избегать сильной и резко усиливающейся
стимуляции. Он предположил, что силуэт, движущийся как ястреб,
вызывает большее беспокойство просто потому, что имеет широкий
передний край. Благодаря этому происходит более резкое затем-
нение поля зрения птицы, чем в случае силуэта гуся, который рас-
ширяется постепенно от узкого и длинного ведущего конца. Шнейр-
ла относится скептически к любому ВРМ, который специально
приспособлен к силуэту ястреба, и предсказывает, что тот же са-
мый дифференцированный эффект можно получить при помощи
обыкновенного треугольника в зависимости от того, каким спосо-
бом его двигать. Шнейрла предполагает, что движение основанием
вперед вызовет у птиц больше беспокойства, чем движение верши-
ной вперед.
Несомненно, мы должны критически отнестись к любому ут-
верждению о том, что обнаружена наследственно обусловленная
специфическая реактивность на сложные стимулы. Поэтому впол-
не понятно, что исходные эксперименты подвергаются множеству
критических замечаний. Прежде всего утверждать что-либо опре-
4*

100
Глава 3
Стимулы и коммуникация
101
деленное в отношении природы наследственной реакции на форму
ястреба можно только в том случае, если в эксперименте использу-
ются птицы, совершенно не имеющие соответствующего опыта.
Птицы, использованные Лоренцем и Тинбергеном, могли до эк-
спериментов узнать, как выглядят ястребы. К тому же эти иссле-
дователи учитывали только реакцию группы, а отдельные особи
могут реагировать по-разному, причем реакция одной особи зависит
от поведения других.
Первая попытка применить более строгие критерии была сдела-
на Хиршем, Линдли и Толменом [226], которые работали с только
что вылупившимися белыми леггорнами, оценивая их реакции ин-
дивидуально. Они обнаружили, что модель ястреба вызывает не
большее беспокойство, чем модель гуся. Когда обе модели впервые
появлялись над цыплятами, они вызывали умеренное беспокойство,
затем цыплята быстро привыкали к ним и почти не обращали на них
внимания.
Тинберген [464] и другие исследователи совершенно резонно
полагают, что нельзя обобщать данные по белым леггорнам и диким
гусям. Первые могли утратить наследственно обусловленную ре-
акцию на силуэт ястреба за многие поколения одомашнивания в
отсутствие соответствующего отбора. Позднее Мельзак и др. [340]
повторили эксперименты этого типа с утятами дикой кряквы. Им
удалось до некоторой степени подтвердить первоначальные данные
Лоренца и Тинбергена. Они показали, что когда над утятами впер-
вые «пролетает» модель ястреба, реакции страха подвергается при-
близительно вдвое большее число не имеющих опыта утят, чем в
случае, когда пролетает модель гуся. Однако, подобно Хиршу с со-
авторами, они нашли, что утята быстро перестают бояться обеих
моделей, хотя они никогда не ослабляют внимания и всегда следят
глазами за пролетающей над ними моделью. Грин и др. [176, 177]
получили те же результаты — «ястреб» вызывает гораздо больше
беспокойства, чем «гусь» у утят, которые до этого не видели над со-
бой никаких предметов. Кроме того, они проверили предположе-
ние Шнейрлы относительно простых треугольных силуэтов и не
смогли подтвердить его. Утята почти не проявляли беспокойства
при виде треугольника, в каком бы направлении он ни двигался.
Работа Шлейдта [420] на индейках внесла в этот вопрос допол-
нительные осложнения. В ней было показано, что реакция тревоги
может быть вызвана при помощи модели почти любой формы —
например простого круга,— если он летит с соответствующей ско-
ростью. Эта скорость довольно мала — от 5 до 10 длин модели в
секунду — и хорошо совпадает с видимой скоростью полета хищ-
ной птицы. Даже крошечная модель всего в дюйм (2,54 см) длиной
вызовет реакцию тревоги, если она движется с этой скоростью.
Шлейдт обнаружил, что индейки быстро прекращают реагировать
на любые модели, с которыми они уже ознакомились, но новые
формы, независимо от того, как они двигались, обычно пугают их.
Он предполагает, что гуси в опытах Лоренца и Тинбергена не реа-
гировали на модель с длинной шеей потому, что они уже привыкли
к виду летящих над ними гусей, однако это не может объяснить
результатов, полученных Мельзаком и Грином.
Гораздо труднее соотнести результаты описанных эксперимен-
тов с естественной ситуацией. Птицы перестают отвечать на зна-
комые модели независимо от того, насколько они похожи на ястреба,
однако в естественных условиях они продолжают реагировать на
хищных птиц, хотя мало кто из них подвергается нападению. Воз-
можно, это связано с тем, что модели всегда появляются в одном и
том же месте и движутся одним и тем же путем. Кроме того, как
указывает Шлейдт, дикие птицы редко встречаются с летящими
ястребами со столь высокой частотой, какая применяется в опытах
с моделями. Редкое появление ястребов должно быть важным фак-
тором в поддержании реактивности.
Итак, существуют доказательства, что дикие птицы имеют ВРМ,
который позволяет им реагировать на хищных птиц при первой
же встрече с ними. Этот ВРМ, вероятно, имеет у разных видов раз-
ные свойства, но ясно, что к ним относится короткая шея и соот-
ветствующая скорость движения. Маловероятно, чтобы его действие
было связано с периферической фильтрацией.
Любая попытка проанализировать свойства ВРМ сталкивается
с необходимостью привлечения данных как по физиологии органов
чувств, так и по поведению. Ясно, что ВРМ, если он имеет какое-то
применение как понятие, должен включать целый ряд возможных
фильтров, начиная с самих органов чувств. Конечным результатом
процесса фильтрации должно быть приспосабливание животного
к избирательному ответу на некоторые знаковые стимулы окружаю-
щей среды. Эти стимулы, возможно, являются специально развив-
шимися ключевыми стимулами, чьи свойства приспособлены к.свой-
ствам ВРМ. Основная нерешенная проблема касается природы цен-
тральных фильтров. Маловероятно, чтобы они были связаны с ме-
ханизмом какого-то простого типа, хотя свойства центральных филь-
тров, которые опосредуют дифференцировочное обучение, и тех,
с которыми мы имеем дело в данном случае (т. е. фильтров с генети-
чески контролируемым развитием), возможно, имеют много общего.,
Опыты психологов, изучавших, как животное опознает предметы,
показывают, что оно реагирует не на все признаки одновременно.
Скорее оно направляет то, что можно назвать центральным «меха-
низмом внимания», сначала на один вид признаков, скажем яркость,
цвет или форму, а затем на другие [168]. Очевидно, такой механизм
внимания имеет много общих свойств с центральными фильтрами,
о которых мы говорили. Этологи были поражены тем, как измене-
ние «настроения» животного вызывает переключение внимания;
один фильтр затормаживается, а другой приводится в действие.

88
Глава 3
Стимулы и коммуникация 119
53
:!
I •
Сразу после освобождения из куколки самец бабочки-сатира силь-
нее реагирует на желтое и синее —признаки, по которым он находит
цветки. Позднее, в период ухаживания за самкой, с помощью тех
же органов чувств он отвечает преимущественно на коричневое.
Когда «настроение» самца меняется и он переходит от кормления
к ухаживанию [460], каким-то образом осуществляется смена филь-
тров.
КОММУНИКАЦИЯ
Как мы говорили в этой главе, многие из наиболее важных внеш-
них стимулов, на которые реагируют животные, исходят от других
животных — сигналы от хищников или от добычи, сигналы от ро-
дителей, на которые отвечает молодняк, сигналы ухаживания от
одного полового партнера к другому и т. д. Имеется много доказа-
тельств, что у животных часто развиваются особые структуры —
релизеры
1
— и особые формы поведения, основная функция кото-
рых состоит в передаче сигналов другому животному. Другими
словами, животные общаются друг с другом, и нам придется обра-
щаться к разным аспектам этого процесса на протяжении всей
книги. Более подробное рассмотрение этой темы можно найти у
Хайнда [218, 219] и Себеока [429].
Прежде всего определим, что мы будем иметь в виду под словом
«коммуникация», а это не совсем простая задача. Подобно многим
словам, используемым и в технике, и в обычной речи, благодаря
постоянному употреблению оно вобрало в себя несколько различных
значений. Коммуникация всегда включает передачу информации,
и в наиболее широком определении это должно учитываться, т. е.
коммуникацию можно определить как «любую передачу информа-
ции». Если принять это определение, то можно сказать, что разные
части вычислительной машины вступают в общение друг с другом,
а вся машина — с человеком-оператором. Однако, если мы использу-
ем столь широкое определение, нам не удастся выявить различия,
которые очень важны при изучении биологических объектов. Так,
если самец зарянки видит другого самца, вторгшегося на его тер-
риторию, и летит, чтобы сесть на соседнюю ветку, мы должны за-
ключить, что и соперник, и ветка вступают в общение с хозяином
территории.
Большинство биологов предпочли бы ограничить значение этого
термина передачей информации между животными, но при этом
все же остается ряд неясностей. Должны ли мы ограничить опреде-
1
В русской литературе термин «релизер» часто используется для обозначения
любых ключевых раздражителей. В данном случае автор использует его для обоз-
начения морфологических структур, выполняющих сигнальные функции.—
Прим. ред.
ление коммуникации передачей информации в пределах вида?
По-видимому, должны, так как в противном случае мы вынуждены
были бы заключить, что антилопа-гну общается со львом, который
устраивает на нее облаву, а лев общается с антилопой-гну, нападая
на нее и обращая ее в бегство. Бургхардт [79] в своем очень полез-
ном очерке проблем определения термина «коммуникация» приводит
другой пример, который делает эту точку зрения еще более убеди-
тельной. Муравьи-фуражиры обычно оставляют пахучие следы,
которые улавливают их товарищи по гнезду и следуют по ним к ис-
точнику пищи. Мелкие змеи (Leptotyphlops) также обнаруживают
эти следы и движутся по ним в противоположном направлении
к муравейнику, где уничтожают яйца и личинок. Вероятно, в
данном случае запах следует рассматривать как сигнал, развив-
шийся у муравьев для общения друг с другом, но не со змеями.
Мы подошли к ограничению нашего определения, которое свя-
зано с использованием понятия «намерения», или цели (intent).
Это понятие отражает тот факт, что отправитель и получатель сиг-
нала взаимно приспособлены друг к другу. Нужно согласиться,
что и само слово «намерение» нелегко определить строго, но, если
его применять с осторожностью, оно оказывается полезным. Ясно,
что намерение передать информацию часто бывает не осознанным
действием, а скорее определяет взаимоотношения между вступаю-
щими в общение животными, которые установились к их взаимной
пользе. Мы уже обсуждали взаимное приспособление ключевых
стимулов и врожденных разрешающих механизмов, которое было
постулировано этологами (с. 86), и термин «социальный сигнал»,
конечно, подразумевает функцию общения. В примере, который
мы только что описали, муравьи намерены при помощи пахучих
следов общаться друг с другом, но одно из осложнений, возникаю-
щих при этом,— возможность использования передаваемой ими
информации хищными змеями. Мы можем быть уверены, что пре-
имущества, получаемые муравьями при этом виде общения, переве-
шивают его случайные недостатки.
Такой способ использования понятия намерения означает, ко-
нечно, что мы не можем разумно ограничить коммуникацию пере-
дачей информации между особями одного вида. Нектарные указа-
тели цветов (см. рис. 3.2) передают информацию пчелам и это по-
лезно как тем, так и другим. Полосатый скунс передает информа-
цию потенциальным хищникам, демонстрируя заднюю часть туло-
вища и поднимая хвост. Звуки, издаваемые гремучей змеей, служат
той же цели.
Не все исследователи процесса коммуникации у животных поль-
зуются нашим определением. Альтман [8] и Хайнд и Рауелл [221]
изучали коммуникацию в сообществе резусов и попытались описать
их зрительные сигналы. Первый выделил их около 50, последние —
только 22. Причина такого расхождения лежит в разном использо-
88
Глава 3
Стимулы и коммуникация 119
54
I
вании термина «коммуникация». По определению Альтмана, комму-
никация в сообществе представляет собой «процесс, посредством
которого поведение одной особи влияет на поведение других».
Под такое определение подходят все виды поз и движений, например
вид кормящейся обезьяны. Вместе с тем Хайнд и Рауелл используют
определение, сходное с рассмотренным нами выше; они считают
зрительными сигналами только те, которые, по-видимому, специ-
ально развились для цели коммуникации. Даже если в этом кратком
рассмотрении процесса коммуникации мы сконцентрируем внима-
ние на специально развившихся социальных сигналах, важно соз-
навать, что животные на самом деле получают массу информации
от других особей по способу, описанному Альтманом. Мы уже упо-
минали (с. 86) о разнообразии стимулов, "исходящих от доминант-
ных самцов обезьян, на которые реагируют другие члены группы.
Помимо каких-то специальных сигналов, например угрожающих
движений (с. 191), которые он может производить, поступающая
от него информация содержится в положении тела и способе пере-
движения. Почти любой чертой облика и в движении, и в покое
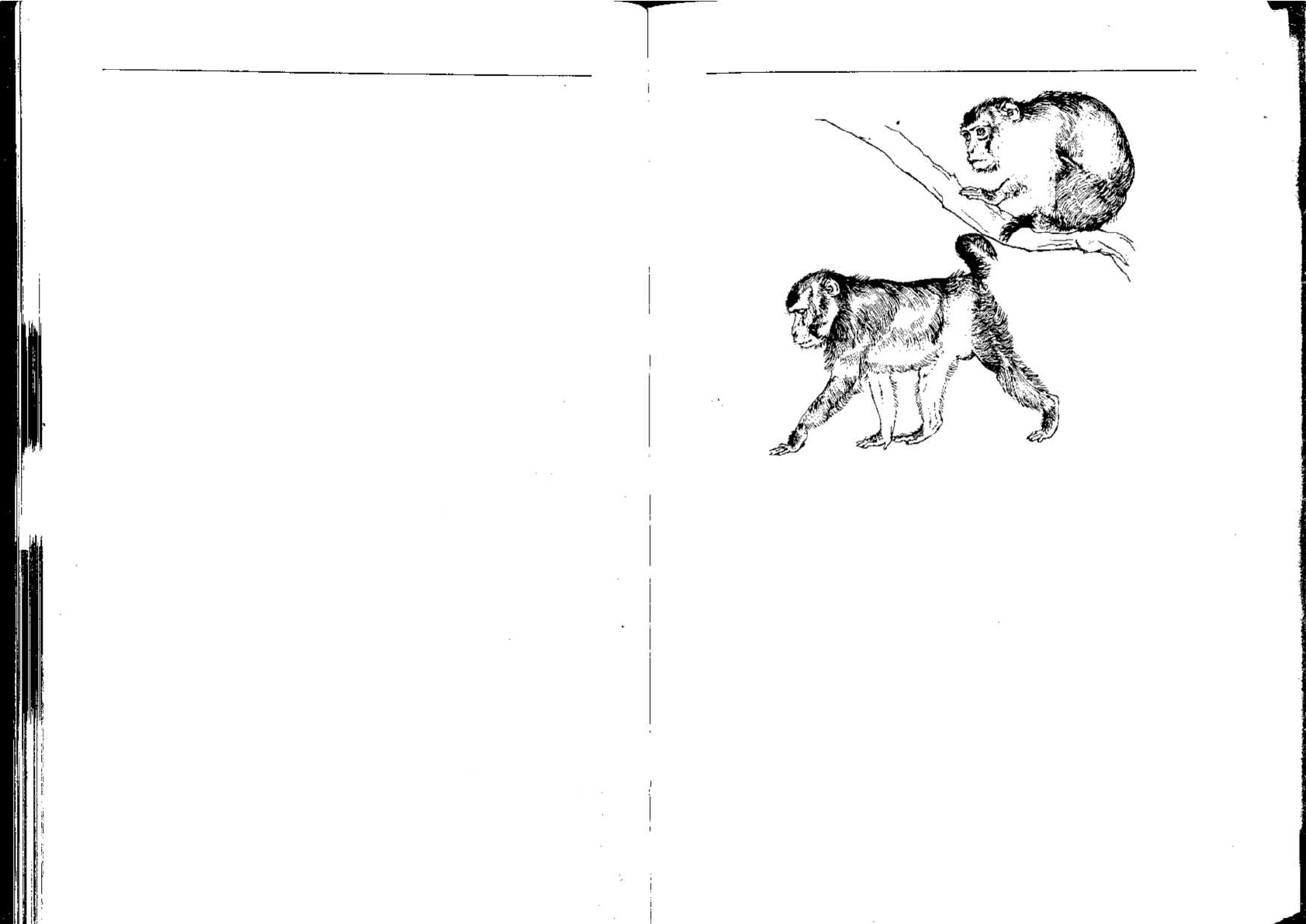
доминантный самец резко отличается от подчиненного (рис. 3.11).
В таких случаях коммуникации может быть трудно решить, уча-
ствует ли здесь намерение вступить в общение или нет; далеко
не всегда можно с уверенностью идентифицировать социальный сиг-
нал, особенно у приматов.
Наше обсуждение проблем коммуникации мы можем свести к
трем взаимосвязанным вопросам:
1. Как социальные сигналы приспособлены к своей роли?
2. Что передается?
3. Как мы можем исследовать, происходит ли коммуникация?
1. Как социальные сигналы приспособлены к своей роли? При
рассмотрении способов общения в царстве животных (прекрасны-
ми обзорами этой темы являются центральные главы у Мэрлера и
Гамильтона [333] и у Брауна [73]) было обнаружено, что различные
группы животных более или менее специализированы по типам ис-
пользуемых сигналов в зависимости от степени развития у них тех
или иных органов чувств. Существует также общая корреляция
между свойствами сигнала и функцией, которую он выполняет.
Осязание ограничено в своих возможностях передачи информа-
ции, но во многих отношениях это главный из каналов коммуника-
ции почти для всех видов живой материи, отвечающих на физиче-
ский контакт. Тактильная коммуникация несомненно доминирует
в общественных взаимодействиях многих беспозвоночных; напри-
мер, в некоторых колониях термитов у слепых рабочих, которые
никогда не покидают своих подземных туннелей, или у дождевых
червей, которые ночью выползают из нор для спаривания. У бес-
позвоночных осязание тесно связано с химической чувствительно-
стью, потому что специализированные тактильные органы, напри-
Рис. 3.11. Типичная поза, принимаемая подчиненным (Л) и доминантным (Б)
макаками-резусами [219].
мер антенны или пальпы насекомых, часто снабжены также хемо-
рецепторами. Общественные насекомые передают большое количе-
ство информации в своих колониях путем комбинации тактильных
и химических сигналов (гл. 8). Тактильная коммуникация сохраняет
свое значение у многих позвоночных, в частности у млекопитаю-
щих, наиболее общественные виды которых проводят значительную
часть времени в физическом контакте друг с другом. Вероятно,
в большинстве таких случаев неверно предполагать наличие спе-
циально развившихся социальных сигналов; по-видимому, так-
тильные раздражения, исходящие от другого члена той же группы
обезьян, передают более общую информацию о взаимоотношениях
между двумя животными, чем специально развившийся социальный
сигнал чистки шерсти (с. 320—321 и рис. 8.9).
Тактильная коммуникация в силу своей природы возможна
только на очень близком расстоянии. Длинные антенны тараканов
и раков действуют как «разведчики», которые позволяют им иссле-
довать мир в радиусе одной длины тела, но это почти предел для
осязания. Другие сенсорные системы — зрение, слух и обоняние —
обеспечивают общение на значительном расстоянии. Звук и запах
имеют дополнительное преимущество благодаря тому, что они спо-
собны преодолевать естественные препятствия, например густую

88
Глава 3
Стимулы и коммуникация 119
107
U
растительность, и даже проходить через нее. Поэтому сигналами,
рассчитанными на большое расстояние, являются обычно крики
или специально издаваемые запахи.
Звуковые сигналы и способы их издавания хорошо приспособ-
лены к их функции. Низкочастотные крики лучше всего прони-
кают через густую растительность; к этому типу сигналов обычно
относятся крики тропических лесных птиц. Хукер и Хукер [231]
описывают негромкие флейтоподобные звуки африканского кустар-
никового сорокопута (Laniarius aethiopicus), самец и самка которого,
скрываясь в густом лесу или кустарнике, поют «дуэтом» по очереди
издавая звуки. Мэрлер [330] указывает, что крики многих прима-
тов, живущих в тропических лесах, также приспособлены к рас-
пространению на большие расстояния. Мы уже обсуждали харак-
теристики идеального крика тревоги и упоминали механизм кон-
вергентной эволюции, в результате которой выработался сигнал,
оптимальный для данной функции. Мы вернемся к этому вопросу при
обсуждении эволюции в гл. 6.
Распространение звукового сигнала зависит также от способа его
издавания. Территориальные птицы поют свои песни, выбирая для
этого самую высокую точку местности («песенный пост»), что повы-
шает эффективность их распространения. Птицы открытых ландшаф-
тов, например жаворонки и луговые коньки, поют, летая высоко
над своей территорией.
В воде звуки распространяются с меньшим затуханием, чем в
воздухе, и потому водные животные широко используют их для
коммуникации. Развитие подводной микрофонной техники позво-
лило открыть удивительный мир звуков рыб.и китов. Пейн и Мак-
Вэй [379] в своем замечательном исследовании горбатых китов ут-
верждают, что их «песни» могут восприниматься другими китами,
находящимися за несколько сотен миль! Это несомненно рекорд
дальности в коммуникации животных.
Химические сигналы особенно хорошо развиты у насекомых и
млекопитающих (обзоры по насекомым сделали Шнейдер [423] и.
Уилсон [505], а по млекопитающим — Ролле [388]). Одна из отри-
цательных сторон химической коммуникации — трудность быстро-
го изменения сигнала. Воспроизвести стандартный запах почти не-
возможно, тогда как это легко достижимо в отношении звуковых
и зрительных сигналов. В соответствии с этим большинство хими-
ческих сигналов используется для передачи одиночных, относитель-
но стабильных сообщений. Многие млекопитающие маркируют тер-
риторию при помощи запаховых меток, часто концентрируя их на
особых местах. Запах используется также для сообщений о состоя-
нии самок млекопитающих в период размножения. Во время эстру-
са они выделяют специальные химические вещества, сигнализируя,
что они готовы к встрече с самцом. Этот тип сигнала аналогичен за-
пахам, издаваемым неоплодотворенными самками бабочек (с. 92).
Такие вещества, выделяемые во внешнюю среду и стимулирующие
размножение, называются феромонами.
Уилсон [505] и другие рассматривают в своих работах, каким
образом химическая структура запахов специфически приспособ-
лена к их функции. Половые феромоны и территориальные метки
должны быть устойчивыми, и потому составляющие их вещества
должны иметь довольно высокую молекулярную массу. В то же
время эта молекулярная масса должна быть не слишком высокой,
иначе такое вещество было бы трудно секретировать и хорошо рас-
пространять. Феромоны бабочек представляют собой поразительный
компромисс по своей устойчивости и способности к хорошему рас-
пространению. При благоприятном ветре самцы могут улавливать
запах феромонов за 4—5 км с подветренной стороны. Вместе с тем
химические вещества, используемые некоторыми муравьями в ка-
честве сигнала тревоги, не должны быть устойчивы — если бы они
были устойчивыми, то точная локализация источника непосредст-
венной опасности была бы невозможной. Эти вещества летучи и
хорошо распространяются в радиусе 3—5 см, но обычно в пределах
минуты или менее они распадаются до уровня концентрации, лежа-
щего ниже уровня обнаружения.
Зрительные сигналы могут действовать лишь на относительно
коротком расстоянии, по крайней мере в том случае, когда они
должны передать много подробной информации. В качестве простых
сигналов тревоги часто используются пятна белого цвета — хвост
оленя, хвосты кроликов,— которые могут действовать на больших
расстояниях. Связь при помощи зрительных сигналов особенно
характерна для позвоночных и головоногих моллюсков, т. е. для
животных с хорошо развитыми глазами. Интересно отметить, что
цветовое зрение практически универсально для всех групп, за ис-
ключением большинства млекопитающих. В ранней истории млеко-
питающих в течение многих миллионов лет развития, когда их
предки были ночными насекомоядными, цветовое зрение давало им
мало преимуществ и было в заметной мере утеряно. Яркая разно-
цветная раскраска некоторых рыб, рептилий и птиц поразительно
контрастирует с универсальной серой, черной и коричневой ок-
раской большинства млекопитающих. Исключениями являются
некоторые белки и приматы, как раз те две группы млекопитаю-
щих, у которых восстановилось цветовое зрение. У них мы видим
возврат к синей, красной, зеленой и желтой окраске.
Членистоногие имеют замечательное цветовое зрение, но, воз-
можно, потому, что они малы и их сложные глаза не позволяют
хорошо различать форму, зрительная сигнализация у них не очень
распространена, хотя цветовые сигналы используются в демон-
страциях ухаживания, например у бабочек и манящих крабов.
У светлячков развилась замечательная форма зрительной комму-
никации, которая может действовать на очень больших расстояниях,

108 Глава 3
Стимулы и коммуникация
109
Рис. 3.12. Самец птицы-фрегата принимает позу саморекламирования [364].
Самка сидит слева от гнезда. Детали см. в тексте.
поскольку они сами излучают свет и применяют сигнализацию
только ночью.
В естественных условиях не очень реально разделять сигналы
различных модальностей, как мы это только что делали, У живот-
ных развились эффективные комбинации сигналов, включающие,
например, и звук, и зрительный стимул. Приводимая ниже цитата
из работы Нельсона [364] (см, также рис. 3.12) создает живое
представление о том, как самцы птиц-фрегатов привлекают внима-
ние самок и приглашают их сесть на выбранное ими место для гнезда.
«Половое саморекламирование» у самцов малого фрегата пред-
ставляет собой демонстрацию раздутого горла и содержит следующие
три элемента: 1) самец показывает поднятый вверх раздутый мали-
новый зоб, голова его при этом отклонена назад и он поворачивает
ее из стороны в сторону; 2) трепеща расправленными крыльями, он
обращает вверх их нижнюю серебристую часть и 3) издает фальцетом
звуки, описанные как журчание или ржание, быстрые ху-ху-ху.
Трепетание крыльев этих птиц поразительно: самцы ориентируют
тело так, чтобы направлять свои демонстрации прямо самке, а их
крылья похожи на две огромные молящие черные руки, между ко-
торыми лежит малиновый мешок. Если самка спускается, то дви-
жения головы самца становятся более сильными и резкими. Эту
демонстрацию сопровождают специфический звук и щелканье клю-
вом, иногда прерываемые особой дрожащей трелью. Любой из трех
таких бросающихся в глаза компонентов был бы достаточно эффек-
тен. Когда же они осуществляются группой птиц все вместе, они
производят поразительное впечатление».
2. Что передается? Мы уже упоминали о некоторых типах
информации, передаваемой животными; крики тревоги и половые
феромоны — это примеры сигналов, которые передают очень про-
стые сообщения. В своем обсуждении коммуникации у позвоночных
Смит [441] предлагает учитывать различие между информацией,
которую животное-отправитель вкладывает в сигнал, и той, кото-
рую животное-получатель из него извлекает. Он называет их сооб-
щением и смыслом соответственно, и важно помнить, что они необя-
зательно совпадают. Очень часто сообщение возникает благодаря
изменению внутреннего состояния животного-отправителя. Самец
зяблика просыпается на своей территории и начинает петь. Его сооб-
щение, вероятно, не что иное, как мера его готовности к размноже-
нию. Однако птица-получатель может извлечь из него больше ин-
формации, потому что есть и другие признаки того, что она должна
удалиться. Отправитель — самец зяблика находится на своем гнездо-
вом участке, а получатель узнает, что он попал на чужую террито-
рию и реагирует соответствующим образом. В зависимости от вре-
мени года можно установить, имеет самец самку или нет (самцы,
самки которых насиживают яйца, поют гораздо меньше). Наконец,
отдельные особенности песни позволяют хозяину соседней терри-
тории отличить певца А от Б и В. Такую интерпретацию песня сам-
ца имеет, конечно, с точки зрения человека. Смысл, который она
имеет для других зябликов, вероятно, не столь детально разработан,
но он соответствует информации, которая потенциально доступна
и состоит из нескольких простых сообщений.
В гл. 5, где речь идет о конфликте, мы обсудим другие примеры
сигналов, передающих информацию о внутреннем состоянии живот-
ного, особенно в связи с вероятностью нападения или бегства.
Сигналы могут передавать целый ряд сообщений. Многие из них
чисто сексуальные, как вышеупомянутые феромоны или мерцание
светлячков. Некоторые сигналы обеспечивают обмен информацией
между родителями и потомством, другие представляют собой крики
тревоги или передают сообщения о пище. Общественные животные
часто имеют сигналы, которые помогают им поддерживать контакт,
и, возможно, не заключают в себе никаких других особых сообще-
ний. Большинство птиц, которые образуют стаи, имеет стайные сиг-
налы этого типа, Известным примером могут служить крики, из-
даваемые гусями и вьюрками в полете или перед тем как взлететь.
Такие крики часто свойственны и одиночным птицам. Вероятно,
они каким-то образом причинно связаны с полетом. В «Кольце царя
Соломона» Лоренц описывает, как стая гусей на земле «принимает
решение» о взлете. Сначала одна птица, а затем еще несколько
издают стайный крик. Если все его подхватывают, стая обычно
взлетает, но если ответило слишком мало птиц, то начавшая кри-
чать птица замолкает и стая остается на земле.
Есть несколько интересных примеров совершенно иного типа
сигналов, чье назначение — подготовить животное к действию
последующих стимулов. Явление, связанное с подобными сигнала-
ми, было названо «метакоммуникацией»; лучше всего оно известно
из игр хищных и обезьян. На рис. 3.13 показан взрослый лев, при-

1 02
Глава 3
Стимулы и коммуникация
Рис. 3.13. Метакоммуникация у львов [521]. Поза самца с прижатыми к земле
передними лапами обычно предшествует игре. Он приглашает львенка играть и
осторожно похлопывает его, если львенок к нему присоединяется.
глашающий львенка поиграть. Его поза с прижатыми к земле перед-
ними лапами не встречается ни в каких других ситуациях. Она
говорит о том, что все агрессивные действия, которые затем по-
следуют,— это игра. Собаки используют почти в точности ту же
позу [45] и могут еще вилять хвостом во время драки-игры (как
это отметил Дарвин в своей книге «Выражение эмоций у чело-
века и животных»). У обезьян в таких ситуациях появляется особая
«игровая» мимика. Игровая агрессивность составляет важную часть
развития поведения хищных, и, очевидно, была насущная необхо-
димость в условиях, которые позволяли бы животным научиться
выслеживать и нападать без повреждений и ран, неизбежных для
настоящих боев.
Градуированные и постоянные сигналы. На рис. 3.14 показаны
различные формы зрительной демонстрации, характерной для сойки
Стеллера во время территориальных столкновений. Эти формы четко
градуированы: Браун [72] обнаружил, что чем выше поднят хохо-
лок, тем более устойчива птица при агрессивных взаимодействиях.
При бегстве, а также во время ухаживания хохолок прижат к голо-
ве. Очевидно, развитие градуированных сигналов обеспечивает
большие преимущества, так как они могут передавать сообщения,
сходным образом градуированные по интенсивности. Различные
крики, испускаемые обезьяной-резусом при агрессивных стычках,
Рис. 3.14. Угол подъема хохолка у сойки Стеллера [72]. Браун использовал этот
градуированный сигнал как показатель агрессивных намерений.
можно расположить по градациям, которые вполне успешно корре-
лируют с вероятностью того, что она нападет [403]. На рис. 5.8 пока-
зана аналогичная попытка градуировать «мимику» кошек. Ком-
муникация с помощью фуражировочного танца у пчел, которую мы
подробно рассмотрим ниже, еще один пример градуированной сис-
темы сигнализации, в которой сообщение очень точно зависит от
формы сигнала.
Однако есть и некоторые отрицательные стороны сигналов, чья
форма меняется в зависимости от обстоятельств. Они могут быть не-
ясны адресату, особенно если видны лишь короткое время. Многие
сигналы постоянны по форме или варьируют только в узких преде-
лах (см. обсуждение «типичной интенсивности», гл. 6). К этому типу
сигналов относятся многие крики тревоги и ряд сигналов, свг. ;анных
с размножением, таких, как феромоны и песни самцов сверчков и
цикад, функция которых состоит в обеспечении встречи половых
партнеров. В подобных случаях имеет смысл пользоваться стерео-
типными сигналами — они производятся, только когда издающая
сигнал особь находится в полностью соответствующем состоянии.
В противоположность этому многие демонстрации ухаживания у
рыб и птиц градуированы, поскольку с их помощью необходимо пере-
дать потенциальному половому партнеру относительно подробную
информацию о готовности к спариванию, агрессивности и страхе
(обсуждение см. в гл. 5).
Различие между градуированными и постоянными сигналами
неизбежно становится менее четким, когда мы рассматриваем про-
цесс коммуникации более подробно. В начале этой главы мы обсуж-
дали способ, при помощи которого стимулы действуют как насосы
или как триггеры. Коммуникация, очевидно, может действовать
по принципу насоса, когда влияния сигналов постепенно накапли-
ваются и изменяют вероятность того, что получатель будет отве-
!
1 02
Глава 3
чать. Постоянный половой сигнал, подобный поклону самцов голу-
бей при ухаживании, может в то же время оказывать градуирован-
ное влияние на самку, потому что его нужно повторить много раз
на протяжении ряда дней, чтобы он полностью оказал свое действие.
Шлейдт [4211 называет это «тонической коммуникацией» и рассмат-
ривает другие ее примеры. Многое зависит от функции, с которой
связаны сигналы. Постоянные сигналы других типов, например
крики тревоги, должны действовать как триггеры и резко изменять
поведение особи — только в этом случае они полезны.
У коммуникационных систем животных отсутствуют неогра-
ниченные ресурсы символического языка человека. Они должны
в большой степени полагаться на обстановку и на комбинации сиг-
налов, чтобы расширить круг своих сообщений. Даже среди наших
ближайших родственников — приматов — число передач довольно
ограниченно, хотя, как мы видели (рис. 3.11 и с. 104), большие спо-
собности обезьян к обучению и долгий опыт общения в разных груп-
пах приводят к тому, что их взаимодействия в сообществе часто
очень хорошо развиты. В гл. 7 мы рассматриваем некоторые опыты
по обучению шимпанзе символическому языку. Замечателен здесь
тот факт, что, хотя они могут усваивать сложные жесты языка зна-
ков, дикие шимпанзе жестикулируют очень мало. Выпрашивание
с протянутой рукой вполне обычно для них, но за многие часы на-
блюдений Мензель [341] никогда не видел, чтобы шимпанзе подзы-
вали друг друга кивком. Этот жест оказывается одним из первых
знаков, которые выучивают молодые шимпанзе во время специаль-
ной тренировки 1164].
Одно из различий между коммуникацией у человека и живот-
ных, которому часто уделяют много внимания, — это возможность
обмана. Люди могут обманывать, но у животных коммуникация в
столь значительной степени оказывается результатом их собствен-
ного состояния, что обман кажется невозможным. Мы не можем
представить себе птицу, издающую, например, ложный сигнал тре-
воги. Похожие на глаза пятна на крыльях бабочек никак нельзя •
отнести к обману. Они вовсе не отражают ее внутреннего состояния,
это просто зрительные ключевые стимулы, эволюционировавшие
так, чтобы вводить хищников в заблуждение. Все многочисленные
насекомые (и животные других групп), которые паразитируют
в муравейниках (мнрмекофилы), подают своим хозяевам фальшивые
(и зачастую сверхнормальные) сигналы, благодаря чему их прини-
мают в гнездо [2291. «Трюк с подбитым крылом», при помощи кото-
рого некоторые из гнездящихся на земле птиц (например, ржанки)
уводят хищников от своего гнезда, объяснить гораздо труднее.
Мы слишком мало знаем о внутреннем состоянии птицы во время
демонстраций, однако не похоже, чтобы этот обман был в какой-то
степени сознательным. В этом случае мы снова видим уловку, воз-
никающую в результате эволюции,
Стимулы и коммуникация
112
Самый замечательный пример обмана, созданного эволюцион-
ным путем, дают нам светлячки. Неоплодотворенные самки Photuris
versicolor испускают видоспецифические комплексы вспышек света
в ответ на сигналы самцов, которые приближаются к ним для спа-
ривания. После спаривания самка перестает светиться, и в следую-
щие две ночи ее поведение меняется. Она принимает позу хищника
с поднятыми передними ногами и открытыми челюстями. Теперь она
снова начинает светиться, но уже не применяет кода, характерного
для ее вида. Она испускает сигналы, характерные для родственного
ей более мелкого вида из рода Photinus. Когда самец Photinus
приближается к ней, она убивает и съедает его! Ллойд [301, 302],
который открыл этот экстраординарный случай обмана, справед-
ливо назвал самок Photuris «роковыми женщинами».
3. Как мы можем изучать, происходит ли коммуникация?
До сих пор, рассматривая процесс коммуникации, мы считали, что
социальные сигналы эффективны в вызывании соответствующих
реакций. Исходя из эволюционных представлений, мы уверены в
том, что у животных существуют системы коммуникации, но мы
никогда не узнаем достаточно подробно, как они действуют и каковы
их пределы, если не перейдем к измерениям.
Многие взаимодействия между особями одного вида осуществля-
ются при помощи разнообразных потенциальных сигналов. Опыты
с использованием моделей или с заменой обычного диапазона сти-
мулов помогают определить, каковы ключевые свойства системы
коммуникации, и это составляет первую стадию настоящего анали-
за. Мы уже описали и проиллюстрировали ряд модельных экспери-
ментов (рис. 3.3 и 3.9), которые доказывают, ч .о красное брюшко
самца колюшки и пятно на клюве серебристой чайки в определенных
ситуациях служат основными компонентами процесса коммуника-
ции. Совершенно аналогичным образом ранние опыты, в которых
было обнаружено, что самки сверчков заходят в рупор громкогово-
рителя, передающего запись песни самца, впервые доказали дей-
ственность звуковой коммуникации у насекомых. Многие опыты
с половыми феромонами у бабочек также дают однозначные резуль-
таты — никакие другие стимулы самок не могут действовать
на таких больших расстояниях.
Доказано, что некоторые виды коммуникации включают пере-
дачу довольно тонких сигналов. У мономорфных видов птиц —
таких, где самцы и самки похожи друг на друга,— определение пола
составляет проблему. Единственное заметное различие между сам-
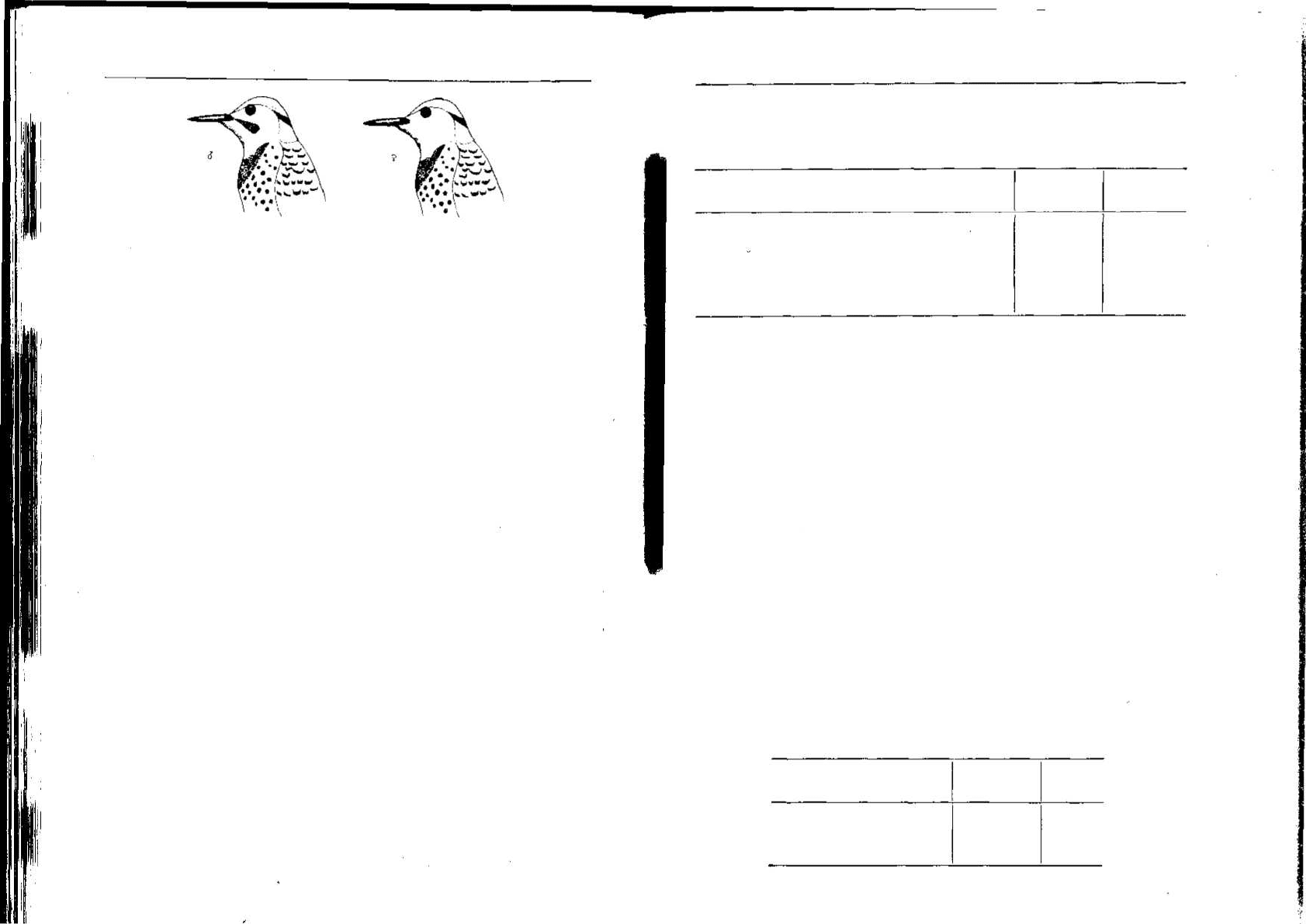
цом и самкой американских, золотистых дятлов состоит в маленьких
черных усиках самца (рис. 3.15). Нобль 1367] поймал самку дятла,
вклеил ей усики из черных перьев, а затем выпустил на ее гнездо-
вом участке. Самец сразу напал на нее и выгнал. Когда усы были
сняты, самец сейчас же снова принял ее обратно. Тот факт, что ре-
акцию самца можно вызвать или прекратить этим способом, ясно
102
Глава 3
Стимулы коммуникация
115
Таблица 3.1
Рис. 3.15. Головы самца (слева) и самки (справа) золотистого дятла. Черные уси-
ки самца — ключевой признак для определения пола.
свидетельствует о том, что сигнальное значение усов перевешивает
значение всех других признаков. Смит [440! исследовал замеча-
тельный пример межвидового общения такого рода у четырех видов
гнездящихся в Арктике чаек. Все они в основном белые птицы с се-
рыми спинами. Человеку трудно различать их — пожалуй, это
можно сделать лишь по рисунку на концах крыльев, однако при
ближайшем рассмотрении оказалось, что эти виды различаются
также по цвету глаз. Они варьируют по цвету радужки, по цвету
узкого кольца пигментированной кожи вокруг глаз, что более за-
метно, и по цвету внутренней поверхности век, видимой, когда глаза
открыты. Смит ловил чаек и перекрашивал кольцо кожи вокруг глаз
в цвета, характерные для других видов. Если это делалось в начале
сезона размножения, то оказывалось достаточным для разрушения
того практически полного барьера, который в нормальных условиях
препятствует межвидовому спариванию. При этом часто удавалось
разрушить естественно сложившиеся пары, хотя они обычно сохра-
нялись, если перекрашивание происходило позднее. Таким образом,
у чаек опознавание сородичей зависит главным образом от окрас-
ки кольца кожи вокруг глаз.
Идентификация стимулов, используемых при общении, состав-
ляет первый шаг анализа, далее важно попытаться измерить их
эффективность. Это не всегда оказывается просто. Во-первых,
часто невозможно решить, означает ли отсутствие ответа-, что полу-
чатель неспособен воспринять сообщение отправителя или же что
он получил его, но просто не может ответить по какой-либо причине.
Эта трудность существует даже в отношении социальных сигналов,
которые — если они действуют,— обычно вызывают немедленный
ответ; гораздо больше трудностей возникает в отношении сигналов,
которые действуют не как триггеры, а тонически, скорее как насосы.
При исследовании
Л
того вопроса ничто не заменит тщательного
многократного наблюдения столкновений животных в группе, а
также тестирования эффективности сигнала путем регистрации отве-
та «адресата» в ситуациях, когда определенный сигнал подается
или когда он отсутствует. В гл. 5 мы опишем некоторые примеры
подобного анализа враждебных столкновений между птицами;
Поведение доминантных мангобеев по отношению к подчиненным
г/осле того, как последние принимают (а) или не принимают (б)
позу подставления
Поведение
Доминант не
нападает
Доминант
нападает
(а) Доминант приближается к подчиненному, ко-
53 0
торый принимает позу подставления
(б)
Доминант приближается к подчиненному, кото-
21 9
рый не принимает позы подставления, не делает
других жестов и не издает звуков
здесь же мы используем в качестве примера работу Чалмерса [88]
с мангобеями.
Мангобеи — это крупные лесные обезьяны, обитающие в Аф-
рике. Чалмерс описал у них ряд звуковых и зрительных демон-
страций, а затем проверил их эффективность для процесса комму-
никации описанным выше способом. Ему пришлось объединить
данные по большому числу столкновений между различными осо-
бями, чтобы набрать достаточное для сравнения количество случаев,
где демонстрации применялись бы или же отсутствовали. Одна из
обычных зрительных демонстраций — это «подставление», когда
подчиненная обезьяна любого пола поворачивается спиной к живот-
ному более высокого ранга и «приглашает» его сделать садку (см.
рис. 8.10 и обсуждение на с. 203 и 321). Подставление часто истолко-
вывается как умиротворяющая демонстрация, которая останавлива-
ет нападение. В табл. 3.1 приведены данные, касающиеся послед-
ствий приближения доминантных животных к подчиненным, когда
последние принимают (а) или не принимают (б) позу подставления.
Поза подставления снижает долю нападений (статистически
достоверно при рЛ0,01), т. е. сигнал подчиненного животного дей-
ствует. Чалмерс изучил также влияние одного из криков манго-
беев — низкого повторяющегося ворчания,— часто издаваемого
при сближении двух особей. На основании данных табл. 3.2 воз-
Таблица 3.2
Количество враждебных и мирных встреч при
сближении с молчащим или ворчащим мангобеем
Поведение
Враждебное
Мирное
Приближается молча
19
25
Приближается с ворчанием
3
30
и

88
Глава 3
никло предположение, что этот крик снижает вероятность враждеб-
ных взаимодействий.
Полученные результаты достаточно показательны, но, как под-
черкивает Чалмерс, сами по себе они не являются веским доказа-
тельством коммуникации при помощи позы подставления или ворча-
ния. Возможно, что обезьяны могут по каким-то признакам опреде-
лять, собирается ли доминант нападать или нет, и принимают позу
подставления только в последнем случае. Кроме того, обезьяны, ко-
торые не собираются нападать, могут ворчать при приближении
друг к другу, тогда как враждебно настроенные обезьяны прибли-
жаются молча. Все это, конечно, не очень вероятные объяснения,
но мы можем исключить их только при дальнейшем изучении всех
случаев, где эти демонстрации встречаются.
ТАНЕЦ ПЧЕЛ
Этот раздел лучше всего заключить описанием танца пчел. Это
не только одна из самых замечательных систем коммуникации у жи-
вотных, до недавнего времени вопрос о танцах пчел служил источ-
ником непрерывной дискуссии, порожденной теми же проблемами
измерения и интерпретации, которые мы обсуждали выше.
Тот факт, что пчелы должны передавать информацию о местах
массового цветения, был известен на протяжении веков, но отныне
он всегда будет связан с именем фон Фриша. Он первым объяснил
природу этого вида коммуникации и в своей книге (1561 дал его
полное описание. Подобно многим исследователям до него, фон
Фриш отметил, что если выставить блюдечко с сахарным сиропом для
привлечения пчел, то пройдут многие часы, прежде чем первая пче-
ла найдет его, сядет и напьется. Однако после того, как одна пчела
обнаружила источник пищи, прибытие массы других сборщиц
(фуражиров) становится делом нескольких минут — информация
о новом источнике пищи каким-то образом распространяется. Фон ,
Фришу потребовалось около 20 лет кропотливых наблюдений и
опытов, прежде чем он разработал удовлетворившее его представле-
ние о системе коммуникации у пчел. Заключения, к которым он
пришел, были столь необычными и беспримерными, что, как сам
он говорил, ни один хороший ученый не должен принимать их без
доказательств После второй мировой войны другие зоологи подтвер-
дили полученные фон Фришем результаты, а некоторые принимали
участие в его заключительных опытах. Сейчас сомнений относитель-
но природы самого танца пчел уже не осталось.
Фон Фриш метил разведчиц, когда они пили из блюдечка сахар-
ный сироп, а затем наблюдал за их поведением при возвращении
в улей, стенки которого были для этого сделаны стеклянными. Раз-
ведчица обычно контактирует с массой других пчел на вертикальной
поверхности сотов и отдает им сахарный сироп из своего перепол-
Стимулы и коммуникация 119
117
Рис. 3.16 Круговой танец рабочей пчелы на вертикальной поверхности сота: ее
путь обозначен штриховой линией. Обратите внимание, как близко следуют за ней
другие пчелы.
ненного зобика. Затем она начинает танцевать, и сначала мы рас-
смотрим случай, когда блюдечко с сиропом, которое она только что
посетила, находится близко к улью, в радиусе 50 м. Ее быстрый та-
нец происходит по кругам, диаметр которых приблизительно равен
длине ее тела. Пчела описывает круги по очереди то влево, то впра-
во. Она остается примерно на одном месте сота и может танцевать
до 30 с, прежде чем двинуться дальше. Другие пчелы окружают тан-
цовщицу, часто своими антеннами они касаются ее тела и точно сле-
дуют всем движениям, повторяя ее кругообразный путь (рис. 3.16).
«Круговой танец», как его называют, стимулирует других рабочих
пчел покинуть улей и искать взяток поблизости. Оказывается, он
передает информацию о том, что нужно «искать в пределах 50 м».
С помощью кругового танца пчела может передавать также некото-
рые обонятельные сигналы, потому что если источник пищи имеет
запах, то пчела принесет его следы на своем теле, а возможно, и
на самом сахарном сиропе. Если же блюдечко с сахаром не имеет
запаха, то сборщица может, пока пьет сироп, «пометить» его выде-
лениями брюшных назановых пахучих желез.
До сих пор в танце пчел нет ничего особенно выдающегося, по-
тому что многие муравьи и термиты имеют похожие «мобилизую-
щие» демонстрации, а также обладают феромонами, которые помога-
ют им организовать сбор пищи при появлении нового ее источника
рядом с гнездом [505]. Возможности коммуникационной системы
пчел не раскрываются до тех пор, пока источник пищи не помещают
дальше от улья, на расстоянии 100 м.
Фон Фриш наблюдал, что, когда блюдечки с пищей отодвигали
более чем на 50 м, круговой танец сборщицы постепенно менялся
по форме. Между поворотами появлялся короткий прямолинейный
пробег, и в это время танцовщица быстро виляла брюшком из сторо-
ны в сторону. При расстоянии до кормушки около 100 м движения
пчелы приобретали типичную форму «виляющего танца», показан-
ного на рис. 3.17, и эта форма сохранялась, если блюдечко отодви-