Pugnaire F.I. Valladares F. Functional Plant Ecology
Подождите немного. Документ загружается.

RESPONSES TO SPATIAL HETEROGENEITY
Photosynthesis and Growth
At the most basic level, spatial heterogeneity in light creates variation in the resources
available and hence in photosynthesis and growth. Because light is highly variable but is
often the most limiting resource in forest understories, correlations between light availability,
photosynthetic parameters, and seedling growth have been demonstrated in many species
(Montgomery and Chazdon 2002, Delagrange et al. 2004, Pearcy et al. 2004). Growth of
understory saplings of two shade-tolerant Hawaiian tree species was highly correlated with
spatial variation in the mean annual potential minutes of sunflecks received as estimated from
fisheye photographs (Pearcy 1983). Similar results have been obtained with understory
saplings in Costa Rica (Oberbauer et al. 1988) and oak seedlings in Japan (Washitani and
Tang 1991). However, spatial heterogeneity in growth is not always correlated with estimates
of sunfleck availability, and in some cases diffuse light availability has been a better predictor
of growth (Tang et al. 1992). Pfitsch and Pearcy (1992) found no correlation between
estimates of sunfleck light availability and growth of the redwood forest understory herb,
Adenocaulon bicolor. Variation in daily carbon gain and in the proportion of the annual
carbon gain contributed by photosynthetic utilization of sunflecks was, however, highly
correlated with the variation in sunfleck availability (Pearcy and Pfitsch 1991). Moreover,
removal of sunflecks with shadow bands designed to block sunfleck transmission along the
solar track but pass most of the diffuse PFD, significantly reduced growth and reproduction
(Pfitsch and Pearcy 1992). Thus, there is no doubt that sunfleck utilization was important to
these plants but other limitations may have prevented translation of the spatial variation in
sunfleck PFD into spatial variation in growth. There is evidence for only minor losses of
carbon gain due to photoinhibition in sunflecks (Pearcy 1994) so other factors such as
drought stress and increased competition in the brighter microsites need to be considered.
Plasticity of the Photosynthetic Apparatus in Response to Spatial Heterogeneity
One of the most universal responses to increased light availability is a plastic response of leaf
photosynthetic properties that results in development of sun leaves that have higher photo-
synthetic capacities per unit area (Amax
area
), greater leaf thickness, and a greater leaf mass
per unit leaf area (LMA). The plasticity expressed by different species in different growth
situations varies widely, ranging from 1.25- to a 3-fold or in a few cases a 5-fold difference in
Amax
area
between sun and shade environments (Bjo
¨
rkman 1981, Pearcy and Sims 1994,
Chazdon et al. 1996, Strauss-Debenedetti and Bazzaz 1996, Huante and Rincon 1998). The
plasticity of Amax
area
can be examined either in terms of the relative change in photosynthetic
rates achieved under light-saturating conditions, or in terms of the range of light environ-
ments over which this plasticity is expressed. Most studies have focused on the former by
comparing plants grown in low- and high-light environments designed to mimic shade and at
least partial sun environments. These are useful for comparing the response to environments,
either simulated or natural, such as a gap and understory but because of the nonlinear
response of most phenotypic traits and the potential for combining acclamatory and inhibi-
tory responses in a stressful environment, they are usually not satisfactory in identifying
species differences in plasticity related to their regeneration niche. When compared in this
way, fast-growing early successional species favoring high-light environments seem to exhibit
greater plasticity of Amax
area
, as compared with more slowly growing, shade-tolerant later
successional shrub and tree species (Strauss-Debenedetti and Bazzaz 1996, Yamashita et al.
2000). Some care is required in interpreting plasticity since the response to the high-light
environment may be a mixture of acclimation and photoinhibition with the latter acting to
reduce Amax
area
(Mulkey and Pearcy 1992). In addition, extremely shaded light environments
Francisco Pugnaire/Functional Plant Ecology 7488_C007 Final Proof page 220 30.4.2007 7:57pm Compositor Name: DeShanthi
220 Functional Plant Ecology
may interfere with the leaf development of shade-intolerant species, reducing their Amax
area
.
The way to avoid this problem is to examine photosynthetic responses over multiple light
environments but this has been done in only a few studies (Sims and Pearcy 1989, Valladares
et al. 2000, Muraoka et al. 2003). An interesting approach is to study transplanted seedlings along
light gradient such as across a forest edge in a gap forest to pasture transition, giving a wide range
of light environments (Chazdon 1992, Montgomery 2004). In general, results obtained from
comparisons across multiple environments reveal a strong dependence of Amax
area
on the
daily light availability up to moderate daily PFDs but little further increase. For example,
Montgomery (2004) found that most growth and photosynthetic traits exhibited nonlinear
responses to light availability with the greatest changes between 0.5% and 20% canopy
transmittance of diffuse light with little additional change at higher canopy transmittances.
Given the interacting effects of leaf development, acclimation, and photoinhibition, the
range of light environments over which plasticity is expressed can differ between species, and
may be as important ecologically as the actual change in Amax
area
itself. Sims and Pearcy
(1989) compared the shade-tolerant, rain-forest herb, Alocasia macrorrhiza to the crop
species, Colocasia esculenta in five different light environments ranging from deep shade to
50% of full sun. Both exhibited a 2.2-fold difference in Amax
area
but the range of light
environments over which this was expressed was higher in Colocasia than in Alocasia.
Comparisons of 16 gap-dependent and shade-tolerant Psychotria species across three light
environments from deep shade to 25% of full sun reveals the diversity of responses that can
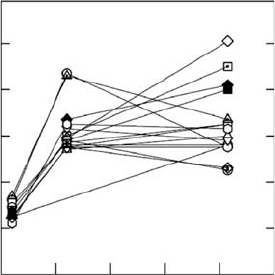
occur (Figure 7.2). Overall, the plasticity of Amax
area
differed little among the species but the
responses to the changes in light environments contrasted markedly. Most species exhibited
10
MAR
BRA
GRC
PUB
EME
HOR
MIC
IPE
CHA
ACU
GRD
TEN
PSY
RAC
PIT
LIM
8
6
4
2
0
024
Growth light environment
(mol photons m
−2
day
−1
)
Photosynthetic capacity
(µmol m
−2
s
−1
)
6810
FIGURE 7.2 Response of photosynthetic capacity (Amax) of 16 Psychotria species to growth in three
different light environments simulating understory shade to gap conditions. The abbreviations on the
right side are for the different species and are ordered identically to the symbols shown for the highest
light environment on the right side. The abbreviations are: MAR, Psychotria marginata; BRA, Psycho-
tria brachiata; GRC, Psychotria graciflora; PUB, Psychotria pubescens; EME, Psychotria emetica; HOR,
Psychotria horizontalis; MIC, Psychotria micrantha; IPE, Psychotria ipecacuanha; LIM, Psychotria
limonensis; CHA, Psychotria chagrensis; ACU, Psychotria acuminata; GRD, Psychotria granidensis,
TEN, Psychotria tenuifolia, PSY, Psychotria psychotrifolia; RAC, Psychotria racemosa; PIT. Psychotria
pittieri. The abbreviations in boldface are for gap-dependent species and show that there is no relation-
ship between gap versus understory species and plasticity of photosynthetic capacity. Each data point is
the mean of at least three determinations on different plants. (Adapted from Valladares, F., Wright, S.J.,
Lasso, E., Kitajima, K., and Pearcy, R.W., Ecology, 81, 1925, 2000. With permission.)
Francisco Pugnaire/Functional Plant Ecology 7488_C007 Final Proof page 221 30.4.2007 7:57pm Compositor Name: DeShanthi
Responses of Plants to Heterogeneous Light Environments 221
the largest increase in Amax
area
between the low and the intermediate light environments.
Some species exhibited a moderate increase between the intermediate and the high-light
environments, others no increase and still others exhibited a decline in Amax
area
from the
intermediate to the high-light environment. These responses of Amax
area
were not correlated
with habitat requirement (gap-dependent vs. understory) of the species, though an overall
measure of plasticity integrating both physiological and morphological characters showed
that gap-dependent species had greater plasticity. Of course Amax
area
is only one of a suite of
characters, and changes in leaf angle and so on may function to reduce the actual range of
incident PFD experienced by the plant to avoid conditions that may cause photoinhibition.
Indeed the PFD on the leaf surfaces is often close to the PFD that is just saturating for
photosynthesis even though the incident PFD on a horizontal surface is much higher. It
would be unrealistic to expect that increasing the PFD would result in a further increase in
Amax
area
. Indeed, the more common result may be photoinhibition.
Only a few studies have examined the plasticity of the photosynthetic apparatus to the
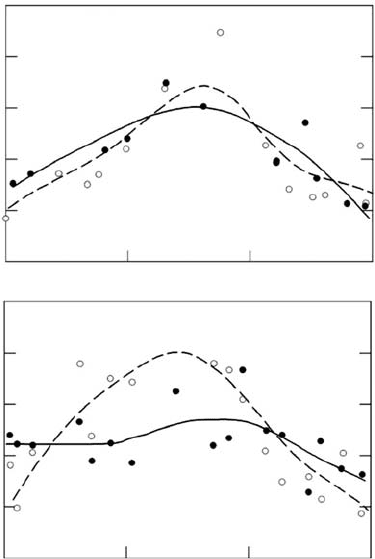
spatial heterogeneity of light environments in the field. Chazdon (1992) found that Amax
area
of an early successionial shrub, Piper sancti-felicis, was closely related to light availability
along transects across natural gaps. A later successional congener, Piper arieianum, exhibited
a weaker association of Amax
area
with light availability, possibly because of a greater
susceptibility to photoinhibition in the gap center (Figure 7.3). Comparisons of a habitat
10
0
10
P. sancti-felicis
P. arieianum
8
6
4
2
0
51015
Percent of full sunlight
8
6
4
2
0
10
0
10
8
6
4
2
0
5
Distance from west
g
ap ed
g
e (m)
Amax (µmol CO
2
m
−2
s
−1
)
10 15
8
6
4
2
0
FIGURE 7.3 Gradients of light (open circles) and Amax (closed circles) of seedling of the gap species,
Piper sancti-felicis (top), and the understory species, P. arieanum (bottom) across a small gap. Photo-
synthetic capacity tracks the light environment much better in the gap-dependent species than in the
understory species. (Adapted from Chazdon, R.L., Oecologia, 92, 586, 1992. With Permission.)
Francisco Pugnaire/Functional Plant Ecology 7488_C007 Final Proof page 222 30.4.2007 7:57pm Compositor Name: DeShanthi
222 Functional Plant Ecology
specialist, the gap-dependent pioneer species, Piper auritum, with a habitat generalist species
with respect to light, Piper hispidum, revealed similar responses of Amax
area
to light avail-
ability but less sensitivity of dark respiration and LMA to light in the specialist species.
Although the net result was similar the two species may differ in the mode of acclimation.
Within the narrower range of light environments in the understory, Chazdon and Field (1987)
found no correlation between microsite light availability and Amax
area
in response to micro-
site variation in the understory but did find increasing Amax
area
with increasing light
availability in gap environments. These results suggest that below a lower threshold PFD
no further adjustments in photosynthetic capacity occur.
Sunflecks are a primary source of heterogeneity in daily PFD in the understory, but these
are certainly too unpredictable for any advantage in daily carbon gain to accrue from
adjustment of Amax
area
. Sims and Pearcy (1993) found no differences in Amax
area
among
Alocasia macrorrhiza plants grown under either short or long sunflecks or constant PFD
environments where each environment had identical daily PFDs. Pearcy and Pfitsch (1991)
found that Amax
area
, LMA, and leaf N per unit area of the A. bicolor plants grown under
shadow bands were all positively correlated with diffuse PFD levels. However, no correlation
between these measures and either total PFD of sunfleck PFD was found for the adjacent
control plants that received sunflecks. A filtering mechanism that ignored the unpredictable
component of variation in daily PFD associated with sunflecks but allowed response to the
much less variable but more predictable diffuse component would explain these results.
Maximizing Photosynthesis in High Light
Maximizing photosynthetic performance in high light requires investment in the carboxyla-
tion and electron transport capacity necessary to support high photosynthetic rates coupled
with a sufficiently conductive pathway for CO
2
diffusion through the stomata and to the site
of carboxylation in the chloroplasts. Growth in high versus low light has been shown to result
in increases in activities (concentrations) per unit area of the primary carboxylating enzyme
ribulose-1,5 bisphosphate carboxylase=oxygenase (Rubisco), Photosystem II (PSII) electron
transport capacity, the electron carrier, cytochrome f, and chloroplast coupling factor that is
involved in ATP synthesis (Bjo
¨
rkman 1981, Anderson and Osmond 1987). It is likely that
nearly all components in the photosynthetic carbon reduction cycle (PCRC) and electron
transport must increase in a coordinated manner to maintain a balance between capacities of
each component step. Indeed, acclimation seems to result in at most only a small increase in
the ratio of electron transport to carboxylation capacity (Sims and Pearcy 1989, Thompson
et al. 1992, Wullschleger 1993). Stomatal conductance must also increase to maintain a
balance between the capacity for supply and for utilization of CO
2
. Since internal diffusion
limitations across cell walls and membranes to the Rubisco in the chloroplast are significant
(Evans and Von Caemmerer 1996) these must also be adjusted to accommodate sufficient
transport as photosynthetic capacity is increased.
Insight into the causes of the plasticity in photosynthetic capacity can be gained by
comparing differences in photosynthetic capacities calculated on a per unit leaf area basis
(Amax
area
) with those calculated at per unit leaf mass basis (Amax
mass
). Across ecologically
and geographically broad species comparisons of high light-adapted species, thinner leaves
with lower LMA have higher Amax
mass
than thicker, high LMA leaves (Reich et al. 1997).
Across species and especially across life forms much of the difference in Amax
mass
is clearly
due to genetically determined differences in concentrations of photosynthetic machinery per
unit mass. From gas exchange analysis, Rubisco carboxylation capacity and electron trans-
port capacity are higher in the thin-leaved species and the stomatal and liquid phase
conductances to CO
2
also increase concurrently. The positive relationship between Amax
mass
and leaf nitrogen per unit mass is also consistent with a greater investment per unit mass in
Francisco Pugnaire/Functional Plant Ecology 7488_C007 Final Proof page 223 30.4.2007 7:57pm Compositor Name: DeShanthi
Responses of Plants to Heterogeneous Light Environments 223
photosynthetic enzymes. Liquid phase diffusion limitations may also be important. Sclero-
phylous leaves that are characterized by low Amax
area
are typically hypostomatous, have
been shown to have a lower CO
2
transfer conductance both through the intercellular air
pathway and across the cell wall and liquid phase to the site of carboxylation than mesophytic
leaves (Parkhurst and Mott 1990, Evans and Von Caemmerer 1996). Investment in thick cell
walls characteristic of sclerophylous leaves dilutes the photosynthetic machinery resulting in
low Amax
mass
. Amphistomaty serves to overcome limitations to intercellular airspace diffu-
sion in high photosynthetic capacity leaves by providing a pathway through both epidermi
(Mott et al. 1982, Parkhurst and Mott 1990).
For light acclimation, the picture that emerges is somewhat different in that leaves that
develop following a transfer of the plant from low to high light have higher LMA, are thicker,
and have a higher Amax
area
and g
s
than leaves that develop in the shade before the transfer.
However, in many, perhaps most, species there is little or no change in Amax
mass
despite the
change in leaf thickness. An increase in leaf thickness occurs because leaves developing in the
sun typically have two layers of palisade, or longer palisade cells (Kamaluddin and Grace
1992), above a spongy mesophyll, whereas those developing in the shade have only a loosely
organized and relatively undifferentiated and thin mesophyll (Chabot and Chabot 1977,
Bjo
¨
rkman 1981). This is consistent with a constant amount of photosynthetic machinery
per unit mass but with the greater LMA in sun leaves resulting in a greater concentration of
photosynthetic machinery per unit area. Similarly, significant relationships between Amax
area
per unit area and N per unit area have been commonly found in comparisons of plants grown
in the sun and the shade (Seemann et al. 1987, Evans 1989b, Hikosaka and Terashima 1996),
whereas there is no significant correlation between assimilation rates and N on a mass basis
for most species. Indeed, for many species, increased leaf thickness resulting in increased cell
and chloroplast volume per unit leaf area may be the only mechanism involved in the
observed plasticity of Amax
area
since Amax
mass
remains nearly constant across light envir-
onments (Ellsworth and Reich 1992, Sims and Pearcy 1992). Since the leaf anatomy is
largely specified early in leaf development (Sims and Pearcy 1992), mostly or fully devel-
oped leaves of many species show little or no capacity for adjusting Amax
area
(Mulkey and
Pearcy 1992, Sims and Pearcy 1992, Newell et al. 1993, Oguchi et al. 2005). Some species,
however, can increase their Amax
area
moderately even in fully developed leaves (Chazdon
and Kaufmann 1993, Kursar and Coley 1999, Yamashita et al. 2000). Species that can
respond may do so primarily by increases in chloroplast numbers or volume that fill cell
wall locations unoccupied by chloroplasts when the leaf developed in the shade. On the
other hand, species that are unable to respond may lack unoccupied locations for chloro-
plasts (Oguchi et al. 2003, 2005).
High-light environments, in addition to supporting higher photosynthetic carbon gains,
also present circumstances where excess light may be potentially damaging. Moreover, this
photoinhibition may be exacerbated by the co-occurrence of high temperatures and water
stress (Mulkey and Pearcy 1992, Koniger et al. 1998). Upper canopy leaves in tropical forests,
and also leaves in treefall gaps can reach temperatures 108C above air temperature (Koniger
et al. 1995, Niinemets and Valladares 2004). On the other hand, lower canopy leaves are
usually near air temperatures. Thus, morphological and biochemical mechanisms that min-
imize photoinhibitory damage are important in acclimation to high light. Steep leaf angles
and preferential orientation of leaves function to minimize the potential for photoinhibition
yet maintain sufficient fluxes to also maximize photosynthetic carbon gain (Ehleringer and
Forseth 1980, Ludlow and Bjo
¨
rkman 1984, Gamon and Pearcy 1989). Within plant crowns,
upper leaves often have steep leaf angles, decreasing the receipt of solar radiation at midday,
whereas lower leaves have shallower leaf angles to maximize light capture (Valladares 1999).
In addition to affording structural photoprotection in the upper canopy, changes in leaf
angles decrease vertical heterogeneity in intercepted light.
Francisco Pugnaire/Functional Plant Ecology 7488_C007 Final Proof page 224 30.4.2007 7:57pm Compositor Name: DeShanthi
224 Functional Plant Ecology
Structural photoprotection is also accompanied by biochemical photoprotection involv-
ing down-regulation of PSII via the xanthophyll cycle where excess energy is dissipated by
heat (Demmig-Adams and Adams 1992b, 1996, 2000). Sun leaves possess greater total pool
sizes of xanthophyll-cycle pigments and a greater capacity for rapid conversion of violox-
anthin to antheroxanthin and zeaxanthin that invokes this photoprotection (Thayer and
Bjo
¨
rkman 1990, Demmig-Adams and Adams 1992a, 1994). In addition, sun leaves have a
greater capacity for scavenging of active oxygen free radicals that can cause photodamage
(Grace and Logan 1996). When shade leaves are exposed to PFD in excess of the capacity of
these mechanisms, photodamage to PSII may occur, requiring operation of a PSII repair
cycle for recovery (Geiken et al. 1992, Oquist et al. 1992, Anderson et al. 1995).
Maximizing Photosynthesis in Shade
Maximizing leaf photosynthetic performance at the low PFDs inherent in shaded, diffuse-
light conditions requires (1) maximizing light absorptance, (2) maximizing the quantum yield
with which this light is used for CO
2
assimilation, and (3) minimizing respiratory losses.
Careful comparisons under conditions where photorespiration is suppressed (low O
2
or high
CO
2
partial pressures) have unequivocally established that there are no intrinsic differences in
quantum yields between C
3
species adapted to sun and shade habitats or, provided that no
stress effects interfere, among leaves on plants grown in high and low light (Bjo
¨
rkman and
Demmig 1987, Long et al. 1993, Singsaas 2001). The observed quantum yields are consistent
to those derived from stoichiometric requirements for production of the ATP and NADPH
necessary for carbon metabolism, small inefficiencies in energy transfer inherent within the
chlorophyll antennae and reaction centers, and energy use for other processes such as nitrate
reduction. Thus, there is essentially no scope for further improvements in shade as compared
to sun leaves. Photorespiratory losses under normal atmospheric concentrations reduce the
achieved quantum yields under natural conditions, but this is slightly different for sun and
shade plants. Recently, it has been reported that quantum yields are reduced in extremely low
light possibly because of insufficient buildup of the transthylakoid proton gradient required for
ATP regeneration (Timm et al. 2002, Kirschbaum et al. 2004). Since quantum yields were
greater than 85% restored by PFDs as low as 5 mmol m
2
s
1
, the loss of quantum yield would
have an effect on carbon gain only early or late in the day, or only in the shadiest microsites.
Quantum yields are frequently reduced because of photoinhibition, especially when high
light is coupled with high leaf temperatures, water stress, or low nitrogen nutrition (Bjo
¨
rkman
et al. 1981, Bjo
¨
rkman and Powles 1984, Ferrar and Osmond 1986, Mulkey and Pearcy 1992,
Koniger et al. 1998). Shade leaves are more susceptible to photoinhibition than sun leaves in
this regard and observed differences in quantum yields may often be due to the occurrence of
these other environmental factors and their interaction with high PFD. On the other hand,
higher CO
2
concentrations in the understory and the high values of intercellular to ambient
CO
2
pressures (c
i
=c
a
) characteristically observed under diffuse light conditions can enhance
quantum yields for CO
2
uptake by 10%–20% due to reduced photorespiration (Pfitsch and
Pearcy 1989a, DeLucia and Thomas 2000, Singsaas et al. 2000). This enhancement could be
important under the conditions of rising atmospheric CO
2
in allowing plants to occur in
shadier microsites than would be the case under current CO
2
concentrations.
Comparisons across many species also reveal no consistent differences in leaf absorptance
or chlorophyll (Chl) concentration per unit leaf area between sun and shade leaves (Bjo
¨
rkman
1981). Chl concentrations per unit area are typically sufficient to absorb 85%–92% of the
incident photosynthetically active radiation with the remainder lost to either reflection or
transmission. The lower LMA of shade leaves means that to maintain Chl per unit area
constant there has to be an increased investment in Chl per unit mass. Although shade leaves
in particular could benefit in terms of increased photosynthesis from increased Chl per unit
Francisco Pugnaire/Functional Plant Ecology 7488_C007 Final Proof page 225 30.4.2007 7:57pm Compositor Name: DeShanthi
Responses of Plants to Heterogeneous Light Environments 225
area and hence leaf absorptance, the strongly diminishing returns of further investment in
Chl–protein complexes required to affect a significant change in light capture may preclude it.
A doubling of the Chl concentrations is required to increase leaf absorptance by just 5%–6%
from the levels given earlier. Although there are only 4 mmol N in a mol of Chl, the associated
proteins in the Chl–protein complexes contain 21–79 mmol N mol
1
Chl (Evans 1986, 1989a).
Chlorophyll a=b ratios are much lower in shade than sun leaves and it was suggested that this
may represent a mechanism for increasing absorption in the 640–660 nm wavelength range
where PFD in the understory is somewhat greater than that at longer wavelengths where Chl
in the overstory canopy has its maximum absorption. The advantage, however, may lie more
in a significant savings of N since Chl b is part of the light-harvesting chlorophyll–protein
complex II (LHCII), which contains only 25 mmol N mol
1
Chl, whereas Chl a in PSII
represents an investment of 83 mmol N mol
1
Chl (Evans 1986). The lower Chl a=b ratio
results from increased LHCII per unit leaf area coupled with decreased concentrations of the
PSII core complex, which contains only Chl a. Since PSII complexes contain more N, the shift
to more LHCII while maintaining total Chl per unit leaf area constant results in a signifi-
cantly greater light capture per unit N invested. This savings is possible in shade, but not sun
leaves because high capacities of PSII electron transport are needed in the high light.
Acclimation of Dark Respiration
To maintain a positive carbon return, gross assimilation rates must clearly exceed respiration
rates. It is now widely documented that one of the most important differences between sun
and shade leaves are the much lower respiration rates of the latter (Grime 1966, Bjo
¨
rkman
1981). Leaf respiration itself and especially the mechanisms responsible for lower rates in
shade leaves are not well understood. Respiration rates may be affected by the capacity of the
respiratory pathways, substrate concentrations, rates of energy and reductant demand, and
the extent of alternative pathway engagement. In addition, mitochondrial respiration is
substantially inhibited by light at PFDs as low as 3 mmol m
2
s
1
(Atkin et al. 1998, 2000).
Thus, measurements done in the dark, the commonly used method for estimating respiration,
rather than the more complex procedure of Laisk (1977), must be corrected in models
exploring carbon balance in understory plants.
Mitochondrial respiration occurs in response to growth and maintenance processes. Since
the low available light severely limits carbon gain and growth in the shade, respiration rates of
the leaves of shade plants are correspondingly low, which is critical for maintaining a positive
carbon balance in understory light regimes. The observed respiration rates of sun and shade
plants reflect mostly the differing resource supplies and demands. Respiration rates appear
to be differently regulated in sun and shade species (Noguchi et al. 1997, Noguchi and
Terashima 1997). In the high-light species, Spinacia oleracea, respiration rates were limited
by the carbohydrate supply in the leaves, whereas in the shade-adapted species, Alocasia
odorata, they were limited by a low demand for ATP due to its slow growth rate. Transfer
from high to low or from low to high light typically invokes a corresponding decrease or
increase in dark respiration rates over the following 4–6 days (Sims and Pearcy 1992, Noguchi
et al. 2001b). In shade plants such as Alocasia odora, feeding experiments involving either
carbohydrates or respiratory inhibitors show that this decline in respiration on transfer
from high to low light occurs because of the reduced demand for ATP. Conversely, in high
light, more respiration may be needed for repair of PSII photodamage or for the greater
phloem loading as growth accelerates and biosynthesizes. Overall, the maintenance costs
of the photosynthetic apparatus are lower in shade than those in high light-acclimated
leaves because of the lower soluble protein levels (Noguchi et al. 2001a) and likely lower
costs for repair of photodamage. However, even in high light-acclimated leaves, maintenance
respiration relative to photosynthetic capacity still appears to be a rather low cost measured
Francisco Pugnaire/Functional Plant Ecology 7488_C007 Final Proof page 226 30.4.2007 7:57pm Compositor Name: DeShanthi
226 Functional Plant Ecology
relative to photosynthetic capacity and perhaps not much different between shade- and
sun-acclimated leaves (Sims and Pearcy 1992).
Of more importance than maintenance respiration to the carbon balance of leaves and the
payback time for a positive return on investment may be the construction costs, which occur
principally early in the leaf ’s development, but which must be paid back over the leaf lifetime.
On a per unit area basis, sun leaves are more expensive to construct because they are thicker
and have higher per unit area concentrations of photosynthetic enzymes. On a per unit mass
basis there is on average only a slightly greater (3%) construction cost of sun as compared
with shade-acclimated leaves (Poorter et al. 2006). Construction costs depend on the com-
position of the leaf and vary depending on the amounts of more costly, reduced compounds
such as soluble phenolics, lipids, proteins, and lignin as compared with carbohydrates. Sun
leaves can recover construction costs within a few days, whereas shade leaves may require
60–150 or more days, depending on the light environment (Chabot and Hicks 1982, Jurik and
Chabot 1986, Sims and Pearcy 1992).
Trade-Offs and Optimality in Sun Shade Acclimation
Modeling approaches have been used that explore the trade-offs involved in acclimation with
respect to whether they maximize the return on investment in a given light environment. At
the leaf level, one trade-off is how a given amount of biomass should be invested in either
increasing the area of the leaf, thereby enhancing light capture, or in increasing leaf thickness
thereby increasing the Amax
area
of the leaf. A second trade-off involves how N should be
partitioned within the photosynthetic apparatus in a given light environment to maximize
photosynthesis. This involves a trade-off between investment of N to maximize photo-
synthetic rate in high light versus investment in pigment–protein complexes that increase
the slope of the light response curve in low light (Evans 1989b). Several studies have shown
that nitrogen partitioning between light capture components (Chl-proteins) and compo-
nents that increase photosynthetic capacity (Rubisco and electron transport carrier com-
ponents) change with acclimation in a way that almost maximizes photosynthesis (Evans
1993, Hikosaka and Terashima 1995, 1996, Niinemets et al. 1998b). Moreover, daily carbon
gain is quite sensitive to the changes in allocation observed in high versus low light-acclimated
leaves, each with 20%–30% lower daily carbon gain in the light environment opposite to the
one that they were grown in (Hikosaka and Terashima 1996). Central to these optimizations
are first that light absorption is a hyperbolic function of chlorophyll content in agreement
with observed changes (Gabrielsen 1948) and second that relatively small changes in the
fraction of N allocated to pigment–protein complexes have a large effect on light absorption.
Comparisons of the assimilation rates per unit leaf area at the simulated optimum partition-
ing to the actual measured assimilation rates revealed that the latter tracked the former quite
closely with deviations only in the highest and lowest light environments. The changes for
shade and sun species did not track in the highest and lowest light environments, respectively.
Thus, nearly optimal partitioning could be maintained over a range of light environments, but
this range differed between shade and sun species.
Changes in LMA mediate the other trade-off in that a given amount of biomass invested
in a leaf can either be spread over a small area increasing its Amax
area
or over a large area
increasing its light capture. Sims et al. (Pearcy and Sims 1994, Sims et al. 1994) modeled the
sensitivity of whole-plant growth rate to changes in Amax
area
and LMA in Alocasia macro-
rrhiza. When just LMA was varied RGR was more sensitive to changes in LMA in the low-
light environment than in the high-light environment. Conversely, when just Amax
area
was
varied RGR was much less sensitive to changes in the low-light environment as compared
with the high-light environment. When LMA and photosynthetic capacity were varied
simultaneously, as occurs in acclimation, then RGR was much more sensitive in low as
Francisco Pugnaire/Functional Plant Ecology 7488_C007 Final Proof page 227 30.4.2007 7:57pm Compositor Name: DeShanthi
Responses of Plants to Heterogeneous Light Environments 227
compared with high light. These simulations emphasize the central role of changes in LMA in
acclimation to sun and shade and especially in improving performance in low light. This is
further emphasized in simulations where changes in N partitioning and in LMA characteristic
of acclimation were compared (Evans and Poorter 2001). This comparison revealed that daily
photosynthesis per unit dry mass is far more sensitive to changes in LMA than to changes in
allocation of N.
PLASTICITY OF WHOLE-PLANT RESPONSES
Integration of the role of photosynthetic plasticity in enhancing plant function in hetero-
geneous light regimes requires an understanding of how leaf photosynthetic rates interact
with and are determined by allocation patterns at the whole-plant level. Since the carbon
content of biomass is nearly constant, growth and whole-plant photosynthesis are indeed
closely linked by the conversion efficiency of fixed carbon into biomass (Dutton et al. 1988).
Whole-plant photosynthesis is simply the photosynthetic rate of the leaves integrated over the
leaf area. Of course, it is necessary to take into account that leaves are in different light
microenvironments within the crown and, because of acclimation and aging phenomena, may
have different capacities to use this light. Moreover, the carbon losses via respiration at the
whole-plant level must be subtracted to arrive at a whole-plant carbon balance.
Enhanced whole-plant photosynthesis can therefore be obtained either by increasing the
photosynthetic rate per unit area under the prevailing conditions, or by increasing the leaf
area per plant. An important linkage in this process is the LMA since this determines how
much leaf area is produced for a given investment of biomass in leaves. Most studies have
found that the leaf weight ratio (LWR: mass of leaves per unit mass of plant) tends to be
higher in sun as compared with shade-grown plants but the differences are usually rather
small (Bjo
¨
rkman 1981, Poorter and Nagel 2000). Instead, shade-grown plants invest more in
stems and less in roots with little change in total leaf area (Reich et al. 1998). Greater
investment in stems may be helpful in minimizing self-shading. Although the proportional
biomass investment in leaves tends not to vary much with sun-shade acclimation, shade-
grown plants typically have a much larger leaf area ratio (LAR: area of leaves per unit mass
of plant) than sun plants because of the large differences in LMA. Since, as discussed earlier,
LMA is also a determinant of Amax
area
, it clearly mediates a trade-off between lower
Amax
area
and greater leaf area for light capture in the shade, versus higher Amax
area
but
less leaf area in the sun. The result in the shade is a large advantage in whole-plant carbon
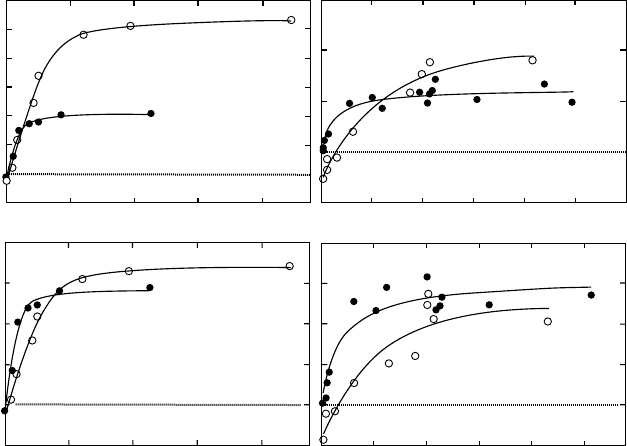
gain for the shade plant (Figure 7.4). Perhaps counterintuitively, experiments and simulations
have shown no carbon gain advantage per unit biomass invested for the sun plant phenotype
in high light since the reduced leaf area per plant more than offsets the advantage accrued due
to the higher Amax
area
(Sims and Pearcy 1994, Sims et al. 1994). Indeed, shade-acclimated
sunflower plants have been shown to have higher relative growth rates immediately after
transfer to high light than do plants grown continuously in the high-light environment (Hiroi
and Monsi 1963). Subsequent acclimation of the shade plant to the sun environment therefore
actually causes the RGR to decrease. This does not mean that sun leaves provide no
advantage in high light, especially over the long term, since they may well contribute to a
greater stress resistance and greater water and nitrogen use efficiency.
RESPONSES TO WITHIN-CROWN SPATIAL HETEROGENEITY
Within a plant crown, growth itself generates a heterogeneous light environment as younger
branches and leaves overtop and shade lower branches and older leaves. This gradient has
been studied in tree canopies (Niinemets et al. 1998b, Ellsworth and Reich 1993), herbaceous
Francisco Pugnaire/Functional Plant Ecology 7488_C007 Final Proof page 228 30.4.2007 7:57pm Compositor Name: DeShanthi
228 Functional Plant Ecology
vegetation (Anten et al. 1995, Hirose et al. 1988), and crops (Gutschick and Wiegel 1988) and
occurs both in closed canopies and in the crowns of isolated plants (c.f. Caldwell et al. 1986).
In all, the gradient in light is primarily a function of the cumulative leaf area index and the
architecture of the leaf and branch elements that controls the aggregation, inclination, and
self-shading of leaves within and between shoots. Response to this highly dynamic gradient
involves both senescence of older, more shaded leaves with reallocation of resources to
the newly developing leaves in the higher-light environments as well as acclimation to the
changed light environment. The modular nature of plants and the semiautonomy of these
modules confers great flexibility to plants in responding to fine-grained differences in light
availability (de Kroon et al. 2005).
Studies of the consequences of light attenuation within plant crowns have focused on the
question of how resources, in particular nitrogen and also LMA, should be distributed
between leaves at different positions in the crown to maximize photosynthesis. Optimal N
use occurs when canopy photosynthesis is maximized for a given whole-canopy N content
(Field 1983, Hirose and Werger 1987a). Optimal N use demands that more N be partitioned
to the upper leaves located in high light, resulting in a higher Amax
area
of these leaves as
compared with lower leaves in more shaded microenvironments. Differences in LMA have
been shown to be central to this partitioning because of its pivotal role in determining
photosynthetic capacity and N contents in sun and shade leaves. The optimal N distribution
between leaves is the one that results in a gradient of photosynthetic capacity through the
Instantaneous assimilation rate
single leaf
12
4
3
2
1
0
−1
0.3
0.2
0.1
0.0
−0.1
10
8
6
4
2
0
0.4
0.3
0.2
0.1
0.0
0 400 800 1200 1600 0 5 10 15 20 25
−0.1
−2
(a)
(c) (d)
(b)
Daily assimilation integral
whole plant
PFD (mol m
−2
day
−1
)
Assimilation (mol m
−2
day
−1
)
Assimilation (µmol m
−2
s
−1
)
Assimilation (µmol g
−1
s
−1
)
Assimilation (mol g
−1
day
−1
)
PFD (µmol m
−2
s
−1
)
FIGURE 7.4 Response of instantaneous assimilation rates to PFD for leaves (a,c) and daily assimilation
rates of whole plants (b,d) of Alocasia macrorrhiza acclimated to sun (open circles) and shade (closed
circles) conditions. The curves for whole plants were obtained by growing individual plants in sun or
shade conditions and then transferring to a whole-plant chamber for gas exchange conditions. Different
daily PFDs were obtained with neutral-density screen and cloth filters under natural sunlight. (Adapted
from Sims, D.A. and Pearcy, R.W., Plant, Cell Environ., 17, 881, 1994. With permission.)
Francisco Pugnaire/Functional Plant Ecology 7488_C007 Final Proof page 229 30.4.2007 7:57pm Compositor Name: DeShanthi
Responses of Plants to Heterogeneous Light Environments 229