Pugnaire F.I. Valladares F. Functional Plant Ecology
Подождите немного. Документ загружается.

NUTRIENT UPTAKE KINETICS: BASIC PRINCIPLES
Nutrient uptake is determined by both supply and demand at the root surface. Nutrients
arrive at the root surface by the mass flow of water toward the root, which is driven by
transpiration. Plants deplete the soil solution near the roots, when the nutrient uptake rate
exceeds the rate at which nutrients arrive. By doing so they create concentration gradients
around the roots that trigger diffusion of nutrients toward the root surface, which adds to the
supply by mass flow. When the depletion at the root surface proceeds, uptake must come in
pace with the supply rate. When the supply by mass flow exceeds the demand, nutrients (and
other solutes) can either be excluded, accumulating near the root, or enter the root and
accumulate in the plant to concentrations that may eventually become deleterious (Marschner
1995, Fitter and Hay 2002).
Depletion and accumulation at the root surface can occur simultaneously for different
elements. Table 8.1 gives an indication of the relation between the demand of the major
nutrients and their supply by mass flow. The listed concentrations in plant biomass are
considered to be sufficient for adequate growth (Epstein 1965). There is a close relation
between biomass production, nutrient demand, and water uptake. Based on the amount of
water transpired during the production of a unit biomass (transpiration coefficient), we can
calculate the nutrient concentration in the soil solution that would satisfy the nutrient
demands as listed in the first column by means of mass flow alone. Actual concentrations
in the soil solution of an average agricultural soil illustrate that mass flow rates of S, Mg, and
Ca amply exceed the demand, whereas mass flow of P falls entirely short of the demanded rate
of supply. In agricultural soils, mass flow rates of N and K are usually sufficient, but in most
natural soils, concentrations of N, P, and K are much lower, so that the supply by mass flow
alone is insufficient to satisfy the demand. Consequently, diffusion must play an important
role in the supply of these nutrients to plants growing on natural soils. This calls for a root
system that has the ability to take up nutrients selectively against a concentration gradient.
Selective uptake and transport through cell membranes is an energy-demanding process.
Passive, nonselective uptake without energy expenses is only possible where nutrients do not
have to pass a cell membrane on their way to the vascular cylinder of the root. Passive uptake
can revert into efflux when the concentration in the soil solution falls below the concentration
inside the root. Passive cation uptake through cell membranes can also proceed against a
TABLE 8.1
Average Nutrient Concentrations in Plant Biomass (Epstein 1965),
Concentrations in Soil Solution Required to Satisfy the Demand by Mass Flow,
Assuming the Transpiration Coefficient is 0.3 dm
23
g
21
d.wt., and Actual
Concentrations in the Soil Solution in an Arable Field
Element Concentration in Plant
Biomass (mmol kg
21
d.wt.)
Sufficient Concentration in
Mass Flow (mM)
Actual Concentration in Bulk
Soil Solution (mM)
N 1000 3.33 3.1
K 250 0.83 0.5
Ca 125 0.42 1.7
Mg 80 0.27 0.5
P 60 0.20 0.002
S 30 0.10 0.6
Source: After Peters, M. in Schriftenreihe des Institutes fu
¨
r Pflanzenerna
¨
hrung und Bodenkunde, H.P.
Blume, ed., Universita
¨
t Kiel, Kiel, 1990.
Francisco Pugnaire/Functional Plant Ecology 7488_C008 Final Proof page 260 30.4.2007 7:58pm Compositor Name: DeShanthi
260 Functional Plant Ecology

concentration gradient, because cells can create an electrochemical gradient by actively
pumping out protons across the membrane. Anions, on the other hand, have to be trans-
ported actively through cell membranes by means of carrier enzymes.
Passive uptake without energy expenses would be sufficient for nutrients that are required
in low quantities and occur in relatively high concentrations in the soil solution, if it were not
for the closed structure of the root. Since passive uptake is not selective and cannot be
regulated, plants that rely too much on passive uptake can easily be overloaded with nutrients
and toxic ions when concentrations in the soil solution are high. This makes it understandable
why plant roots possess an endodermis that prevents passive nutrient transport (see Chapter
5, this volume). Solutes can enter the root via the apoplastic pathway between the cortex cells,
but no further than the endodermis with its bands of Caspari. Solutes that are transported by
mass flow to and into the root at a higher rate than the active uptake rate by rhizodermal,
cortex, or endodermis cells accumulate between the cortex cells and at the root surface. This
leads to diffusion in the direction opposite to the water flow, away from the root, back into
the soil. Most nutrients enter the root across a cell membrane somewhere in the cortex by
means of active transport and continue their way inside along the symplastic pathway, from
one cell to another via intercellular cytoplasma connections (plasmodesmata), to pass
through the endodermis and finally enter the vascular cylinder (Marschner 1995). A small
fraction of the nutrients can circumvent the endodermis at the root tip where the bands of
Caspari are not yet formed, and at places where the endodermis is pierced by lateral roots,
torn, or damaged otherwise.
Active uptake against a concentration gradient, by means of an energy-demanding
process, is the predominant uptake process for the major nutrients N, P, and K. The uptake
rate depends on the nutrient concentration at the root surface. Usually an asymptotic relation
is found between the concentration in a nutrient solution and the uptake rate, when measured
in short-term experiments with excised roots from plants that have been deprived of nutrients
for a few weeks. The uptake as a function of the concentration in the surrounding solution is
generally described by:
V ¼ V
max
C
l
C
l
þ K
m
,
in which V is the gross uptake rate (mmol g
1
fw h
1
), V
max
is the maximum uptake rate,
C
l
is the nutrient concentration in the soil solution at the root surface (mM), and K
m
is the
Michaelis constant, which is the value of C
l
where V is half V
max
. The Michaelis–Menten
equation is typical for the kinetics of enzymatic processes and reflects the fact that the
carrier enzymes in the cell membranes become saturated with increasing nutrient concen-
tration. K
1
m
expresses the affinity of the carriers for the substrate ion (i.e., the slope of the
curve in the origin, which is measured by V
max
=K
m
). Although different nutrient ions are
transported by different carriers, the selectivity of the carriers is not perfect. Different
nutrients may compete for the same carriers, so that K
m
may be increased by the presence
of other ions with the same electrical charge.
The maximum uptake rate (V
max
) is realized when the concentration (C
l
) is so high that all
carrier enzymes are continuously occupied with a substrate ion. V
max
depends on the density
and the activity of carriers in the cell membranes. It depends also on the internal nutrient
status of the plant, because the activity of the carriers can be suppressed by high nutrient
concentrations in the root (compare V
max
under deprived and well-fed conditions, Figure 8.1).
This negative-feedback control operates when plants are growing under nutrient-rich condi-
tions. It can reduce V
max
to less than 20% of its value in a deprived plant (Loneragan and
Asher 1967) and prevents accumulation of too high nutrient concentrations in the root. On
the other hand, it has been found that plants growing under nutrient poor conditions can
Francisco Pugnaire/Functional Plant Ecology 7488_C008 Final Proof page 261 30.4.2007 7:58pm Compositor Name: DeShanthi
Acquisition, Use, and Loss of Nutrients 261
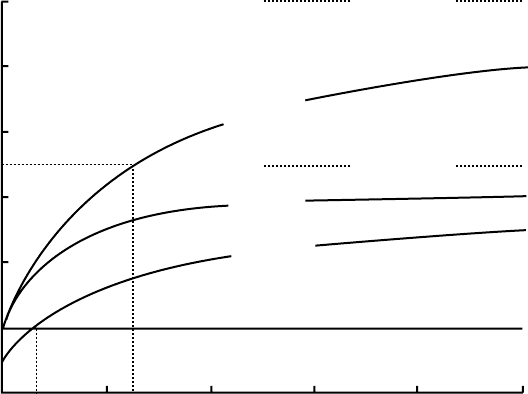
temporarily increase V
max
when the concentration in the soil is increased (Lefebvre and Glass
1982, Jackson et al. 1990).
Consequently, roots of a well-fed plant are operating far below their maximum uptake
capacity. When the soil becomes depleted and the nutrient concentration in the plant starts to
drop, the negative-feedback control on V
max
is relaxed, so that V
max
increases, which com-
pensates for the decrease in supply rate. The responses of V
max
to changes in internal and
external nutrient concentrations allow a plant to regulate its nutrient uptake and rapidly use
temporarily high nutrient concentrations that may occur locally in a predominantly poor soil.
Under rich conditions, it allows a plant to maintain its overall uptake rate even when a large
part of the root system is removed.
Besides uptake, efflux of nutrients may occur. When active uptake takes place, nutrient
concentrations inside the root are usually higher than those outside the root. Since roots are not
perfectly closed, leakage of nutrients can reduce the net uptake rate (V
net
). When nutrient influx
and efflux occur simultaneously, the plant can only decrease the nutrient concentration in the
soil solution until it reaches a minimum concentration (C
min
) at which influx and efflux are
equal. At values of C
l
lower than C
min
, the efflux is larger than the gross influx, so that the net
influx is negative and the roots lose nutrients to the solution until C
l
equals C
min
(Figure 8.1).
Like V
max
, C
min
is not a constant. Under nutrient poor conditions, when nutrient
concentrations inside the root are low, the efflux is also low. Together with the release of
the feedback control on V
max
, this results in very low values of C
min
under nutrient-poor
conditions (Figure 8.1). It is unclear at present whether the K
m
value is also able to respond to
changes in internal or ambient nutrient concentrations (Marschner 1995). The reported
10
8
6
4
2
0
0.00
Concentration at the root surface C
1
(mM)
Net uptake rate V
net
(µmol g
−1
fw h
−1
)
0.02
C
min
K
m
0.04
Long term
Short term
deprived
V
max
deprived
V
max
well fed
Short term
well fed
0.080.06 0.10
−2
FIGURE 8.1 Relation between external nutrient concentration at the root surface (C
l
) and net nutrient
uptake rate (V
net
¼VE). The upper curve represents the short-term uptake by excised roots from
previously deprived plants (V
max
¼10, K
m
¼0.025, efflux ¼0). The lower curve represents short-term
uptake by excised roots of previously well-fed plants (V
max
¼5, K
m
¼0.025, efflux ¼1). C
min
is the
concentration at which V
net
¼0. The curve marked long term represents nutrient uptake by whole plants
grown for several weeks on nutrient solutions with constant concentration, so that V
max
and efflux are in
steady state with the internal nutrient concentration and C
l
.
Francisco Pugnaire/Functional Plant Ecology 7488_C008 Final Proof page 262 30.4.2007 7:58pm Compositor Name: DeShanthi
262 Functional Plant Ecology

changes in uptake kinetics of roots in response to localized nutrients (e.g., Drew and Saker
1975, Jackson et al. 1990) may be due to changes in V
max
or efflux rate alone. In most plant
species, the value of K
m
for N, P, and K is so low that the concentration of these nutrients at
the root surface can become virtually zero, for example, 0.35 mM for NO
3
(Freijsen et al.
1989), 1 mM for K
þ
(Drew et al. 1984), and less than 0.01 mM for H
2
PO
4
(Breeze et al. 1984).
NUTRIENT ACQUISITION IN SOILS
The description of active nutrient uptake given earlier applies mainly to uptake by single roots
in a well-mixed nutrient solution. However, the relation between uptake and concentration in
the soil solution is of little consequence for the overall nutrient uptake by a whole plant
growing in a poor soil. In soils, uptake of N, P, and K is almost always limited by the rate
of transport toward the root surface and not by the capacity of the uptake mechanism.
Concentrations of these nutrients in the soil solution are often so low, and the uptake is so
efficient that all available nutrients near the root surface can be taken up within a few
minutes. When nutrient uptake is not immediately compensated by nutrient transport from
the bulk soil toward the root surface, the nutrient concentrations at the root surface fall and
the uptake rate decreases. Even in nutrient solutions, where transport rates (TRs) are high,
depletion at the root surface may reduce nutrient uptake rates, as appears from the stimulating
effect of stirring (Freijsen et al. 1989). The importance of nutrient transport to the root in
nutrient-poor soils is illustrated by the following analysis of the balance between nutrient
supply and uptake at the root surface (Nye and Tinker 1977).
As explained earlier, nutrient uptake is determined by the concentration at the root
surface (C
l
), which, in its turn, is the resultant of nutrient transport to the root and the net
nutrient uptake rate (V
net
). The transport rate is the sum of the mass flow rate and the
diffusion rate (DR). The mass flow rate is the product of water flow (V
w
) and nutrient
concentration in the bulk of the soil solution (C
b
). The diffusion rate is the product of the
concentration gradient toward the root surface (dC=dx) and the effective diffusion coefficient
(D
e
). The effective diffusion constant, in turn, depends on the moisture content of the soil, the
tortuosity of the diffusion pathway, and the buffer power of the soil, which accounts for the
degree to which nutrient transport is impeded by interaction with soil particles. Phosphate is
strongly adsorbed to soil particles, which leads to low values of C
b
resulting in low rates of
mass flow and diffusion. At the other end of the spectrum, nitrate is much more mobile,
because adsorption is negligible. Potassium takes an intermediate position.
The net uptake rate (V
net
) is calculated as the gross uptake rate (V ), minus the efflux rate
(E ). This leads to the following equations:
TR ¼ V
w
C
b
þ D
e
dC
dx
,
V
net
¼ V
max
C
l
C
l
þ K
m
E:
When the transport rate equals the net uptake rate, a dynamic equilibrium develops (with
equilibrium concentration and uptake rate C
l
*andV
net
*). When C
l
is lower than C
l
*, the
transport rate is higher than the net uptake rate, so that C
l
increases until it reaches C
l
* (cf.
Figure 8.2). When C
l
is higher than C
l
*, the transport rate is lower than the net uptake rate, so
that C
l
decreases until it reaches C
l
*. At a short term (hours), the equilibrium concentration
(C
l
*) and the corresponding equilibrium uptake rate (V
net
*) are stable. When uptake proceeds
for a few days, the equilibrium shifts gradually to lower C
l
values, because the depletion zone
grows, the diffusion gradient becomes less steep, and the transport rate decreases. Around
thin roots, depletion proceeds slower than around thick roots, because of the radial geometry
Francisco Pugnaire/Functional Plant Ecology 7488_C008 Final Proof page 263 30.4.2007 7:58pm Compositor Name: DeShanthi
Acquisition, Use, and Loss of Nutrients 263
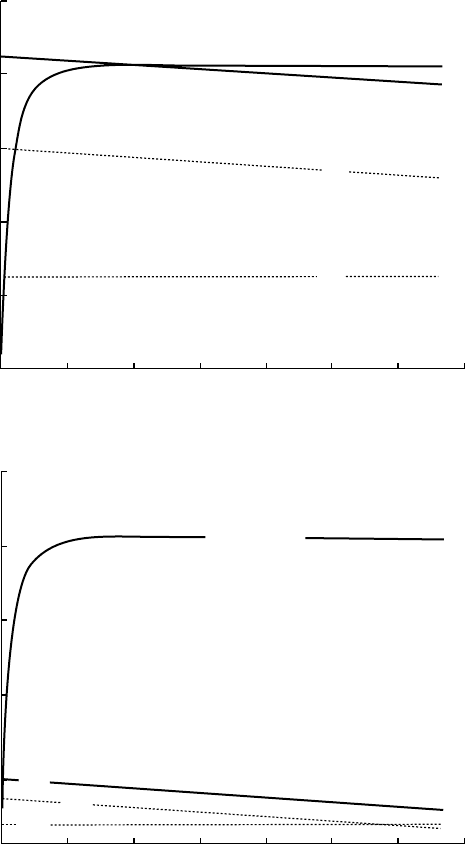
of roots. The amount of soil per unit root surface present within the same distance from the
root surface is larger (and contains more nutrients) when the root diameter is smaller.
The depletion zone around the root continues to grow until the transport rate becomes so
low that it equals the rate at which nutrients are released from the soil within the depletion
zone by means of dissolution, desorption, or mineralization. Immobile nutrients, like
V
net
long term
TR
DR
MR
5
4
3
2
1
0
5
V
net
long term
4
3
2
1
0
TR
DR
MR
0.0 0.1 0.2 0.3
Uptake V
net
or transport (µmol g
-1
fw h
-1
)
Concentration at the root surface C
1
(mM)
0.4 0.5 0.6 0.7
(a)
(b)
Concentration at the root surface C
1
(mM)
Uptake V
net
or transport (µmol g
-1
fw h
-1
)
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7
FIGURE 8.2 Net uptake rate (V
net
) and nutrient transport rate to the root surface (TR) as a function of
the nutrient concentration at the root surface (C
l
). The diffusion rate (DR), the mass flow rate (MR),
and their sum (TR) at (a) high (C
b
¼5) and (b) low concentrations (C
b
¼0.3) in the bulk soil. In either
case, the equilibrium concentration (C
1
*) and equilibrium uptake rate (V
net
*) are found at the intersec-
tion of the lines V
net
and TR. The parameter values used are hypothetical, keeping midway between
NO
3
and K
þ
. V
net
is identical to the long-term uptake in Figure 8.1.
Francisco Pugnaire/Functional Plant Ecology 7488_C008 Final Proof page 264 30.4.2007 7:58pm Compositor Name: DeShanthi
264 Functional Plant Ecology
phosphate, are released slowly and have narrow depletion zones, low uptake rates, and low
C
l
*. Eventually, when the nutrients adsorbed to soil particles are depleted up or when
mineralization is interrupted due to low temperatures, the release rate falls and uptake stops.
To maximize uptake, plants can reduce the distance over which nutrients are transported
through the soil, by increasing the density of their root system. Plants can realize higher root
densities by increased allocation of carbon to their rooting system but also by reduced root
diameters which leads to an increased root length and root surface per unit of root biomass.
Root hairs are important in this respect, because they are thin and require little investment of
biomass per unit of soil explored. However, their vulnerability and short life span make them
less profitable when the bulk of the soil is already depleted, so that a plant has to ‘‘sit and
wait’’ for nutrients that are released from the solid phase. In such nutrient-poor situations
many plant species are living in symbiosis with fungi that form mycorrhizas. Such associ-
ations strongly increase the total surface area by which nutrients can be taken up. Fungal
hyphae have much smaller diameters than roots (see Chapter 5, this volume).
Silberbush and Barber (1984) have studied the effect of changes in plant and soil charac-
teristics on the equilibrium uptake rate of plants growing in soil. Figure 8.2 illustrates their
results. The equilibrium uptake rate (V
net
*) and nutrient concentration (C
l
*) at the root
surface are given by the intersection of the lines TR and V
net
. When nutrient concentrations
in the soil are high (Figure 8.2a, with C
b
¼5), C
l
* is situated at the horizontal part of the
uptake curve. In this case, the equilibrium uptake rate V
net
* is determined largely by the
maximum uptake capacity of the roots (V
max
) and not by the affinity of the uptake mechan-
ism (K
m
), nor by the mass flow or diffusion rate. Consequently, we may expect that natural
selection on rich soils will favor plants with a high maximum uptake capacity (V
max
).
When nutrient concentrations in the bulk soil are low (Figure 8.2b, with C
b
¼0.3), C
l
*is
situated at the ascending part of the uptake curve. Here, V
net
* is mainly determined by the
effective diffusion coefficient D
e
and by C
b
, which determine the slope of the line that
represents the diffusion rate and the intercept with the horizontal axis, respectively. In this
case, the value of V
net
* is relatively insensitive to changes in the kinetic parameters that rule
the uptake process (K
m
and V
max
). Consequently, we may expect that natural selection of
plants on poor soils will not lead to increased affinity or capacity of the nutrient uptake
mechanism, but to properties that reduce the transport limitation, bringing the root surface
closer to the nutrients (i.e., by fine and dense root systems and mycorrhizal associations). By
increasing the root surface per unit plant biomass, a plant can sustain adequate growth rates
with lower nutrient uptake rates per unit root surface and thus with lower nutrient concen-
trations at the root surface than competitors with a smaller root system.
The nutrient concentration in the bulk of the soil solution (C
b
) can differ dramatically from
the concentration at the root surface (C
l
), implying that C
b
is not a good indicator of nutrient
availability. When the nutrient pool in the bulk of the soil solution is depleted, the nutrient
supply to the root depends on the rate at which available forms of the nutrient are released from
the organic and the mineral substrates. Plants can increase the release rate of nutrients by
lowering the concentration in the soil solution or by affecting the chemical conditions or the
microbial activity in the rhizosphere. Some species can use chemical forms or physical states
(solid, dissolved, adsorbed, or occluded) of a nutrient that other species cannot use. Nitrogen,
for example, can be taken up by most species only as NO
3
and NH
4
þ
, but some species can also
take up amino acids and other dissolved organic molecules that contain nitrogen (Kielland
1994, Northup et al. 1995, Schimel and Chapin 1996, Leadley et al. 1997). Some species can
mobilize iron in calcareous soils by lowering the pH or by exudating chelating or reducing
substances (Ro
¨
mheld and Marschner 1986). Phosphorus can be taken up by most species only
as H
2
PO
4
, but some species are able to mobilize solid calcium phosphate by changing the
chemical conditions in the rhizosphere, for example, by lowering pH, exudation of organic
acids, or lowering the Ca concentration in the soil solution (Hoffland 1992). The ability to
Francisco Pugnaire/Functional Plant Ecology 7488_C008 Final Proof page 265 30.4.2007 7:58pm Compositor Name: DeShanthi
Acquisition, Use, and Loss of Nutrients 265
mobilize insoluble nutrients by altering the chemical conditions in the rhizosphere can be
enhanced by forming dense clusters of lateral roots that intensify the rhizosphere effects.
These morphological adaptations are called proteoid roots, after the Proteaceae family, but
they occur also in species of other taxa (e.g., Lupine). An excellent review of the ability of plant
species to use alternative nutrient sources is given by Marschner (1995).
Many plant species from nutrient-poor soils have special adaptations that enable them to
use nutrients from other sources than the soil solution. The most widespread of these are
associations with mycorrhizal fungi and symbiosis with N-fixing bacteria such as Rhizobium
and Frankia. Mycorrhizal fungi are able to decompose dead organic material and to transport
the mineralized nutrients to the plant root (see Chapter 5, this volume). Recently, Jongmans
et al. (1997) suggested that mycorrhizal fungi are also able to penetrate rocky materials and to
absorb P, Mg, Ca, and K from minerals by the excretion of organic acids and to transport
these nutrients to connected roots. More peculiar adaptations that occur mainly in extremely
nutrient-poor ecosystems are parasitism on other plants (e.g., Rhinanthus, Pedicularis) and
carnivory (e.g., Drosera, Pinguicula, Utricularia).
UPTAKE OF ORGANIC NITROGEN COMPOUNDS
A few decades ago, it was assumed that most plant species absorbed nitrogen as nitrate and
ammonium except for species with ecto- or ericoid mycorrhizal associations (Read 1991). It
was long considered that the mineralization of organic nitrogen to ammonium and its subse-
quent oxidation were the major bottlenecks restricting the nitrogen supply to plants (Chapin
1995). However, these ideas were strongly disturbed by the observation that measured rates of
net microbial production of inorganic nitrogen were often less than half the observed rates
of nitrogen acquisition by plants (Fisk and Schmidt 1995, Kaye and Hart 1997). In the 1990s, it
became clear that quite a few species can absorb amino acids from solution and can easily
survive when no other nitrogen forms are supplied (Chapin et al. 1993, Kielland 1994). Schimel
and Chapin (1996) showed that two tundra sedges, Eriophorum vaginatum and Carex aquatilis,
which were unlikely to have any of these mycorrhizal associations, nevertheless absorbed
amino acids (glycine and aspartate) under field conditions. The amino acids that they provided
to the plants were labeled with
15
N and
13
C to test whether complete amino acids were taken up
or that ammonium that was derived from decomposing amino acid molecules was absorbed.
The authors did not detect any of the
13
C in the produced plant tissues and they attributed this
failure to respiration of the labeled C atoms. Glycine and aspartate can be easily converted into
glycolysis or TCA cycle intermediates and subsequently respired. Na
¨
sholm et al. (1998) solved
this problem by
15
N and dual
13
C labeling of the glycine molecule. Here both C atoms were
labeled instead of only the carbon in the carboxyl group, preventing that all
13
C were rapidly
respired after decarboxylation. Their measurements in a boreal forest showed that at least 91%,
64%, and 42% of the nitrogen from the absorbed glycine was taken up as intact glycine
molecules in the dwarfshrub Vaccinium myrtillus, the grass Deschampsia flexuosa, and the
trees Pinus sylvestris and Picea abies, respectively. These results showed unambiguously that
these different species, irrespective of their completely different mycorrhizal associations, can
bypass nitrogen mineralization. The dwarfshrubs and trees have ericoid and ectomycorrhizal
associations and were expected to absorb organic nitrogen. But it was surprising that the
arbusco-mycorrhizal grass species also appeared to be able to absorb amino acids. A recent
review (Aerts and Chapin 2000) mentioned that ‘‘the ability to take up organic N sources . . .
hardly occurs in species with arbuscular mycorrhizas.’’ But it seems that the ability to absorb
dissolved organic nitrogen compounds is much more widespread among plant species than we
earlier believed (Schmidt and Stewart 1997, Lipson et al. 1999, Persson et al. 2003). Neverthe-
less, the quantitative significance of the uptake of dissolved organic nitrogen as compared with
the uptake of ammonium and nitrate is still to be assessed.
Francisco Pugnaire/Functional Plant Ecology 7488_C008 Final Proof page 266 30.4.2007 7:58pm Compositor Name: DeShanthi
266 Functional Plant Ecology
ROOT FORAGING IN HETEROGENEOUS ENVIRONMENTS
So far we have examined nutrient uptake and transport in a homogenous substrate. However,
nutrient availability in soils may vary dramatically beyond the zone of influence of the roots
themselves. Soil patches of different quality are created at various scales by abiotic factors
(soil type differences, soil depth, microtopography) as well as by biotic factors such as
treefalls and stemflow in forests (Gibson 1988a,b, Hook et al. 1991, Lechowicz and Bell
1991, Farley and Fitter 1999). In arid environments, organic matter accumulates in the
vicinity of isolated trees, shrubs, and persistent turf grasses creating islands of fertility in a
nutrient-deprived matrix (Jackson and Caldwell 1993, Alpert and Mooney 1996, Ryel et al.
1996, Schlesinger et al. 1996). Consequently, from the point of view of the plant individual, in
many habitats the spatial distribution of water and nutrients is profoundly heterogeneous
from scales as small as a few centimeters, to tens of meters and more. How effectively can
plants capture the resources in such a heterogeneous world? What fraction of the growth
achieved at a homogeneous supply of soil resources can be realized when similar amounts of
resources are patchily distributed? Do species from habitats of different resource status have
different foraging abilities?
Especially for the less-mobile ions such as phosphate, pockets of nutrients may only be
captured by the plant if roots expand their surface area into the richer patch (Hutchings and
de Kroon 1994, Robinson 1994, 1996). This foraging behavior may be very effective, as is
perhaps best illustrated with the classical study by Drew and coworkers with barley (Hordeum
vulgare). Single root axes of barley were grown into three compartments in which the
concentration of nutrients could be controlled separately. High nitrate concentration in a
given compartment promoted the formation of more first- and second-order laterals per unit
of primary root length within that compartment and greater lateral root extension (Drew et al.
1973). When one-third of the entire root system received a nutrient-rich solution, total lateral
root length per unit of length of the primary axis was 10 times higher, and the total root
biomass 6 times higher, in the high-nutrient compartment than those in the low-nutrient
compartments. Later in the experiment, when the lateral roots had grown out, whole-plant
relative growth rate (RGR) under localized supply of nutrients approached the RGR of
control plants growing under homogeneous nutrient supply (Drew and Saker 1975). When
phosphate was supplied to 2 cm of the main root axis—a fraction amounting to only a few
percent of its total length—whole-plant RGR was more than 80% of its value in control
plants in which the whole root system received phosphate. When applied to 4 cm of the main
root axis, the RGR was similar to that of controls. The higher local nutrient uptake from
small pockets of nutrients to which part of the root system was exposed was not only due to
an enlargement of the local root surface area. In addition, phosphate absorption rates per unit
of root length increased in the enriched compartment, compared with both other parts of the
root system in treated plants and the root system of control plants (Drew and Saker 1975).
The enhanced formation of roots in nutrient hotspots is now referred to as root prolifer-
ation, selective root placement, or root foraging precision (de Kroon and Mommer 2006).
Local conditions determine where lateral root growth and uptake is promoted (Drew et al.
1973, Drew and Saker 1975), but the magnitude of the local response depends on the
conditions experienced by the rest of the root system and the entire plant. An experiment
by Drew (1975) illustrates this well. He subjected roots of barley plants to either a uniform or
localized nutrient supply. Part of the root system given a high phosphate supply produced
more and longer lateral roots when the rest of the root system was receiving low phosphate
rather than high phosphate (Figure 8.3). This suggests that the local morphological response
is stronger when phosphate is more limiting to the plant. Broadly similar effects were
produced when the nitrate and ammonium supply to different sections of the root system
was varied (Drew 1975). However, effects were less clear for nitrate (Drew et al. 1973,
Francisco Pugnaire/Functional Plant Ecology 7488_C008 Final Proof page 267 30.4.2007 7:58pm Compositor Name: DeShanthi
Acquisition, Use, and Loss of Nutrients 267
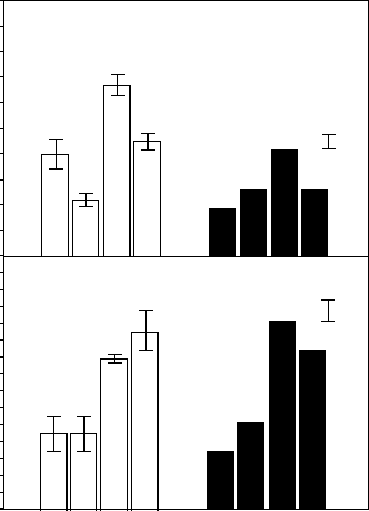
see Figure 8.3). In most studies in which nutrients were supplied heterogeneously, root
growth was suppressed in the part of the root volume that experiences low nutrient supply
(Robinson 1994).
The merits of the ability to forage for patchily distributed nutrients can perhaps best be
illustrated by comparing the biomass production of plants grown on homogeneous and
heterogeneous substrates each with the same overall nutrient availability. Fransen et al.
(1998) created such treatments by mixing poor riverine sand with black humus-rich soil either
homogeneously or by concentrating most of the black soil in a small column within the pot.
Five grass species were grown individually in each of these treatments. Their roots readily
reached and penetrated the enriched column but the responses were significant only for the
three species characteristic of relatively nutrient-rich habitats. Combined for all species,
whole-plant nutrient accumulation and biomass at the end of the experiment was significantly
higher in the heterogeneous treatment than that in the homogeneous treatment. These results
indicate that plant species may profit and grow faster at a heterogeneous distribution of soil
nutrients, rather than slower. In this experiment with bunchgrasses the growth stimulus in the
Phosphate
Nitrate
10
9
8
7
6
5
4
3
2
1
0
10
5
0
LLHH
LHLH LHLH
LLHH
Middle zone
Length (cm)
Number per cm
Adjacent zones
(a)
(b)
FIGURE 8.3 Effects of nitrate and phosphate supply on (a) the number of lateral roots per cm of main
root axis and (b) the lengths of individual lateral roots in barley. Main root axes were divided into three
zones and nutrients were supplied independently to each of these zones. Data given are those for the
first-order laterals that developed in the middle zone. This zone experiences either a low (L) or a high (H)
concentration of nitrate or phosphate. Adjacent rooting zones also grew in either a high- or low-nutrient
solution. In the nitrate experiment, plants were grown hydroponically; in the phosphate experiment,
they were grown in sand. Nitrate data are given as mean +SE, phosphate as means with separate bars
showing the LSD at the 5% level. (After Drew, M.C., Saker, L.R., and Ashley, T.W., J. Exp. Bot., 24,
1189, 1973; Drew, M.C., New Phytol., 75, 479, 1975; adapted from Hutchings, M.J. and de Kroon, H.,
Adv. Ecol. Res., 25, 159, 1994. Courtesy Academic Press. With permission.)
Francisco Pugnaire/Functional Plant Ecology 7488_C008 Final Proof page 268 30.4.2007 7:58pm Compositor Name: DeShanthi
268 Functional Plant Ecology
heterogeneous treatment compared with the homogeneous treatment was small. Clonal
species that spread horizontally have the ability to take up nutrients locally and produce
most of the biomass beyond the nutrient-rich patch. For such species, a several-fold increase
in biomass production may occur if the distribution of nutrients is not homogeneous, but
concentrated in small hotspots (Birch and Hutchings 1994, Hutchings and Wijesinghe 1997,
Wijesinghe and Hutchings 1997).
To what extent these results on root foraging ability in heterogeneous soils are generally
valid? In a recent meta-analysis covering the results of over 100 species, Kembel and Cahill
(2005) showed that the responses are very variable, confirming earlier overviews (Robinson
1994, Hodge 2004). Species varied from little or no root proliferation at all, distributing their
roots equally over the rich and poor parts of the soil, up to the very plastic responses as
observed for barley in Drew’s experiments. Species may also differ markedly in the time
between nutrient application and response. For example, when exposed to nutrient enrich-
ment, roots of the cold desert species Agropyron desertorum showed a fourfold increase in the
RGR of root length within one day, whereas Artemisia tridentata and especially Pseudor-
oegneria spicata responded less vigorously (Jackson and Caldwell 1989). In the latter species,
extension growth was not affected until several weeks after nutrient application.
For one of the three datasets analyzed, Kembel and Cahill (2005) found support for the
notion of Grime et al. (1986) that plant species with a higher RGR place their roots more
selectively in heterogeneous soils. When in a given experiment, plants of different growth rates
are harvested after a fixed period of time, as is usually the case, the degree of selective root
placement is indeed positively correlated to growth rate (Fransen et al. 1999, Aanderud et al.
2003). This correlation has its origin in the modular nature of the root system (sensu de
Kroon et al. 2005), in which roots respond locally to the nutrient concentrations that they
experience. Species with larger RGRs in terms of plant biomass are likely to also have larger
root RGRs (more lateral root formation and higher root extension rates) in the richer
microsites. However, when corrected for growth rate differences, the degree of selective
root placement is the same for slow-growing and fast-growing species (Fransen et al. 1999,
Aanderud et al. 2003).
However, it should be realized that species with higher root proliferation in enriched
patches do not necessarily obtain more nutrients from heterogeneous soil than species with
less root proliferation, unlike the results of Drew and Saker (1975) and Fransen et al. (1998)
suggest. For their larger datasets, Kembel and Cahill (2005) found no significant correlation
between the response to nutrient heterogeneity in terms of biomass production and the
precision by which the roots were placed in the nutrient-richer patches. Some of these variable
results may be explained by slow response of root proliferation that may come too late
relative to nutrient release in the patches (Robinson 1996, Van Vuuren et al. 1996), suggesting
a much more prominent role of enhanced maximum uptake capacity (i.e., physiological
plasticity) for the acquisition of finite nutrient patches. The gain in biomass in heterogeneous
versus homogeneous soils also becomes smaller when the experiments last longer because
patches deplete and the precision of root placement reduces (Kembel and Cahill 2005). This
makes sense because when all soil nutrients are taken up, no differences in biomass produc-
tion are to be expected between homogeneous and heterogeneous soils if in both treatments
the same total amount of nutrients is supplied. Despite these methodological caveats, our
current understanding is that the root proliferation in enriched microsites is less important for
nutrient acquisition in heterogeneous soils than previously thought (de Kroon and Mommer
2006), except when plants are in competition (Hodge et al. 1999, de Kroon et al. 2003; but see
Fransen et al. 2001).
To evaluate the ecological significance of selective root placement, its benefits must be
compared with its costs. The immediate benefits may be limited but if the costs of wrong
placement are small, selective root placement may still be profitable. Jansen et al. (2006)
Francisco Pugnaire/Functional Plant Ecology 7488_C008 Final Proof page 269 30.4.2007 7:58pm Compositor Name: DeShanthi
Acquisition, Use, and Loss of Nutrients 269