Санагурський Д.І. Об’єкти біофізики: монографія
Подождите немного. Документ загружается.

181
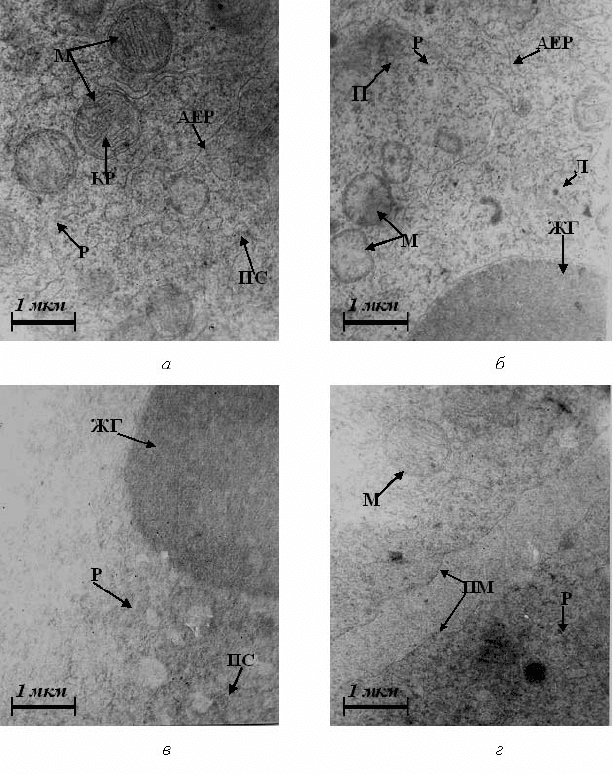
Рис. 4.16. Ультраструктура бластомерів зародків в’юна на шостій годині
розвитку за умов впливу катіонів марганцю. Позначення – як на рис. 4.1
У ділянках гіпертрофованого комплексу Гольджі виявлено
значні скупчення дрібних первинних лізосом, поодинокі первинні
лізосоми великого розміру та дезорганізовані мітохондрії
(рис. 4.17, г).
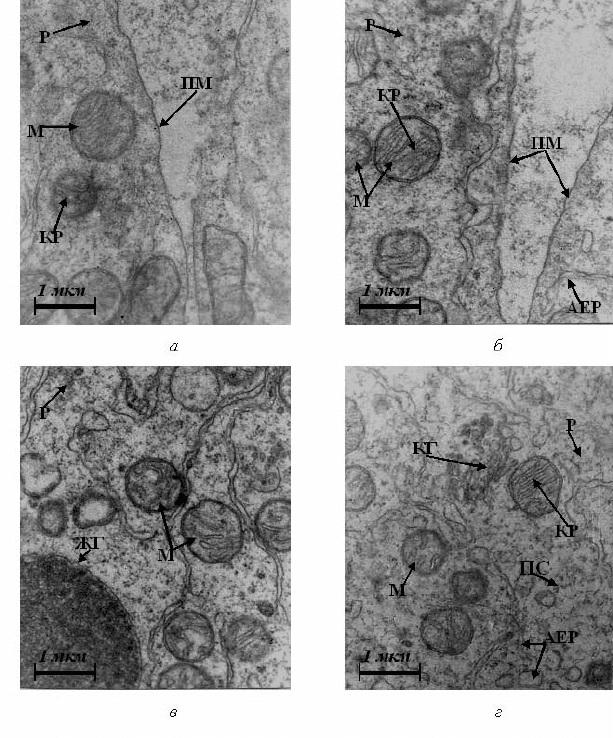
182
Рис. 4.17. Ультраструктура бластомерів зародків в’юна на шостій годині
розвитку за умов впливу катіонів кадмію. Позначення – як на рис. 4.1
Гіпертрофія комплексу Гольджі свідчить про підвищену
його діяльність, що направлена на утилізацію пошкоджених
ультраструктур та метаболітів.
Підсумовуючи, скажемо, що інкубування зародків в’юна на
різних стадіях розвитку за наявності катіонів важких металів
183
призводить до виникнення значних змін ультраструктури
бластомерів зародків, порівняно з контролем. Спостерігаються
такі зміни, як пошкодження та дезорганізування органел,
гіпертрофія видільної системи (комплексу Гольджі), значний у
деяких прикладах набряк цитоплазми та органел, розпушення та
пошкодження мітохондріальних і плазматичних мембран
(Н.М. Бойко та ін., 2002).
Зауважимо, що, незважаючи на такі значні зміни
ультраструктури бластомерів, зародки в’юна, що були інкубовані за
наявності катіонів досліджуваних важких металів, зберігали
життєздатність, хоча і відставали у розвитку від контрольних. На
більш пізніх стадіях (7–10 діб) у 25–30% з них спостерігали
наявність певних вад розвитку, малорухливість, сповільнення
серцебиття, але відсоток загибелі цих зародків був ненабагато
більший, ніж у контролі (Н.М. Бойко, О.А. Гойда, 1996;
Н.М. Бойко, Д.І. Санагурський, 2000).
Ðîçä³ë 5. ÀËÃÎÐÈÒÌÈ
ÏÎÁÓÄÎÂÈ «ËÎò×ÍÈÕ»
ÑÒÐÓÊÒÓÐ, ÑÕÅÌ, ßÊ ÀÍÀËÎÃ
Á²ÎËÎò×ÍÈÕ ßÂÈÙ
5.1 ßê³ñíèé òà ïîð³âíÿëüíèé
àíàë³ç äèíàì³êè
á³îåëåêòðè÷íèõ,
åíåðãîçàáåçïå÷óâàëüíèõ
³ ìåìáðàííî-òðàíñïîðòíèõ
ïðîöåñ³â ó ðàííüîìó
åìáð³îãåíåç³ â’þíà òà
øïîðöåâî¿ æàáè
З метою дослідження особливостей
динаміки біоелектричних і метаболічних процесів та з’ясування
часового організування між ними на стадії дроблення зародків риб
та амфібій нами проведено математичний аналіз аперіодичних та
періодичних змін параметрів, якими описують ці процеси.
Відомо, що часове організування полягає у сукупності всіх
ритмічних процесів, які відбуваються у біосистемах, що
185
взаємодіють, узгоджуються в часі між собою та зі змінними
умовами середовища (Ю.А. Романов, В.В. Маркина, 1989).
Вважають, що ритмічні зміни відтворюють діяльність регулятор-
них механізмів у підтриманні гомеостазу, тоді як аперіодичні
(зміни загального рівня) – діяльність, спрямовану на зміну рівня
функціонування різних процесів (Н.Б. Игошева и др., 2001).
Останнє твердження узгоджується з отриманими нами
результатами крос-кореляційного аналізу аперіодичних змін біо-
електричних і метаболічних характеристик зародків в’юна та
шпорцевої жаби.
Для зародків в’юна у першій серії крос-кореляційного
аналізу (табл. 5.2) нами виявлено 12 статистично значимих
кореляційних залежностей між досліджуваними показниками. Як
й інші автори (Л.С. Мильман, Ю.Г. Юровицкий, 1973; Д.І. Са-
нагурський, 1983; Е.А. Гойда, 1993) ми виявили позитивні
кореляції між АП–ТМП, ШГ–ШПК, ТМП–ШПК, а, крім цього, між
біоелектричними параметрами з ШГ і АК, АК з ШПК та негативні
достовірні кореляції між ТМП і АК з КЗ. Ці результати свідчать
про те, що гіперполяризація зародкової мембрани супровод-
жується підсиленням інтенсивності процесів енергетичного
метаболізму, а також підтверджують відомі експериментальні дані
про те, що джерелом енергії у певний період розвитку зародків
тварин є зазвичай глікоген (Л.С. Мильман, Ю.Г. Юровицкий, 1973;
Ю.Г. Юровицкий и др., 1996; Ю.Г. Юровицкий, 1999). Цілком
зрозумілими є кореляційні зв’язки між АК з іншими показниками
(рис. 5.1), оскільки цей фермент відіграє головну роль у
регулюванні метаболізму і, зокрема, значною мірою її активністю
186
регулюється обмін гліколізу, процеси транскрипції і трансляції
(Л.В. Рябова, С.Г. Васецкий, 1996, 2000; Е.А. Земсков и др., 1998).
Щодо механізму змін середнього рівня ТМП, то
гіперполяризація плазматичної мембрани бластомерів може бути
пов’язана зі змінами розміру зародкових клітин, об’єм яких з
кожним поділом зменшується. Можливість цього механізму
підтверджується у роботі Веренінова зі співавт. (А.А. Веренинов,
1986), де промодельовано зв’язок між змінами ТМП та вмістом у
клітині води (клітинним об’ємом).
Разом з тим, у десяти прикладах може бути ймовірність
причинно-наслідкових зв’язків (рис. 5.1) між змінами певних
показників. Усім виявленим часовим співвідношенням характерні
довготривалі зсуви 30–60 хв (оскільки інтервал квантування
становив 30 хв). Звідси зрозуміло, що зміни досліджуваних по-
казників відображають події, які відбуваються на рівні макроор-
ганізму, та часові взаємозв’язки між ними. Таке припущення
узгоджується з результатами експериментальних досліджень.
Доведено, що починаючи зі стадії 32-х бластомерів,
відбувається активне перенесення з жовтка у бластодерму гліко-
ген-фосфорилазного комплексу (Л.С. Мильман, Ю.Г. Юровицкий,
1973; Ю.Г. Юровицкий, 1999), яке супроводжується збільшенням
швидкості гліколізу. У цей же час у зародків в’юна між
бластодиском та жовтком відбувається формування перибласту, за
рахунок якого здійснюється активне засвоєння компонентів
жовтка (E.I. Furshpan, D.D. Potter, 1968). Крім того, зміни
інтенсивності дихання у цей час можуть бути опосередковані
змінами в ультраструктурі популяції мітохондрій, оскільки
187
встановлено, що зі стадії 16-ти бластомерів змінюється
співвідношення між паличкоподібними та кулястими мітохонд-
ріями, зокрема, збільшується кількість перших (М.Ю. Сухомли-
нова и др., 2000). Ці дані дають змогу пояснити також отримані
нами результати сильної достовірної кореляції між змінами ТМП
та ШПК, яку спостерігають під час довготривалих зсувів, у період
розвитку 16–32 бластомерів, що узгоджується також з даними
крос-кореляційного аналізу коливних складових цих показників.
Отже, виявлені часові співвідношення між динамікою
тренду біоелектричних і метаболічних показників, імовірно,
пов’язані з морфофізіологічними процесами, що супроводжують
розвиток зародків в’юна.
Аналогічне пояснення мають деякі отримані результати
крос-кореляційного аналізу з п’ятихвилинним інтервалом
квантування значень характеристик, якими описують зміни
середнього рівня біоелектричних, метаболічних та регуляторних
факторів у ранньому ембріогенезі в’юна. У результаті цього аналізу
виявлено ймовірність наявності 32-х достовірних причинно-
наслідкових зв’язків, які описано у підсумкових таблицях 5.1–
5.6, де вказано максимальні достовірні значення коефіцієнтів крос-
кореляції (R
xy
) і часові зсуви (t), при яких їх спостерігають. Індекси
R
xy
: x – показник першого стовпчика, y – показник першого рядка;
** – p < 0.01, * – p < 0.05.
Як і в попередньому аналізі виявлено достовірні крос-
кореляційні зв’язки, що простежуються при довготривалих зсувах
між змінами рівнів: pH
ц
з усіма іншими показниками (крім зв’язку
з серотоніном), [Ca
2+
]
в
з активністю ЛДГ і ШПК, а також між

188
активністю ізЦДГ і ШПК. Аналіз зсувів засвідчує, що ці зв’язки,
найімовірніше, пов’язані з процесом запліднення (В.П. Божкова,
1986).
Таблиця 5.1
Крос-кореляційні зв’язки між біоелектричними
та метаболічними характеристиками зародків в’юна
Показ-
ники
кореля-
ції, R
xy
ТМП
Ca
2+
pH
ц
Із
ЦДГ
α-
КГДГ
СДГ
ЛДГ
ШПК
5-HT
ТМП – 0,84** 0,76** – – -0,87** – 0,74** -0,55**
t, хв – 5 25 – – 5 – 80 5
pH
ц
– -0,54* – – – – -0,53** 0,66** –
t, хв – 25 – – – – 40 25 –
Із ЦДГ -0,80** -0,86** -0,72** – – 0,71** – -0,50** –
t, хв 5 5 45 – – 5 – 80 –
α-КГДГ
– -0,70** -0,40* – – – – – –
t, хв – 10–30 55 – – – – – –
СДГ – -0,86** -0,60** – – – – -0,79** –
t, хв – 5 25 – – – – 5 –
ЛДГ – -0,50* – 0,50* – 0,39** – -0,42** –
t, хв – 30 – 25 – 10 – 10 –
ШПК – -0,58** – – -0,74** – – – 0,80**
t, хв – 40 – – 5 – – – 20
5-HT – -0,44* 0,52* 0,46* 0,54* 0,50* 0,51* – –
t, хв – 5 5 10 5 5 5 – –
Відомо, що під час запліднення відбувається зростання pH
ц
на 0,2–0,3 од., яке можливо регулює біосинтетичні процеси,
транспортування речовин через мембрани (В.П. Божкова, 1986),
збільшення мембранної провідності для йонів K
+
(S.S. Shen,
R.A. Steinhardt, 1978). Водночас короткочасно збільшується [Ca
2+
]
в
(за рахунок активування інозитолтрифосфатного шляху), що бере
участь у підсиленні поглинання кисню, пов’язаного з окисненням
ненасичених жирних кислот (D.B. Epel, 1980).
189
Отже, ці дані дають змогу пояснити виявлені часові
співвідношення між певними показниками, але разом з тим не
можна стверджувати про те, що зміни одних показників є
безпосередньою причиною змін інших.
На підставі аналізу кореляційних залежностей, причину
слід шукати у спільних факторах, які однаково впливають на зміни
тих чи інших процесів. Як свідчать факти, які наведено у огляді
Божкової (В.П. Божкова, 1986), можлива незалежність змін одних
показників від інших. Наприклад, збільшення [Ca
2+
]
в
та pH
ц
є
наслідком метаболізму фосфогліцеридів, що відбувається двома
різними шляхами: через диацилгліцерид та інозитолтрифосфат
(M.J. Berridge, 1984).
Діацилгліцерид функціонує в мембрані, активуючи про-
теїнкіназу С та Na
+
/H
+
-обмін, а інозитолтрифосфат дифундує з
мембрани в цитоплазму та сприяє вивільненню Ca
2+
з внутрішньо-
клітинних депо.
Доведено, що активність протеїнкінази С веде до залу-
ження цитоплазми у яйцеклітинах морського їжака не впливаючи
на процеси, що відбуваються за рахунок збільшення [Ca
2+
]
в
(K. Swann, M. Whitaker, 1985).
Оскільки йони Ca
2+
є найбільш універсальним вторинним
посередником, виявлені крос-кореляційні зв’язки за п’ятихви-
линним зсувом між ТМП–Ca
2+
, Ca
2+
–ізЦДГ, СДГ реалізуються на
рівні молекулярних механізмів.
Зокрема, встановлено, що зміни цитозольної концентрації
Ca
2+
мають вплив на мітохондріальну фізіологію, оскільки
регулюють окиснювальний метаболізм впливаючи на окремі
190
матриці дегідрогеназ (C.V. Ammini, W.W. Haus wirth, 1999), а
також на інтенсивність перикисного окиснення ліпідів (И.О. Му-
калов и др., 1984), регулювання мембранної провідності та ін.
(П.Г. Костюк, 1978; С.Э. Шноль, 1979; П.Г. Богач і ін., 1984;
М.Д. Курський, С.М. Кучеренко, 1993; П.Г. Костюк /за ред. і ін., 2001).
Виявлені крос-кореляційні зв’язки між динамікою
концентрації серотоніну з іншими досліджуваними показниками, в
яких динаміка серотоніну проходить з деяким упередженням на 5–
10 хв до змін інших показників (табл. 5.1.). На сьогодні відомо, що
донервовий 5-HT (5-гідрокситриптамін) може брати участь у
сегрегації цитоплазми та клітинних поділах. Внутрішньо-
клітинними ефекторами в реалізації обох функцій служать
елементи цитоскелета (Г.А. Бузников, 1987). Одержано також дані
про можливість функціонального взаємозв’язку серотоніну та
нейропептидів у регуляторних процесах ембріогенезу (А. Виллоуз
и др., 2000). Звідси зрозумілою є кореляція між серотоніном і Ca
2+
,
оскільки йони кальцію беруть участь у полімеризації та скороченні
актинових мікрофіламентів, які входять до складу примемб-
ранного кортикального шару цитоплазми (В.П. Божкова, 1986).
Цей зв’язок може бути опосередкований через аденілатциклазну
систему за участю вторинних месенджерів – іонізованого кальцію
або фосфатидилінозитидів. Підтвердженням цьому є виявлені у
мембранах ооцитів морських їжаків функціонально активні по-
верхневі рецептори до біогенних моноамінів (Н.Е. Ламаш и
др., 1994).
Із досліджень диференційованих клітин відомо, що дія
серотоніну реалізується через вплив на аденілатциклазну систему,