Townsend C.R., Begon M., Harper J.L. Essentials of Ecology
Подождите немного. Документ загружается.


Chapter 3 Physical conditions and the availability of resources
109
REVIEW QUESTIONS
Review questions
Asterisks indicate challenge questions
1* Explain, referring to a variety of specific
organisms, how the amount of water in
different organisms’ habitats may define
either the conditions for those organisms, or
their resource level, or both.
2 Discuss whether you think the following
statement is correct: ‘A layperson might
describe Antarctica as an extreme environment,
but an ecologist should never do so’.
3 In what ways do ectotherms and endotherms
differ, and in what ways are they similar?
4* Drawing examples from a variety of both
animals and plants, contrast the responses of
tolerators and avoiders to seasonal variations in
environmental conditions and resources.
5 Describe how plants’ requirements to increase
the rate of photosynthesis and to decrease the
rate of water loss interact. Describe, too, the
strategies used by different types of plants to
balance these requirements.
6* Describe and account for the differences in
both root and shoot architecture exhibited by
different plants.
7 Account for the fact that the tissues of plants
and animals have such contrasting C : N ratios.
What are the consequences of these
differences?
8 Describe the various ways in which animals use
color to defend themselves against attacks by
predators.
9 Explain, with examples, what exploitation and
interference intraspecific competition have in
common and how they differ.
10 What is meant when an ecological niche is
described as an n-dimensional hypervolume?
9781405156585_4_003.qxd 11/5/07 14:44 Page 109

110
Chapter 4
Conditions, resources and
the world’s communities
CHAPTER CONTENTS
4.1 Introduction
4.2 Geographic patterns at large and small scales
4.3 Temporal patterns in conditions and resources
4.4 Terrestrial biomes
4.5 Aquatic environments
Chapter contents
KEY CONCEPTS
In this chapter you will:
l
understand that conditions and resources interact to help determine
the composition of whole communities
l
appreciate that climatic patterns over the surface of the Earth are
responsible for the large-scale pattern of distribution of terrestrial
biomes (such as tropical rain forest, desert and tundra)
l
recognize that biomes are not homogeneous because local topography,
geology and soil influence the communities of plants and animals
that occur
l
appreciate that conditions and resources at a location may change
over time scales ranging from hours to millennia, leading to parallel
temporal patterns in the composition of communities
l
understand that in most aquatic environments it is difficult to
recognize anything comparable to terrestrial biomes: communities
tend to reflect local conditions and resources rather than global
patterns in climate
Key concepts
9781405156585_4_004.qxd 11/5/07 14:47 Page 110

4.1 Introduction
Having examined in Chapter 3 the way individual organisms are affected by con-
ditions and resources, we now turn to the larger question of how the interplay
of conditions and resources influences whole communities (the assemblages of
species that occur together). The answer to this question depends fundamentally
on the scale at which we choose to study communities; this will be a pervasive
theme throughout the chapter.
Not surprisingly, because of its influence on both conditions and resources,
climate plays a major role in determining the large-scale distribution of different
types of community across the face of the Earth. However, local factors, such as
soil type in terrestrial environments and water chemistry in aquatic environments,
are responsible for patchiness in community composition on much smaller scales.
We discuss some of the causes of spatial patterns in community distribution in
Section 4.2. Then, in Section 4.3, we turn to temporal patterns in conditions and
resources that can change community composition over time scales from days to
millennia. Section 4.4 describes the characteristics of the Earth’s major terrestrial
biomes and Section 4.5 deals with the diversity of aquatic communities.
4.2 Geographic patterns at large and
small scales
4.2.1 Large-scale climatic patterns
At the largest scale, the geography of life on Earth is mainly a consequence of the
planet’s movement through space. The tilt of the Earth as it orbits the sun causes
solar radiation to strike the Earth’s surface with different intensities at different
latitudes (Figure 4.1). Because the equator is tilted toward the sun, equatorial and
tropical areas receive more direct sunlight and are warmer than other latitudes.
Warm air holds more moisture than cold air, increasing the water-holding capa-
city of air around the tropics. Solar radiation draws water from the vegetation
by evaporation, but because the air is so moist, much of the water condenses and
Chapter 4 Conditions, resources and the world’s communities
111
The interplay of conditions and resources profoundly influences the composition
of the world’s communities. At the global scale, patterns of climate circulation are
largely responsible for distinctive terrestrial biomes, such as deserts and rain
forests, with their characteristic assemblages of plants and animals. Distinct types
of marine and freshwater communities can sometimes also be identified at a broad
geographic scale. Within each biome or aquatic category, however, there are
enormous variations in conditions and resources that are reflected in
community patterns viewed at a smaller scale.
scale and patchiness – central
themes of this chapter
solar radiation,...
9781405156585_4_004.qxd 11/5/07 14:47 Page 111

falls back as rain. Thus, the air that cycles to the atmosphere from the tropics is
relatively dry, having lost most of its moisture as local rainfall before it ascends
to the lower atmosphere.
The rotation of the Earth causes air masses from the tropics to curve to the north
and south. Air that was warmed at the tropics (and which lost moisture as local
rain) cools in the atmosphere and descends again at latitudes of approximately 30°
(north and south). The air mass warms as it descends, increasing its capacity to hold
water and causing the descending air mass to ‘soak up’ available water from the land.
As a result, this is where most of the major deserts, including the Sahara, Kalahari,
Mojave and Sonoran, are found. Another smaller evaporation/precipitation system
occurs between 30° and 60° latitude, as warm air, now moist, rises and is blown
further north or south, respectively. As it cools, the air descends again and rains,
producing wetter environments.
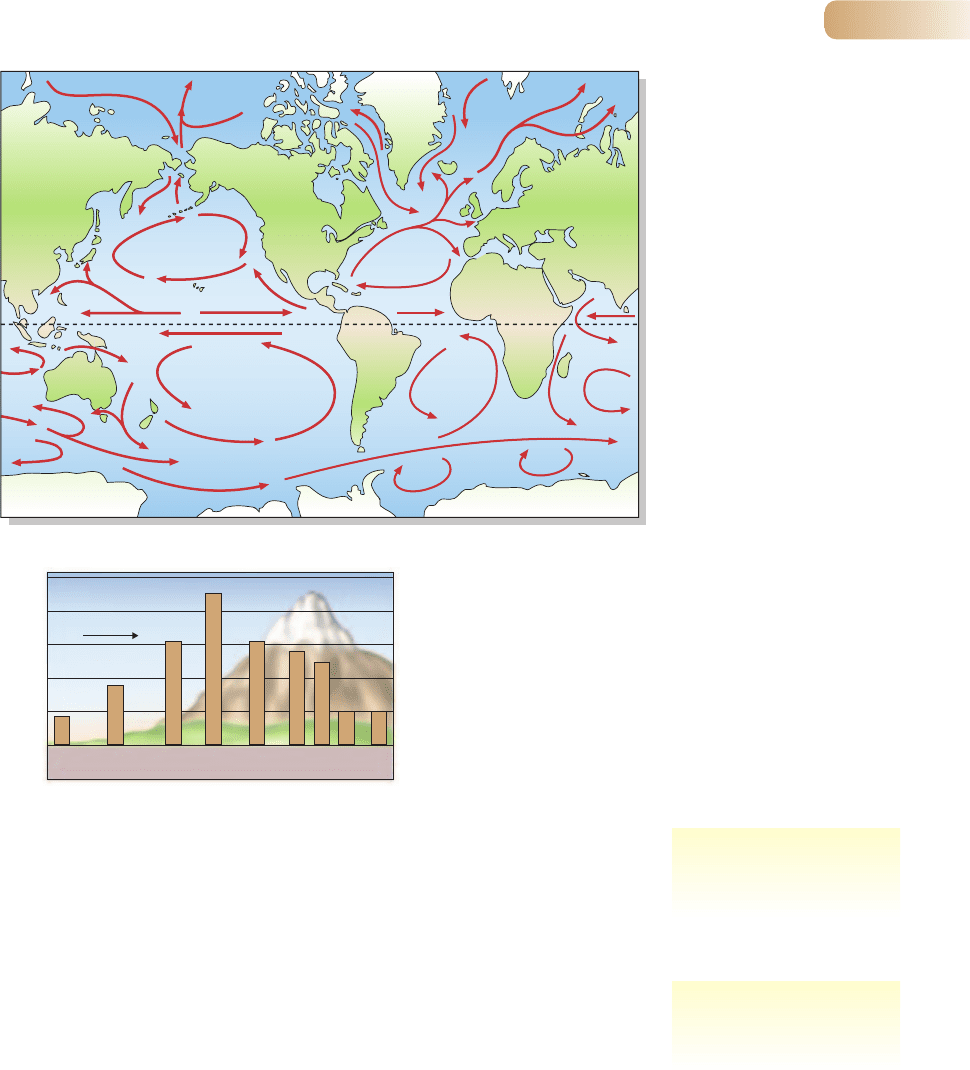
Ocean currents have further powerful effects on climatic patterns. Southern
waters circulate counterclockwise; they carry cold Antarctic waters up along the
western coasts of continents and distribute warmer waters from the tropics along
their eastern coasts (Figure 4.2). In the northern hemisphere, currents circulate
clockwise, carrying cold Arctic waters along the eastern coasts of continents and
returning warm tropical currents along western coasts. The cool, dry climate of
eastern South America is an effect of the Antarctic Humboldt current; the relatively
dry climate of California is a result of Arctic currents. Conversely, on the eastern side
of North America the strong tropical Gulf Stream carries with it warm and moist
air far into the Atlantic Ocean, affecting even the climate of Western Europe.
The topography of the land has consequences at an intermediate scale for the
pattern of terrestrial climates. As winds meet mountain ranges they are forced up
and become cooler as they rise. The cooler air holds less moisture so that water
is released (as rain and snow) on the windward slopes of the mountains (the
Rockies and Himalayas provide striking examples of this effect). As the air passes
over to the leeward sides of the mountains it descends, becomes warmer and now
absorbs water. This produces a desiccating effect and causes a rain shadow along
the leeward slopes (Figure 4.3).
Part II Conditions and Resources
112
E
q
u
a
t
o
r
Sunlight
North Pole
South Pole
Sunlight
Figure 4.1
The tilt of the Earth on its axis and its rotation around the sun define
the amount of radiation striking the atmosphere around the Earth’s
surface. This, in combination with the daily spin of the Earth on its
axis, is responsible for the large-scale patterns of rainfall and solar
radiation that define the pattern of global climate. This diagram
shows winter in the northern hemisphere with radiation falling
almost vertically south of the equator, but the same amount of
radiation is spread over greater areas north of the equator; less
is therefore received, and there is less heating per unit area.
AFTER AUDESIRK & AUDESIRK, 1996
. . . ocean currents...
. . . and mountain ranges...
9781405156585_4_004.qxd 11/5/07 14:47 Page 112
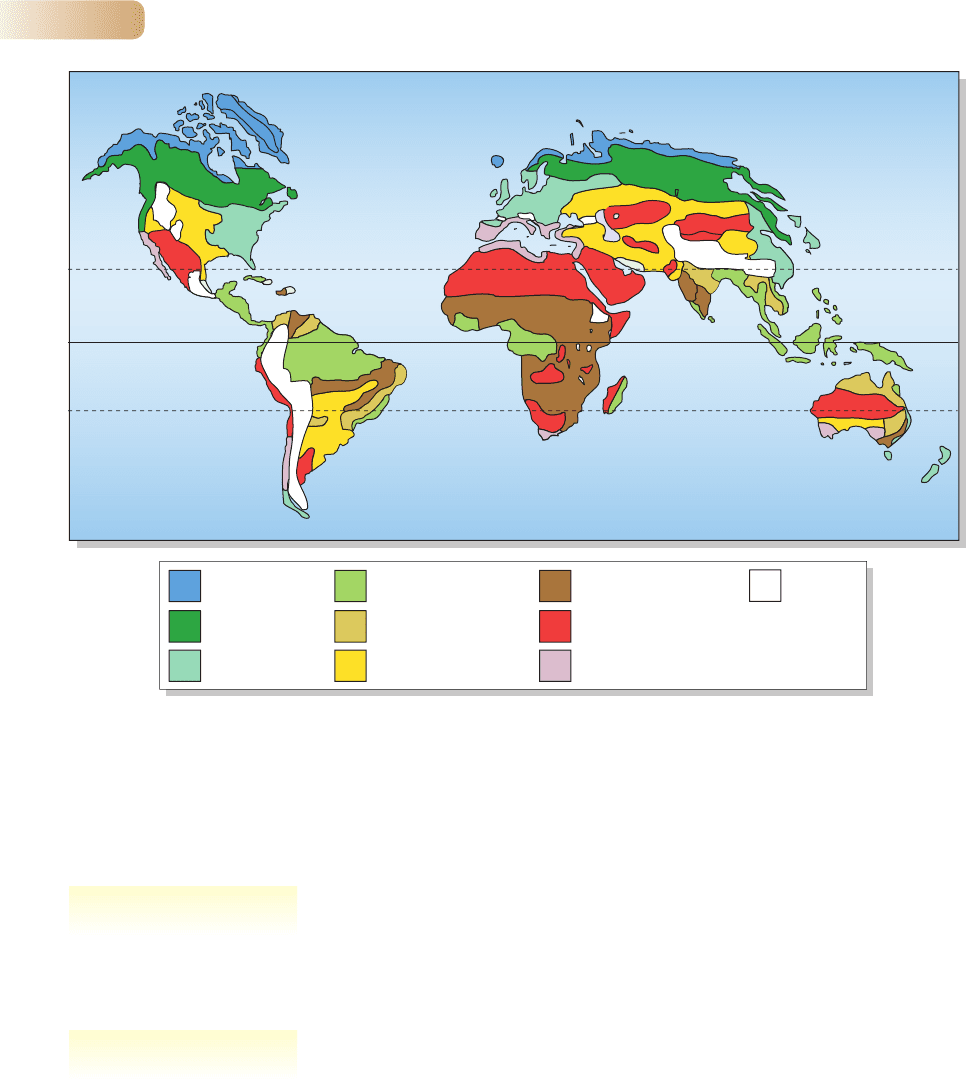
The variety of influences on climate produces a mosaic of dry, wet, cool and
warm climates over the surface of the globe. In the patches of this mosaic, dis-
tinctive terrestrial associations of vegetation and animals have formed. A world
traveler sees repeatedly what can be recognized as characteristic types of
vegetation, which ecologists call biomes (such as desert, savanna and rain forest).
Figure 4.4 recognizes a set of biomes and shows their distribution as a global
map. Figure 4.5 shows the ranges of rainfall and mean monthly minimum
temperature that are critical in determining where the biomes are found. The
characteristics of the communities inhabiting major biomes are described in
Section 4.4.
4.2.2 Small-scale patterns in conditions and resources
It is easy to be seduced by cartographers who draw sharp lines on maps to
show geographic boundaries. But neat pigeonholes, sharp categories and tidy
boundaries are a convenience, not a reality of nature. Moreover, biomes are not
homogeneous within their hypothetical boundaries; every biome has gradients
Chapter 4 Conditions, resources and the world’s communities
113
Average annual precipitation (cm)
100
50
0
West East
Winds
5000
4000
3000
2000
1000
0
Altitude (m)
Figure 4.2
The movements of the major ocean
currents. The general circulation
in the northern hemisphere is
clockwise, in the southern
hemisphere counterclockwise,
with consequences for continental
climate patterns.
Figure 4.3
The typical influence of topography on rainfall (histogram bars) in
the northern hemisphere. Moisture-laden westerlies are forced
higher by a mountain range. As they rise they become cooler and
release the moisture as rain or snow. This leaves a drier rain
shadow on the eastern slopes.
AFTER AUDESIRK & AUDESIRK, 1996
. . . produce a mosaic of dry,
wet, cool and warm climates
over the face of the Earth...
. . . that, in turn, are responsible
for the large-scale distribution of
terrestrial biomes
9781405156585_4_004.qxd 11/5/07 14:47 Page 113
of physicochemical conditions related to local topography and geology. The com-
munities of plants and animals that occur in different parts of this heterogeneous
patchwork may be quite distinct.
Local variations in topography can override the broad climatic pattern described
in Section 4.2.1. For example, temperature falls with increasing altitude and
one effect is that vegetation high on a mountain in the tropics tends to resemble
vegetation at low altitudes in northern latitudes. Traveling up a mountain in the
tropics involves passing along a similar ecological gradient to that experienced
when traveling northward from equator to pole (Figure 4.6).
It is worth remembering that the Earth’s surface would consist of a mosaic
of different environments even if climate were identical everywhere. Geological
history has provided a variety of rocks that differ in their mineral composition.
When the surfaces of these rocks are decomposed by heat, frost and thaw, they
give rise to a variety of types of soil that reflect their geological origin. Without
soil, it is impossible for significant terrestrial vegetation to grow. Soils provide
a source of stored water, a reserve of mineral nutrients, a medium in which
atmospheric nitrogen can be fixed for plant use, and the support that allows plants
to stand up and expose their leaves to the sunlight.
Part II Conditions and Resources
114
Northern
coniferous forest
Arctic tundra
Desert
Mountains
Temperate forest
Tropical rain forest
Tropical seasonal forest
Temperate grassland
Tropical savanna,
grassland and scrub
Mediterranean vegetation,
chaparral
Figure 4.4
World distribution of the Earth’s biomes. Their characteristic plant and animal communities are described in Section 4.4.
local topography
local geology and soil
9781405156585_4_004.qxd 11/5/07 14:47 Page 114

Limestone rocks and chalk originated as marine deposits of calcium carbonate,
often containing some magnesium and other carbonates. Where these deposits
have been raised and exposed as land surfaces they become the basis for neutral
or slightly alkaline calcareous soils, which bear a characteristic calcium-loving
flora. On the other hand, plants normally found on more acid soils, such as
Rhododendron and Azalea, are unsuccessful on calcareous soils. Strict calcium-lovers,
in contrast, suffer on acidic soils, where they are intolerant of aluminum ions
released at low pH. In the United States, for example, the calcium-loving yellow
poplar (Liriodendron tulipifera) and northern white cedar (Thuja occidentalis)
are found only on neutral or alkaline soils, whilst balsam fir (Abies balsamea) and
eastern hemlock (Tsuga canadensis) are usually confined to highly acidic soils.
Variability in the organic matter component of soil also influences the biota
that can occur. Organic matter accumulates at different rates in different soils and
local variations in the balance between mineral and organic material in the soil
contribute to the complexity of environmental mosaics. In extreme conditions,
especially where the rocks are acidic, the temperatures are low and/or the soil is
waterlogged, the decomposition of organic matter may be seriously impeded.
Chapter 4 Conditions, resources and the world’s communities
115
Minimum tem
p
eratur
e
(monthly avera
g
e,°C
)
40
–
60
0
5000
M
inimum tem
p
eratur
e
(monthly avera
g
e,°C
)
40
–
60
0
5000
4
0
–
60
0
5000
Minimum tem
p
eratur
e
(
monthly avera
g
e,°C
)
40
–
60
0
5000
40
–
60
0
5000
(
a
)
Tro
p
ical rain fores
t
(b) Savann
a
(c) Temperate deciduous forest
(
d
)
Northern coniferous forest
(
e
)
Tundr
a
Manaus (South America)
alia)
T
otal annual rainfall (mm
)
Figure 4.5
The variety of environmental
conditions experienced in terrestrial
environments can be described in
terms of annual rainfall and mean
monthly minimum temperatures.
The diagrams show the range
of conditions experienced in
(a) tropical rain forest, (b) savanna,
(c) temperate deciduous forest,
(d) northern coniferous forest (taiga),
and (e) tundra. Data points for a
given biome come from different
locations around the world.
To illustrate this, data points for
tropical rain forest on three different
continents are shown in (a). Tropical
rain forest has characteristically
high mean monthly minimum
temperatures and high rainfall.
In contrast, tundra has both low
temperatures and low precipitation.
The other biomes occupy
intermediate positions in this
two-dimensional representation.
acidic and calcareous soils bear
very different vegetation
9781405156585_4_004.qxd 11/5/07 14:47 Page 115

Then, peat bogs, with their very specialized plants and animals, form on the
partially decomposed organic matter.
To an ecologist, a patch in a community is an area in which a single variable
distinguishes it from its surroundings. Thus, a fallen tree in a forest leaves a gap in
the canopy and a patch on the forest floor where sufficient radiation may penetrate
to allow seedlings to grow and eventually fill the gap. A tide pool is a patch on a
rocky shore, but within that pool snails may graze and clear a patch free of algae.
It is often useful to think of patches as the scale at which particular organisms
experience the environment around them. For an aphid in a forest, an individual
leaf of a particular species of tree is a patch – it provides both the conditions and
the resources necessary for the insect. For a warbler feeding on caterpillars, the
canopies of individual trees are patches that it encounters in its daily life. But owls
or hawks hunt over a large part of the forest, and for them a patch may be the
territory that each bird defends or perhaps even the whole forest over which it ranges.
4.2.3 Patterns in conditions and resources in
aquatic environments
In most aquatic environments it is difficult to recognize anything comparable to
terrestrial biomes. The exceptions occur at the ocean’s edge; tropical mangrove
swamps, coral reefs and temperate kelp forests have biotas that are as distinctive as
Part II Conditions and Resources
116
High
Low
Altitude
Equatorial regions Polar regionsLatitude
Tundra
Tropical forest
Rock, snow, and ice
Coniferous forest
Deciduous forest
Figure 4.6
The effect of altitude and latitude on the distribution of biomes. Moving up in altitude is very similar to moving from equator to pole.
patchiness is in the eye of
the observer...
. . . and all communities
are patchy
9781405156585_4_004.qxd 11/5/07 14:47 Page 116

any of the various terrestrial biomes, but this is largely due to their close relation-
ship with major terrestrial climates. In contrast, the open oceans form a continuum
in which there is flow of water and dissolved chemicals across the globe. We have
seen how variation in the intensity of solar radiation from place to place and
between the seasons has dramatic effects on the temperature and water relations of
terrestrial environments. But this is not the case in the oceans. The high thermal
capacity of water makes the oceans slow to heat and slow to cool. One effect is that
the temperature of the water at one point on the globe is a better reflection of where
the water has come from (along ocean currents) than of the local climate.
The world’s large lakes can be distinguished and classified according to their
physical conditions. For example, large lakes in lowland equatorial regions gener-
ally experience permanent stratification (distinct layers of water at particular
temperatures), whereas seasonal patterns of stratification (in summer) and mixing
(in fall) are the rule in temperate regions. Within the polar circles, permanent ice
cover with no mixing is characteristic of large lakes. However, local geological
conditions and basin size and shape have strong influences on conditions and
resources in lakes, particularly in terms of water chemistry, a key determinant of
lake flora and fauna. Consequently, a broad geographic classification of lake com-
munities has only limited merit. In the case also of streams, rivers, estuaries and
the open ocean, we will see that local conditions and resources are paramount in
determining community patterns (see Section 4.5).
4.3 Temporal patterns in conditions
and resources
The composition of communities can change over time scales ranging from hours
to millennia, as conditions and resources themselves change. For example, the
microbial community that colonizes and decomposes a dead mouse or fragment
of a leaf may change from hour to hour. At the other extreme, we can trace
patterns in community composition over tens of thousands of years. Thus, changes
in climate during the Pleistocene ice ages bear much of the responsibility for
present patterns of distribution of plants and animals. In the 20,000 years since
the peak of the last glaciation, global temperatures have risen by about 8°C. Many
tree species continue, even today, to migrate northward, following the retreat of
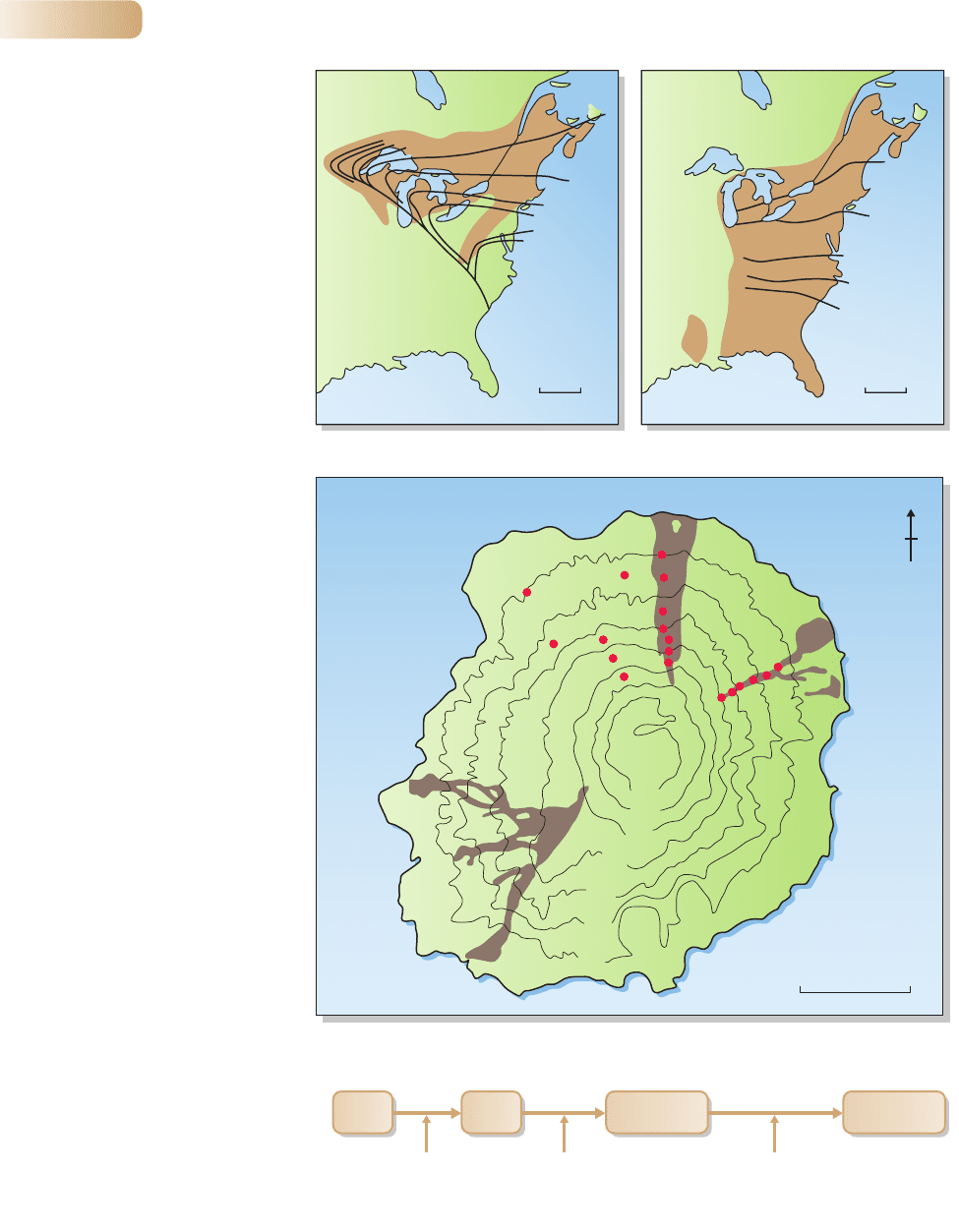
the glaciers (Figure 4.7).
At intermediate temporal scales, predictable sequences of plant species may
occur over periods ranging from years to centuries. For example, the successional
sequence that occurs on cooled volcanic lava takes several centuries to run its
course. This has been documented by comparing the plants living on lava flows
from eruptions that occurred at different times on Miyake-jima Island, Japan
(Figure 4.8). In the earliest stage of succession, conditions are harsh and soil
is sparse and lacking in nitrogen-containing ions, an essential plant resource.
Alders are first to colonize because they can fix atmospheric nitrogen into usable
form. As nitrogen availability in the soil increases, many species of fern, herb,
liana and tree enter the succession. After a century or two, late-successional trees
(Machilus then Castanopsis) shade out many of the earlier arrivals. Succession – the
predictable sequence of colonization and extinction after a disturbance – depends
partly on changing conditions and resources, and partly on the differential com-
petitive abilities of the plants themselves, a topic we return to in Chapter 9.
Chapter 4 Conditions, resources and the world’s communities
117
plant succession – the species
sequence on volcanic lava flows
9781405156585_4_004.qxd 11/5/07 14:47 Page 117
Part II Conditions and Resources
118
0 400
km
0 400
km
8,000
8,000
9,000
9,000
10,000
10,000
11,000
12,000
13,000
1,000
2,000
7,000
7,000
6,000
(a) (b)
Figure 4.7
A map showing the spread of
two species of forest tree in eastern
North America after the retreat of
the last ice age glaciation. Note that
the two species of (a) eastern white
pine (Pinus strobes) and (b) beech
(Fagus grandifolia) have not followed
the same invasion path. The lines on
the maps (isochrones) define the
time of arrival of each species at
1000-year intervals. The numbers
on the map refer to thousands of
years before present. The shaded
brown areas show their present
distributions.
Figure 4.8
(a) Locations of sampling sites (red
dots) on 37 and 125-year-old lava
flows on Miyake-jima Island, Japan.
Sampling on 16-year-old lava was
non-quantitative (no sampling sites
shown). Sites outside these flows
are at least 800 years old. Altitudinal
contours are shown in meters.
(b) In the earliest stage of
succession the only vegetation
consists of a few small alder trees
(Alnus sieboldiana). In the older
plots (37–800 years old),
113 species were recorded,
including ferns, herbs, lianas and
trees. This succession consisted of:
(i) colonization of the bare
lava by the nitrogen-fixing alder;
(ii) facilitation (through improved
nitrogen availability) of mid-
successional Prunus speciosa and
the late-successional evergreen
tree Machilus thunbergii;
(iii) establishment of a mixed forest
in which Alnus and Prunus were
shaded out; and (iv) competitive
replacement of Machilus by the
longer lived Castanopsis sieboldii.
FROM DAVIS, 1976AFTER KAMIJO ET AL., 2002
(b)
(a)
02 km
700
600
500
400
300
200
100
125-year-old lava flow
37-year-old
lava flow
16-year-old
lava flow
Colonization
of Alnus
Facilitation by N
fixation of Alnus
Colonization of Prunus
and Machilus
Disappearance of
Alnus and Prunus
Colonization of Castanopsis
Castanopsis
forest
Machilus and
Prunus forest
Alnus
shrub
Bare
land
0-
year-old
16-
year-old
37-
year-old
125-
year-old
800-
year-old
N
9781405156585_4_004.qxd 11/5/07 14:47 Page 118