Волькенштейн М.В. Общая биофизика
Подождите немного. Документ загружается.

§
4.3.
ГЕНЕРАЦИЯ
ИМПУЛЬСА
191
он
не станет меньше срг, т. е.
|-^)-
l]exp[—^
2,
(4.43)
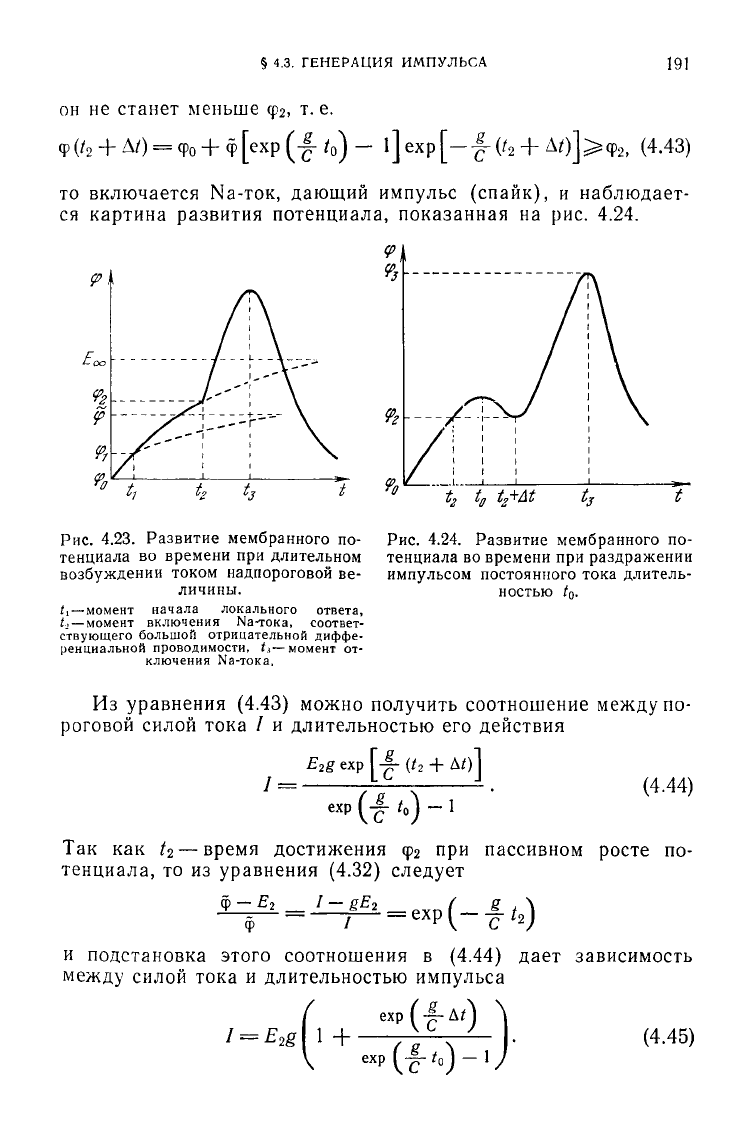
то включается Na-ток, дающий импульс (спайк), и наблюдает-
ся
картина развития потенциала, показанная на рис. 4.24.
9>
Рис.
4.23. Развитие мембранного по-
тенциала во времени при длительном
возбуждении током надпороговой ве-
личины.
Л
—момент
начала локального ответа,
U—
момент
включения
Na-тока,
соответ-
ствующего
большой отрицательной
диффе-
ренциальной
проводимости, (.i—момент от-
ключения
Na-тока,
Рис.
4.24. Развитие мембранного по-
тенциала во времени при раздражении
импульсом постоянного тока длитель-
ностью to.
Из
уравнения
(4.43)
можно получить соотношение
между
по-
роговой силой тока / и длительностью его действия
E
2
g
ехр [-£- (t
2
+ Л/)]
(4.44)
Так
как ^
2
— время достижения <р
2
при пассивном росте по-
тенциала, то из уравнения
(4.32)
следует
и
подстановка этого соотношения в
(4.44)
дает
зависимость
между
силой тока и длительностью импульса
(4.45)
192
ГЛ.
4.
НЕРВНЫЙ ИМПУЛЬС
При
очень малых /
0
получаем приближенно
/
0 \
Е
2
С ехр I -рг Л/ I
I
(4.46)
где Q — количество электричества, проходящее через мембрану
при
очень коротких пороговых толчках тока. При очень большой
длительности импульса пороговая сила тока равна /
п
= E
2
g.
Эта величина является реобазой (см. стр. 166). Характеристиче-
ское время T
S
определяется соотношением
x
s
~Q/I
a
.
(4.47)
С
учетом
уравнения
(4.46)
получаем
С __._ ( g
4
Л
(448)
(4.49)
Значение
величины ///
п
в точке /
0
= т
« равно
/
/п
=
1
+
exp
[(g/C)
At]
exp
[(g/C)
At] - 1
Для аксона кальмара RC = 0,7 мс. Время Д^ ж
0,3—0,4
мс.
Подстановка этих величин в
(4.49)
дает
1,36—1,42;
экспери-
ментально найденное значе-
///„
| ние равно 1,38 [66].
Соотношение
«сила —
длительность» в единицах
реобазы имеет вид
_/_,
exp
[(g/C)
At]
I
n
~T~
exp
[(g/C)
t
o
]-\ •
(4.50)
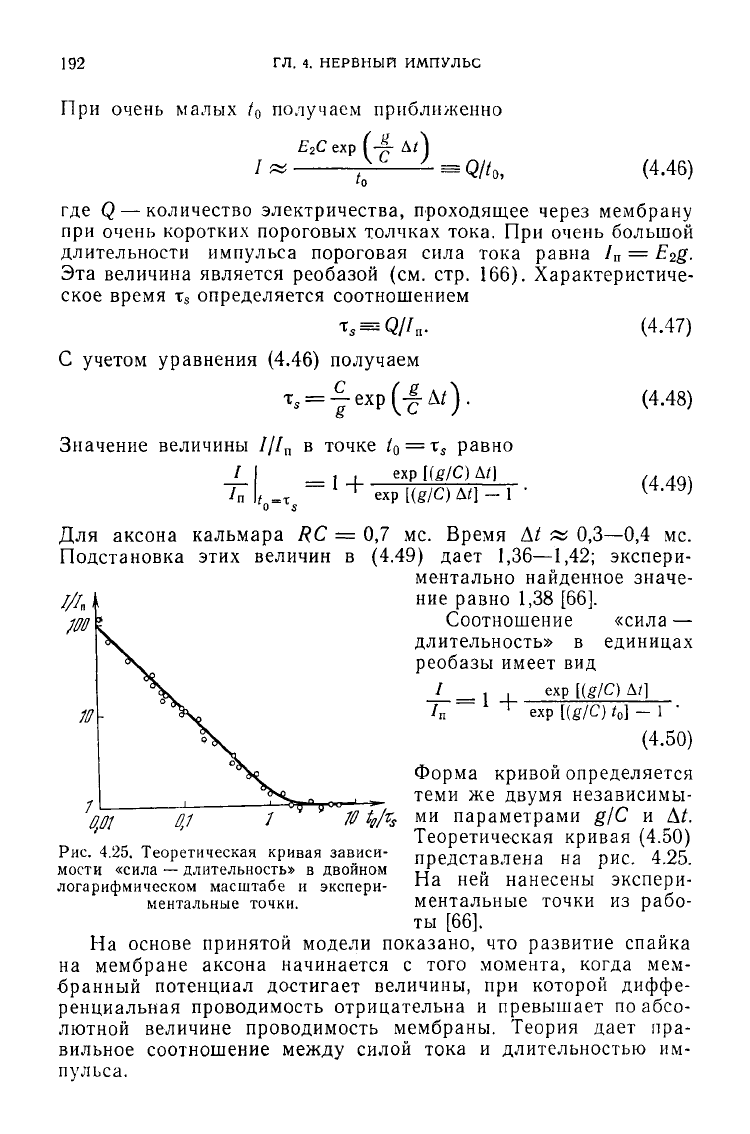
Форма
кривой определяется
теми же двумя независимы-
10
t
e
/r
s
ми параметрами g/C и Д^.
Теоретическая кривая
(4.50)
Рис.
4.25. Теоретическая кривая зависи- представлена на рис. 4.25.
мости «сила — длительность» в двойном
логарифмическом
масштабе и экспери-
ментальные точки.
На
ней нанесены экспери-
ментальные точки из рабо-
ты [66].
На
основе принятой модели показано, что развитие спайка
на
мембране аксона начинается с того момента, когда мем-
бранный
потенциал достигает величины, при которой диффе-
ренциальная
проводимость отрицательна и превышает по абсо-
лютной величине проводимость мембраны. Теория
дает
пра-
вильное соотношение
между
силой тока и длительностью им-
пульса.

§
4.4.
АКТИВАЦИЯ
И
ИНАКТИВАЦИЯ
НАТРИЕВОЙ
ПРОВОДИМОСТИ
193
§
4.4. АКТИВАЦИЯ И ИНАКТИВАЦИЯ НАТРИЕВОЙ ПРОВОДИМОСТИ
Обратимся теперь к модельной теории активации и инакти-
вации
Na-проводимости. Такая теория предложена в работах
[67—70].
В опытах на аксоне кальмара, проведенных методом фикса-
ции
напряжения на мембране, Ходжкин и Хаксли установили
экспоненциальную зависимость
между
величиной начального
изменения
мембранного потенциала ср и пиковым значением
натриевой проводимости gNa [51]. Изменению ср на 4—6 мВ в
сторону деполяризации отвечает е-кратное повышение gNa- Мо-
жно предположить, что изменения ионной проводимости яв-
ляются результатом влияния электрического поля на положе-
ние
каких-то ионов в мембране. Считая, что проводимостьgNa
пропорциональна
доле
этих ионов, смещенных мембранным по-
тенциалом из исходного положения, получаем
а =
gNa
'
max
, (4 5П
«Na i
+
b exp (-
zayTIRT)
V
>
где z— валентность гипотетического иона, а — число, выража-
ющее
долю
напряжения ф, действующего на ион при его пере-
мещении,
Ь — множитель, не зависящий от ср. В области ср <0
получаем асимптотически
In
g
Na
ж const +
za^grjRT.
(4.52)
Из
начального
угла
наклона экспериментальной прямой
In
gNa (ф) получается za « 6 [51]. Параметр za можно опреде-
лить также из центральной части кривой
(4.51)
по ее наклону
В
ТОЧКе
gNa =
0,5gNa,max
[67].
Формула
(4.51)
ХОрОШО
СОГЛЭ-
суется с многочисленными экспериментальными данными, по-
лученными на различных аксонах [71—73]. Параметр za, най-
денный по (4.52), имеет, согласно этим работам, значения от
3,6 до 6, по
(4.51)
—от 2 до 4,4. Среднее значение za равно 3,5.
Как
уже сказано (см. стр. 178), Ходжкин и Хаксли предпо-
лагали, что эти гипотетические ионы определяют активацию и
инактивацию каналов для натрия и калия. Была высказана ги-
потеза о том, что эти ионы — ионы Са**. Действительно, из-
вестно, что изменения концентрации Са
++
сильно влияют на
возбудимость нервных и мышечных клеток [74] (см. гл. 5).
Было
показано, что при пятикратном изменении концентрации
ионов
Са
++
кривые зависимости gNa, gK
и
инактивации от ср
смещаются вдоль оси напряжения на 10—15 мВ. Понижение сс
а
смещает кривые в сторону меньших ср. Концентрация Са
++
в
растворе сильно влияет на скорость уменьшения gNa при сня-
тии деполяризации. В цитируемой выше работе было выдви-
нуто предположение о блокировании каналов в мембране ио-

194 ГЛ. 4.
НЕРВНЫЙ
ИМПУЛЬС
нами
Са
++
. Отношение доли р каналов, закрытых кальцием, к
доле открытых каналов 1 — р
р/(1
— р) со с
Са
ехр (—
2ay&~/RT),
(4.53)
где аф — напряжение, под влиянием которого ионы Са
++
пере-
мещаются от внешнего раствора к месту закрытия канала.
Если
это место расположено в середине мембраны, то а = 0,5,
если у внутреннего края, то а = 1,0. Согласно
(4.53)
е-кратное
изменение
сс
а
эквивалентно изменению <р не менее, чем на
12,5 мВ (если ос= 1,0). Между тем опыт
дает
от 4,5 до 9 мВ
[71—73, 75].
Вторая трудность «кальциевой гипотезы» связана со значе-
нием
z<x в формуле (4.51). а всегда меньше 1, z для Са
++
рав-
но
2. Между тем из экспериментального значения га « 3,5
следует,
что z J> 4. Предположение о том, что в блокировке
каналов
участвуют
пары ионов Са
++
наталкивается на суще-
ственное противоречие — получается смещение кривой gNa(cp),
значительно превышающее наблюдаемое на опыте [67].
В работах [6, 19] исследовалось возбуждение мембраны
аксона
с помощью метода перфузии. Было установлено, что
разбавление внешнего раствора изотоническим неэлектролитом
повышает потенциал покоя. Аналогичное разбавление внутрен-
него раствора не оказывает влияния. Напротив, замена С1~ во
внешнем
растворе на Br", SO
4
"и т. д. не влияет на потенциал
покоя,
а во внутреннем растворе она влияет на него. Следова-
тельно, на внешней стороне мембраны имеется много
фикси-
рованных отрицательных зарядов, а на внутренней их мало.
Возрастание внешней концентрации двухвалентных катионов
стабилизует состояние покоя, а ее уменьшение приводит к за-
метным флуктуациям мембранного потенциала.
Ионы
Са
4+
ока-
зываются необходимыми для сохранения мембраны в состоя-
нии
покоя. На этих основаниях предполагается, что мембрана
обладает свойствами катионообменника — в состоянии покоя
доступные для обмена отрицательно заряженные центры на
внешней
стороне мембраны заняты двухвалентными ионами
Са
++
.
При стимуляции возникает ионный обмен. Процесс воз-
буждения можно считать подобным фазовому
переходу
макро-
молекул мембраны из одной устойчивой конформации в дру-
гую. Обмен ионов Са
++
на Na
+
развивается кооперативно. Воз-
буждение триггерно стимулируется переносом внутрь ионов Na
+
,
замещающих внешние ионы Са
++
, причем изменяется
физико-
химическая
структура
мембраны.
Проведем, пользуясь методом молекулярного поля, анализ
статистической модели возбудимой мембраны [67]. Предста-
вим
мембрану как совокупность периодически расположенных
ячеек,
каждая из которых может находиться в проводящем или

§
4.4. АКТИВАЦИЯ И ИНАКТИВАЦИЯ НАТРИЕВОЙ ПРОВОДИМОСТИ 195
непроводящем состоянии. Проводимость мембраны пропорцио-
нальна доле ячеек, находящихся в проводящем состоянии. Пред-
положим, что энергия взаимодействия
двух
соседних ячеек рав-
на
ец. если их состояния одинаковы, и ею, если они различны.
Разность е^ец — ею является параметром кооперативности.
Каждая ячейка может присоединить ион валентности z, кото-
рый при перемещении из раствора в мембрану пересекает иоле
с разностью потенциалов аср. Присоединение иона изменяет
энергию ячейки на U
-\~
za.(p!¥~
(в расчете на моль) и
делаег
ячейку непроводящей. В приближении молекулярного поля по-
лучаем
{
'_ exp [ve (2p —
l)/RT]
со с
Са
exp(—
zatp&'/RT),
(4.54)
где
v
—координационное
число.
Это
выражение
совпадает
с
(4.53)
в
отсутствие
кооперативности
(при е—0). Из
(4.54)
следует,
что
га
:0
.5~
2
(2-(5)'
*•—
RT'
а
так как g
Na
со 1 — р, то
я
™
(4.55)
^г«т
•
(4-56)
d (fST/RT)
В отсутствие кооперативности, т. е. при р = 0, эта производная
равняется га/4. Как уже сказано, опыт
дает
га да 3,5. Такой же
наклон
кривой £к
а
(ф) можно получить из формулы
(4.56)
при
любых га < 4, если задать соответствующее значение р > 0.
Таким образом, наклон кривой £^а(ф) можно объяснить при
г = 2. Однако смещение кривой при изменении сс
а
с этим не
согласуется. Из
(4.54)
следует
условие смещения кривой
In
сса — zaytF/RT — const
и
величина изменения ф при е-кратном изменении сса равна
Дф :=
25/га
(в мВ) независимо от значения р. Совпадение этого
значения Дф с опытом получается лишь при га да 3,5.
Отсюда
следует,
что одновременное объяснение
двух,
не-
сомненно связанных
друг
с
другом
эффектов, — формы кривой
£ка(ф) и смещения этой кривой при изменении Сса, нужно
искать на основе
другой
модели. Резкое возрастание проводи-
мости обусловлено перемещением внутри мембраны заряда ве-
личины z ^ 4, которое должно быть связано с десорбцией од-
ного иона Са
++
. Если заряд совершает в поле работу
2ац)&~,
где га — 3,5, то для получения согласия с опытом надо при-
нять
близкое к нулю значение р, т.е.
малую
кооперативность
или ее отсутствие.

196 ГЛ. 4.
НЕРВНЫЙ
ИМПУЛЬС
Модель, объясняющая основные экспериментальные факты,
предложена в работе [68].
Допустим, что в мембране имеется N ячеек, каждая из ко-
торых содержит молекулу или группу молекул (активирующую
частицу, способную находиться в состоянии*) А или В. Пред-
положим, что переходы А
ч=^
В связаны с совершением работы
в электрическом поле. Такие переходы
могут
состоять в пере-
мещении заряженной молекулы, в поворотах диполей или в
конформационном
превращении макромолекулы, приводящем к
одновременному перемещению нескольких зарядов. Последний
вариант подтверждается опытами, свидетельствующими о нали-
чии структурных изменений мембраны во время увеличения
проницаемости [76, 77]. Если z — суммарный заряд, перемещае-
мый при переходе
А—*В,
то работа моля частиц равна гссф#".
Предположим, что в состоянии А частица образует комплексы
А
•
Са с ионами Са
++
. В состоянии В активирующая частица
обладает такой структурой или положением, что способствует
прохождению иона Na
+
через мембрану. Перенос Na
+
лимити-
руется числом частиц в состоянии В. Обозначим числа активи-
рующих молекул в состояниях А и В через х я у, а число комп-
лексов А
•
Са через s. Имеем
£
Na
= by.
Таким образом, исследование временных изменений проводи-
мости при сдвиге ф сводится к изучению зависимости y(t).
Модели отвечает схема реакций
А-Са Дса + А^В.
*_, *_
2
Имеем
K
2
= k-
2
/k
2
=
qexp{-zacpg-/RT),
(4.57)
где q—фактор, не зависящий от поля.
Кинетические уравнения имеют вид
(4.58)
Из
изложенного выше
следует,
что пиковой Na-проводимо-
сти соответствует значение у, близкое к равновесному значе-
нию у
щ
. В стационарных условиях, т. е. при у = s = 0, из
(4.58)
получаем
К
2
+
К^К
2
с
с&
),
(4.Б9)
*) Здесь и далее сами частицы мы также
будем
обозначать через А, В
и
т. д.

§
4.4. АКТИВАЦИЯ И ИНАКТИВАЦИЯ НАТРИЕВОЙ ПРОВОДИМОСТИ 197
где Ki = k-i/ki. Учитывая значение /С
2)
получаем
£N3 = *y
eq
= £
N
a. m.4
1 +
(
l + K
^)
Я
^Р (~ ^f)] . (4.60)
где
gNa,msiX
= kN (k — коэффициент пропорциональности). Как
мы видели, экспериментальная форма кривой gNa(<p) хорошо
описывается формулой такого вида, если га « 3,5. Смещение
кривой
(4.60) вдоль оси ф при изменении сс
а
дается условием
(1
+ /^
Са
) <?ехр (-^f-) = const. (4.61)
l
Если
К\ > ccl, что согласуется с представлением о комплек-
сах А • Са как о резервуаре активных частиц [68], то из (4.61)
следует
In
c
Ca
- ^f- = const (4.62)
и
величина Дер при е-кратном изменении сс
а
равна RTjza&", что
дает 7 мВ при га = 3,5. Это хорошо согласуется с опытом [71,
73,
75]. Такие же смещения кривой ^а(ф) вызывают другие
двухвалентные ионы: Mg
++
, Co
++
, Cd
++
, Ba
++
, Zn
++
и
Ni
++
.
Оче-
видно,
что они
могут
замещать Са
++
в рассматриваемой модели.
Кривая
ё^а(ф) смещается также при изменении рН [63].
При
рН < 5,5 Аф = 13,5 мВ, при рН > 5,5 Дф = 1,3 мВ при
е-кратном изменении концентрации Н
+
. Если ионы Н
+
могут
попарно
связываться активирующими частицами в состоянии А,
то их требуется в два раза больше, чем ионов Са
++
. Условие
смещения
(4.62) приобретает вид
In
(1 + Ki [H
+
]
2
) -
гщ&~1ЯТ
= const (4.63)
и,
если /Ci[H
+
]
2
» 1, то
2 In [H
+
] - za(f^/RT = const. (4.64)
Увеличение [Н
+
] в е раз равносильно уменьшению ф на вели-
чину
Дф
= 2RT/za&- = 2RTfi,bT = 14 мВ,
что совпадает с опытными данными в области рН < 5,5.
При
малых [Н
+
], если /Ci[H
+
]
2
<l, формула (4.63) также
приводит к совпадению с опытом [68].
В нестационарных условиях решение уравнений (4.58)
имеет вид
У (/) =
</о
+ J/i ехр (г
у
() + г/
2
ехр(л
2
/). (4.65)

198 гл. 4.
НЕРВНЫЙ
ИМПУЛЬС
Кривая,
описываемая (4.65), имеет
S-образную
форму
(рис.
4.26),
что согласуется с видом экспериментальной кривой
gNa(0- Константы г/; и г,- зависят от параметров системы &,-,
k-i, N, сса- Подобрав численные значения констант, можно до-
биться полного согласия теоретической кривой
(4.60)
с экспе-
риментальной.
Аналитическое исследование y{t) позволяет
установить зависимость начальной скорости изменения gNa(O)
от сса- Расчет
дает
эту начальную скорость [68]
£
№
(0) = —
k_
i
Nc
Ca
K
2
/(\
+ К
2
).
(4.66)
Опыт подтверждает линейную зависимость скорости падения
gNa от Сса при снятии деполяризации. Действительно, пониже-
ние
Сса в пять раз от 112 до 22 мм умень-
шает скорость изменения g^
a
также в пять
раз,
а от 112 до 4,4 мм — в 25 раз. Из
формулы
(4.66)
следует
также, что, ском-
пенсировав уменьшение сса изменением Ф
так, чтобы произведение /(гСса оставалось
постоянным,
мы должны наблюдать умень-
—^шение
скорости падения g^
a
, потому что/(
2
^
входит
еще и в знаменатель.
Рис.
4.26. график Та
же
модель позволяет рассчитать
функции
у (г). влияние сса на кинетику подъема gNa при
возникновении
деполяризации [68]. В согла-
сии
с опытом теория
дает
замедление скорости роста gNa при
увеличении сс
а
-
Таким
образом, модель объясняет большую совокупность
фактов,
относящихся к Na-проводимости клеточной мембраны.
Можно
думать,
что сходный физико-химический механизм оп-
ределяет и изменения К-проводимости.
Рассмотрим теперь инактивацию Na-проводимости возбуди-
мой
мембраны.
Ходжкин и Хаксли высказали две альтернативных гипотезы
о
природе повышения g^
a
[50]. Первая из них предполагает
химическое превращение активирующих частиц после их пере-
мещения
из положения, которое они занимали при потенциале
покоя.
Вторая предполагает, что уменьшение g^
a
обусловлено
перемещением особых инактивирующих частиц, постепенно
блокирующих каналы проводимости. Вторая гипотеза при-
нята
в теории Ходжкина — Хаксли (см. стр. 179). Модель,
предложенная в работе [67], исходит из первой альтерна-
тивы.
При
потенциале покоя активирующие частицы находятся
преимущественно в состоянии S, когда они связаны с Са
++
и А.
Эти состояния отвечают отсутствию Na-проводимости. При де-

4.4.
АКТИВАЦИЯ
И
ИНАКТИВАЦИЯ НАТРИЕВОЙ ПРОВОДИМОСТИ
199
поляризации
мембраны происходит переход
А—>В
в
проводя-
щее состояние, связанный
с
перемещением заряда
z в
полеаф.
Из
состояния
В
активирующая частица может переходить
в не-
активное состояние
С.
Этот переход
не
связан
с
перемещением
заряда. После снятия деполяризации происходит переход
С —*
S
(
=
А
•
Са)
с
работой заряда 2аср#~. Схема этих процес-
сов показана
на
рис.
4.27.
Соответствующая система кинетиче-
ских уравнений имеет
вид
х
= kis —
k-ixcc
a
•+•
k~
2
y — k
2
x,
у = k-
3
u
—
k
3
y — k-
2
y + k
2
x,
Ы
= —
k-
3
U
+ k
3
y + k-
4
S
— k
4
UC
C
a,
x-f-y-f-u-r-s—1.
(4.67)
Здесь x,y,u,s
—
доли молекул
в
состояниях
А, В, С, S
(=
А
•
Са)
соответственно.
сс
а
—
концентрация Са
++
в
омываю-
щем растворе.
Как уже
сказано,
gs
a
= ky (см. стр. 196).
Исследова-
ние
изменений
gNa во
времени
сво-
дится
к
определению
y(t).
Считая,
что в
реакции
//
быстро
устанавливается равновесие
(это со-
гласуется
с
наблюдаемым влиянием
с
Са
на
кинетику активации), заменим
первое уравнение
(4.67)
на
(4.68)
Рис.
4.27.
Схема
процесса
активации
и
инактивации
Na-проводимости
[59].
где константа равновесия
Ki =
k-i/k{.
Константы
К[ и Кз,
очевидно,
не
свя-
заны
с
перемещением зарядов
в
поле
и
не
зависят
от
<р.
К
2
и
/С
4
должны зависеть
от
(р
согласно
фор
:
мулам
К
2
— q
2
exp
(-
=
q
t
exp {z
(4.69)
Система уравнений (4.67) приводится
к
виду
- 0
й
=
k-
t
[l
-
У
-
и].
fff)
и]
•
(4.70)
200
ГЛ. 4. НЕРВНЫЙ ИМПУЛЬС
Нужно найти корни г
и
г
2
характеристического уравнения си-
стемы
Комплексным корням соответствует переход системы в стацио-
нарное состояние в
результате
затухания колебаний около этого
состояния.
При фиксированном потенциале ионные токи не ис-
пытывают
затухающих
колебаний. Ищем решение
(4.70)
с ве-
щественными корнями Г\ и г
2
. Они отрицательны и решение за-
писывается следующим образом:
у (t) = a
1
+ b
1
exp(- | n \t) + с, ехр(- | r
2
|/),
и (t) = a
2
+ b
2
exp (- | r, | /) + c
2
exp (- | r
2
\
t).
(4>72)
Введя начальные значения y
0
, u
0
, г/о, «о и равновесные значения
Уоо,
и
х
, отвечающие
t—+
oo, получим
У
о
. _ Уо +
С]
r\ — r
2
'со - Уо).
/"2
—
и
аналогичные выражения для а
2
. ^2,
С2.
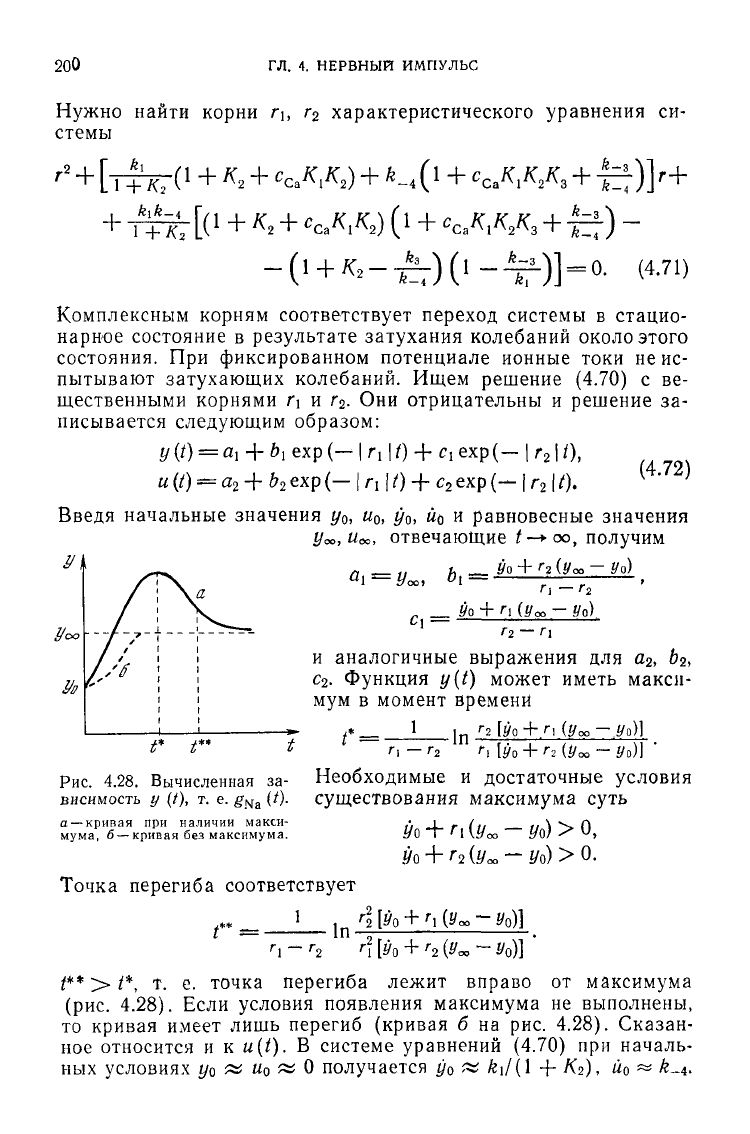
Функция г/(^) может иметь макси-
мум в момент времени
/* = •
1
-In
Г
2
[У0+/1 (Уоо — .Уо)]
Рис.
4.28. Вычисленная за-
висимость у (0, т. е. g
Na
(О-
а — кривая при наличии макси-
мума, 6 — кривая без максимума.
r
i —
r
2 f\ [Уо Н"
i~2
(Уоо — Уо)]
Необходимые и достаточные условия
существования максимума
суть
Уо
+ г, («/„ — г/о) > О,
Точка перегиба соответствует
In
4
[Уо
+
г
\
{Уоо
~
Уо)]
[У
о
+
~
У
о)]
Р* > /*, т. е.
точка
перегиба
лежит
вправо
от
максимума
(рис.
4.28).
Если
условия
появления
максимума
не
выполнены,
то
кривая
имеет
лишь
перегиб
(кривая
б на рис. 4.28).
Сказан-
ное
относится
и к u(t). В
системе
уравнений
(4.70) при
началь-
ных
условиях
г/о « «о ~ 0
получается
у
0
« k\/(\ -j- ^2), й
0
« k^.