Волькенштейн М.В. Общая биофизика
Подождите немного. Документ загружается.


§
5.6. ТЕОРИЯ СТАЦИОНАРНОГО МЫШЕЧНОГО СОКРАЩЕНИЯ 251
т. е. число одновременно работающих мостиков
убывает
с уве-
личением скорости v. Если v = О, то w = \\ если v =
y
max
=
=
b (1 — г) /г, то w = ш
т1п
= г:
^ =
Г
=
Т+7^7
*
(5
-
63)
Естественно, что изложенная теория
дает
результаты,
экви-
валентные теории Хаксли — Дещеревского (см. стр. 244). Сопо-
ставим формулы. Из уравнений
(5.38)
и
(5.48)
следует
£-Ц£.
(5.64)
Подставляя в уравнение
(5.34)
стационарное значение т =
=
nvlLk
2
из (5.35), находим
(±±b)
(5.65)
и
при п = О получаем стационарное отношение n/«
0
= w. При-
равнивая
его выражению (5.62), получаем
+
v).
(5.66)
Из
(5.60), (5.61),
(5.64)
и
(5.66)
находим
г-^_г_г-
(567)
Т
(5-68)
Таким
образом, можно выразить k\ и k
2
через молекулярные
постоянные.
В изложенной теории трение возникает в
результате
замыка-
ния
и размыкания мостиков, так как эти процессы
требуют
энер-
гии
активации. Коэффициент трения, естественно, оказывается
экспоненциально
зависящим от температуры. В макроскопиче-
ской
теории молекулярная модель заменяется формальной мо-
делью
Хаксли — Дещеревского — мостик либо тянет, либо тор-
мозит, хотя в действительности эти явления должны происходить
одновременно.
Преимущество излагаемой теории состоит в том,
что она согласуется с принципом микроскопической обратимости
и
непосредственно выражает кинетические константы через мо-
лекулярные параметры L, cos 9, G, f
0
. Тем самым объясняется
температурная зависимость b и, как мы увидим, f
0
.
В изложенной теории предполагается, что мостики замы-
каются независимо
друг
от
друга.
Насколько основательно это
предположение? Нет ли здесь кооперативности в том смысле,
что замыкание одного мостика влияет на замыкание
других?

252 ГЛ. 5.
МЕХАНОХИМИЧПСКИЕ
П1ЮЦЕССЫ
Идея
о кооперативных, аллостерических свойствах актина и мио-
зина
высказывалась не раз.
Введем
кооперативность в кинетику действия мостиков, пред-
положив, что свободная энергия активации G сама является
функцией
числа работающих мостиков
G = G
o
- wG' = G
o
- [г + (1 - г) f/f
0
] С.
(5.69)
Характеристическое уравнение получается в виде (Ь имеет преж-
нее значение с G, замененным на G
o
)-
(а + f) v = Ь ехр (^) ехр \^г~\ (/о - f>.
(5-70)
При
заметных значениях
G'/k^T
кривая v(f) оказывается вы-
пуклой, а не вогнутой. Для согласования с опытом
следует
предположить, что G' <C ksT. В этом
случае
(а
+
f)
v «
b
[l +
1=^(а
+ f)-^] (f
0
-
f),
(5.71)
т. е. кооперативность рассматриваемого типа вносит лишь незна-
чительную поправку в уравнение Хилла, чго не представляет
особого интереса.
Исходная идея изложенной теории состоит во влиянии внеш-
ней
силы / на активационный процесс замыкания мостика и кон-
формационного
превращения белка. В свое время Полиссар
предполагал экспоненциальную зависимость скорости некой не-
известной реакции, определяющей сокращение, от внешней
силы [127] (см. также [53]). Т. Хилл также вводил
сходную
экспоненциальную зависимость в своей теории фазового пере-
хода
в мышце [119]. Эти работы, однако, не давали возможности
получить характеристическое уравнение и истолковать его кон-
станты. Нельзя считать изложенную теорию молекулярной в ис-
тинном
смысле слова, так как она не
дает
реальной молекуляр-
ной
модели работы мышцы. Построение молекулярной теории
невозможно на современном уровне знаний о строении и взаи-
модействии мышечных сократительных и регуляторных белков
и
об их конформационных превращениях. Соответственно за-
дачи исследования структуры и свойств этих белков, исследова-
ния
их поведения в надмолекулярной мышечной системе осо-
бенно
актуальны.
§
6.7. ТЕОРИЯ ТЕРМОМЕХАНИЧЕСКИХ СВОЙСТВ МЫШЦЫ
В мышце, рассматриваемой как изолированная система, ра-
бота и тепло, выделяемые в элементарном акте, выражаются
через уменьшение химической энергии
UL + (/о = - Z И/ Ля,,
(5.72)

§
5.7. ТЕОРИЯ ТЕРМОМЕХЛЫИЧЕСКИХ СВОЙСТВ МЫШЦЫ 253
или,
согласно закону сохранения энергии,
^E
= T^S +
f
o
L+
1
Zv•iЬп
i
=
O,
(5.73)
i
причем ГА5 = q^. Изменение свободной энергии в рассмотрен-
ном
элементарном акте составляет, как мы видели,
fo'cosB
(см.
стр. 249). Имеем
AG = Д£ - Т AS = f
0
L + E \i
t
An
t
=
/„/
cos 9,
откуда
X
^
д
"<
(5-74)
Максимальная
сила fo (и, следовательно, Р
о
) должна слабо ра-
сти с температурой как химический потенциал, т. е. пропорцио-
нально
абсолютной температуре. Это согласуется с опытом.
Коэффициент
полезного действия мышцы, согласно изложен-
ной
теории, равен
I
—
i
Если
бы выполнялся закон сохранения энергии (5.73), то
q
^THS
=
-Ы cos9, A'=
x
l
При
cos 0 > 0 получаем
г)'>1,
т. е. мы приходим к противо-
речию.
Как
мы видели, проблема теплопродукции мышцы не проста
(см.
§ 5.4). Она не может быть решена на основе столь про-
стых соображений. Необходимо
учесть
запасание энергии и раз-
личные способы ее выделения. Соответствующая модельная тео-
рия
предложена в работе [66].
Авторы
этой работы считают, что
энергия,
освобождаемая при сокращении мышцы, должна сла-
гаться из запасенной энергии, диссипируемой в центрах одного
типа, и энергии, получаемой за счет распада АТФ в центрах дру-
гого типа. Теплота укорочения получается из диссипированной
энергии.
Предполагается, тем самым, что в
двух
типах центров
действуют
две различные актомиозиновые АТФ-азы. Факты,
свидетельствующие в пользу такого предположения, состоят в
следующем.
1. Центры Р и S, связывающие две молекулы АТФ (или
АДФ), находятся в головках миозина [128]; в Р-центре лими-
тирующая стадия для Mg-зависимой АТФ-азы есть отделение

254 ГЛ. 5. МЕХАНОХИМИЧЕСКИЕ ПРОЦЕССЫ
продукта, т. е. АДФ и Ф
н
от миозина [129], в S-центре лимити-
рующая стадия есть гидролиз АТФ [55].
2. Актин активирует гидролиз АТФ в S-центре миозина.
3. Реакция Р-центра с АТФ необходима для сокращения.
4. Актин ускоряет освобождение продукта Р-центрами [129].
5. Энергия запасается в комплексе миозин —продукт. Факты,
свидетельствующие об этом, приведены в работе [66]. И, на-
конец,
6. При контакте с актином в присутствии АТФ миозин не
образует «комплекса окоченения» (rigor complex) с актином,
т. е. миозин всегда нагружен продуктом и (или) субстратом.
Из
этих шести положений и из предположений, приведенных
на
стр. 237,
следует
модель работы актомиозиновой АТФ-азы [66]:
быстро _ т
М + АТФ *• М —*• М + АДФ + Ф„, (I)
М + АТФ + FA —V м • (АТФ) • FA —^ М + FA + АДФ + Ф
н
+ G, (II)
h
—>•
М + FA + АТФ + G
2
. (Ill)
Здесь М — миозин, М—комплекс миозин — продукт, т. е. комп-
лекс М-АДФ-Фн, запасающий энергию, FA —F-актин, G\ — сво-
бодная энергия, выделяющаяся при гидролизе одной молекулы
АТФ, G
2
— энергия, выделяемая М, т — время жизни запасенной
энергии,
/о, /ь h — реакционные потоки (скорости реакций).
Процесс (I) представляет реакции в Р-центре. Энергия, за-
пасенная
в комплексе М, выделяется в процессах (II) или (III).
При
исчерпании запасенной энергии комплекс М освобождает
АДФ и Ф
н
и затем вновь реагирует с АТФ и запасает энергию.
Процессы (II) и (III) происходят при сокращении мышцы, в
Р-центре. АТФ-аза катализирует гидролиз АТФ в S-центре, ис-
пользуя часть запасенной энергии, генерируемой в Р-центре.
Это — своеобразная аллостерическая система.
Изменение
свободной энергии в единицу времени в процес-
сах (II) и (III) равно
— G = /iG, -f /
2
G
2
.
(5.76)
В стационарном режиме
/
0
= /
1 +
/
2
.
(5.77)
Поток
J\ выражает скорость гидролиза АТФ в S-центре. Он ра-
вен произведению числа комбинаций актин —миозин, создаю-
щих напряжение (числа работающих мостиков), на число оборо-
тов реакции (II). Первый сомножитель пропорционален нагрузке

§
5.7. ТЕОРИЯ ТЕРМОМЕХАНИЧЕСКИХ СВОЙСТВ МЫШЦЫ 255
Р, второй линейно зависит от скорости укорочения V. Поэтому
J
i
= P(K
1
V + K
2
)^K
l
PV + K
2
P, (5.78)
где К\ и К
2
— константы. Перепишем (5.76) в виде
-G
=
/oG,-/
2
(G,-G
2
)
(5.79)
и
предположим, что поток /г линейно зависит от
«сродства»
А = G\ — G
2
и нагрузки
-]
2
= с
х
А + с
2
Р. (5.80)
Наконец,
пусть скорость укорочения линейно зависит от тех же
величин
(5.81)
Из
соотношений (5.77), (5.78), (5.80) и (5.81)
следует
уравнение
Хилла в форме
(«-«о
Определим коэффициенты с,. При Р = 0 имеем V = К
гаах
,
А
==
А
т
, h = /о и
(5.83)
При
P —
PQ
имеем У = 0, Л = Л
0
и
С
4
—
— С
3
— — -| —-, (5.84)
•^т "о
Подставляя эти значения с
г
в (5.82), находим
т. е. снова получаем уравнение Хилла, в котором
a
= J
0
/K
l
V
max
, Ь =
/
0
//С
1
Р
0
.
(5.87)
Одновременно имеем
А = -^А
0
+ ^-А
т
. (5.88)
"о ' max
При
выводе уравнения Хилла сократились все члены, содержав-
шие
К
2
и значения
«сродства»
Л
о
и А
т
при нулевой и максималь-

256 ГЛ. 5. МЕХАНОХИМИЧЕСКИЕ ПРОЦЕССЫ
ной
скорости укорочения. Сопоставление с теорией, изложенной
в
§ 5.6, показывает, что в расчете на один мостик
(5-89)
/Ci
= (/o^)~
!
.
(5.90)
т. е. уравнение
(5.78)
имеет вид
Считая,
что энтальпия гидролиза АТФ примерно равна соот-
ветствующей свободной энергии [58], находим количество тепла,
выделяемое в единицу времени
Q =
-G-PV=J
l
G
1
+
J
2
G
2
-PV.
(5.92)
При
Р = Ро и V = 0 имеем
Q
o
= Ki&Po [G, - A,(l -
Кг/Kib)].
(5.93)
По
определению скорость выделения тепла укорочения равна
Q-Q
Q
= aV.
(5.94)
Из
приведенных формул и уравнения Хилла получим
(Б.95)
где х = P/Po.
Для нахождения Д
т
и Л
о
это сложное уравнение приравни-
вается выражению
(5.27)
а/Ро = 0,16 + 0,18 х при х = 0 и
х = 0,5. В области 0 •< х < 0,5 получается хорошее согласие с
результатами Хилла. То же относится к полной скорости выде-
ления
энергии. Эффективность мышцы (как функции х)
Е/Р
о
х+а/Р
0
(5
.
96)
вычисленная
на основании излагаемой теории, хорошо согла-
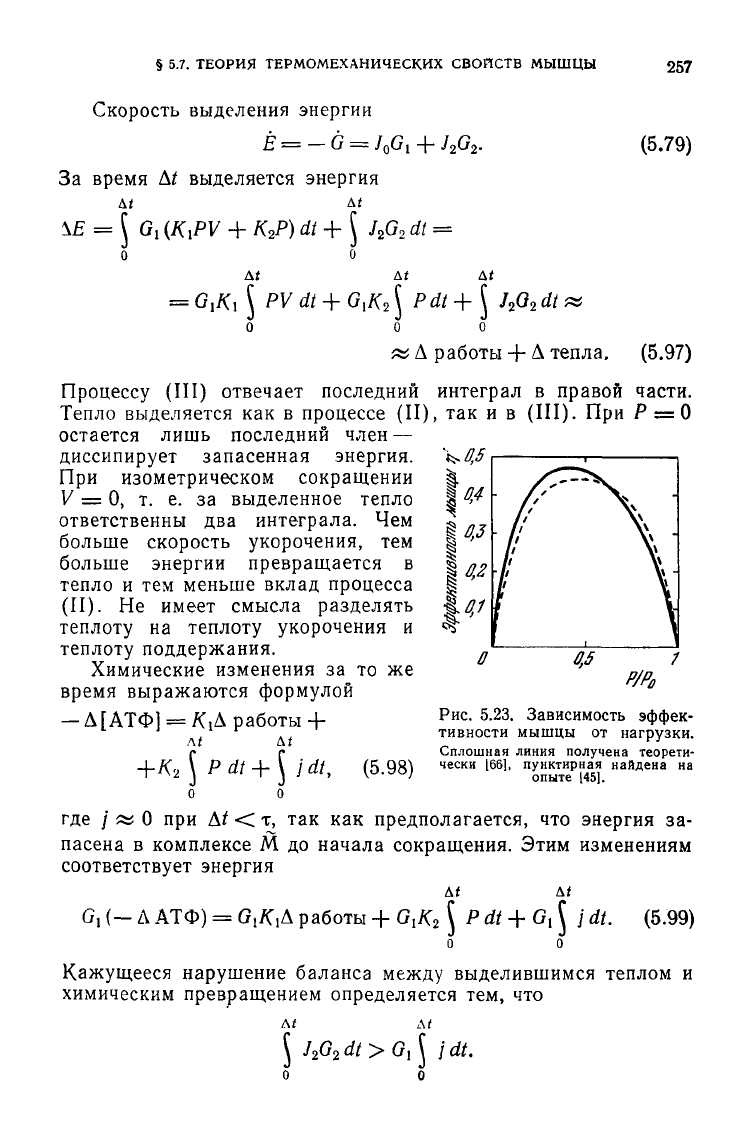
суется с найденной Хиллом (рис.
5.23).
Из
экспериментальных данных
следует,
что Gj, т. е. измене-
ние
свободной энергии при гидролизе одной молекулы АТФ есть
величина порядка 10 ккал/моль.
§
5.7.
ТЕОРИЯ ТЕРМОМЕХАНИЧЕСКИХ СВОЙСТВ МЫШЦЫ
Скорость
выделения энергии
257
(5.79)
За
\Е
время
L
At
0
\t выделяется
(KiPV
+ K
2
P)
At
=
0,/c,
5
PV
0
энергия
dt-
dt-\
At
0
\-GiK
>G
2
At
Л
0
A
=
At
Pdt+
jj J
0
А работы
+
•
jA«
А тепла.
(5
.97)
Процессу
(III)
отвечает последний интеграл
в
правой части.
Тепло выделяется
как в
процессе (II),
так и в
(III).
При Р = 0
остается лишь последний член
—
диссипирует запасенная энергия.
При
изометрическом сокращении
V
= О, т. е. за
выделенное тепло
ответственны
два
интеграла.
Чем
больше скорость укорочения,
тем
'bs.ff.5
|
0,4
0,3•
больше энергии превращается
в
%
#£
тепло
и тем
меньше вклад процесса
|
(II).
Не
имеет смысла разделять
||^
теплоту
на
теплоту укорочения
и #
теплоту поддержания.
„
Химические изменения
за то же
время выражаются формулой
j
=
/CiA работы
+
At
At
+/C
2
J PA+5
/A,
(5.98)
0,5
Р/Ро
Рис.
5.23. Зависимость
эффек-
тивности мышцы от нагрузки.
Сплошная линия
получена
теорети-
чески
[661,
пунктирная найдена
на
опыте
145].
6
где
/ « 0 при А^ < т, так как
предполагается,
что
энергия
за-
пасена
в
комплексе
М до
начала сокращения. Этим изменениям
соответствует энергия
At At
G,
(-
А АТФ)
= G^A
работы
+ G,ff
2
\ Р А +
G,
J / dt.
(5.99)
о
о
Кажущееся нарушение баланса
между
выделившимся теплом
и
химическим превращением определяется
тем, что
At
At
J
2
G
2
A > G, J
/
dt.

258
ГЛ. 5.
МЕХАНОХИМИЧЕСКИЕ
ПРОЦЕССЫ
Разность этих
двух
интегралов равна заранее запасенной
энер-
гии.
Время жизни запасенной энергии
запасенная
энергия
так как /2G2 есть скорость освобождения запасенной энергии.
Численные оценки
дают
0,2 с ^ т ^ 0,9 с в интервале
V
max
^
^ V > 0, т. е. 0 < Р ==£ Р
о
.
В опытах, проводимых за время, меньшее 0,2 с, / » 0 и не
наблюдается соответствия
между
химическим превращением и
выделившимся теплом.
Теория, предложенная в работе [66], хорошо объясняет основ-
ные факты, относящиеся к теплопродукции мышцы. Однако эта
теория основана на модели, хотя и хорошо аргументированной,
но
все же содержащей ряд постулативных положений. Подлин-
ный
молекулярный механизм процесса остается неизвестным.
Путь к его познанию идет через детальное изучение строения
и
свойств мышечных белков.
§
5.8. ИНИЦИАЦИЯ МЫШЕЧНОГО СОКРАЩЕНИЯ
Предшествующее изложение ограничивалось стационарным
сокращением мышцы. Не меньший, если не больший интерес
представляют нестационарные кинетические процессы в мышеч-
ном
волокне.
Первая стадия сокращения — его инициация ионами Са
++
, вы-
ходящими из саркоплазматического ретикулума под влиянием
электрического импульса. Импульс передается либо по аксонам
нервных клеток, либо путем прямого электрического стимули-
рования мембраны мышечного волокна. При стимуляции мы-
шечного волокна коротким электрическим импульсом возникает
напряжение,
развитие которого во времени выражается кривой
с максимумом и перегибом [130]. При стимуляции серией корот-
ких импульсов напряжение выражается совокупностью
пико-
образных кривых, причем каждый последующий максимум
лежит выше предыдущего, пока не достигается стационарный
режим. При больших частотах пики сливаются, образуя
экспо-
ненциальную кривую гладкого тетануса [131]. В этих условиях
потенциал на мембране волокна меняется в течение опыта. В ра-
боте [132] были проведены измерения изометрического напряже-
ния
при фиксированном потенциале. Развитие напряжения во
времени описывалось суммой
двух
экспонент. Из температурной
зависимости скорости процесса получаются барьеры d »
19—20
и
G
2
« 15—16 ккал/моль. Вместе с тем высота барьера для диф-
фузии Са
++
из ретикулума, определяемая по скорости насыщения
ретикулума ионами Са
++
[133], составляет
18—20
ккал/моль.

§
5.8.
ИНИЦИАЦИЯ МЫШЕЧНОГО СОКРАЩЕНИЯ
259
В работе
[30]
рассмотрена соответствующая двухбарьерная
модель
—
первый барьер
G\
соответствует диффузии
Са^,
второй
G
2
—
замыканию мостика, расщеплению
АТФ
и
конформацион-
ному
переходу.
Этот барьер соответствует рассмотренному
в
предыдущем разделе. Профиль свободной энергии изображен
на
рис.
5.24.
Минимум
/ на
рис.
5.24
соответствует состоянию
си-
стемы,
в
котором большая часть Са
++
сосредоточена
в
мембране
ретикулума,
//
—
состоянию,
в
котором Са
++
находится
в
сарко-
плазме
или
на
мостике
до его
замыкания,
/// — на
сработавшем
мостике.
На
основании опытов
по
локальной деполяризации мышеч-
ного волокна предполагается,
что
напряжение
ф,
приложенное
к
его
мембране, передается
на
мембраны саркоплазматического
ретикулума. Ранее
уже
были проана-
лизированы
различные модели
уча-
стия
ионов Са
++
в
механизме измене-
ния
ионной проницаемости мембран
нервных волокон
под
действием элек-
трического поля
(см.
§
4.4).
Пусть
Хи х
2
,
x%
—
числа ионов Са
44
в
состояниях
/, //, ///;
N
—
их
сум-
марное количество. Предполагается,
что
Хг
<С Х\
-f- х
2
и,
следовательно,
N
« xi
-\-
х
2
.
Кинетические уравнения
имеют
вид
Xi
— k-iN
\k\-\-
k-\)X\, Л
Рис. 5.24.
Профиль свобод-
Xi
= k
2
N —
k
2
xi
-
k-
2
x
3
.
)
{
'
ной
энергии.
Константы
скорости указаны
на
рис.
5.24.
Решение уравнений
(5.100)
записывается следующим образом:
xi
(0
= xi» +
(*i
(0)
-
*i
„о)
ехр
Yi*,
|
*3(0
=
*3<x,
+
aexpYi/
+
PexpY2*,
J
(5.101)
где
YI=—(^i
+ ^-0»
Y2=^~2,
-«юс
и
х
3<
х»
—
стационарные значения
х\
и х
3
.
Константы
аир
находятся
из
начальных условий. Имеем
=
N
T
^
K7
.
(5.102)
где /Ci,
2
= k-\
t
-2/^1,2-
В
принятой модели
/С]
=
ехр (—
q>z&~/RT)\
(5.103)
здесь
г
—
валетность частицы, #"
—
число Фарадея. Таким обра-
зом,
k\, k-\
зависят
от ф.
Можно
считать,
что
при
изометрическом сокращении напря-
жение,
стимулированное импульсом, пропорционально числу

260
гл
- 5-
МЕХАНОХИМИЧЕСКИЕ ПРОЦЕССЫ
замкнутых мостиков, линейно зависящему
от х
3
.
Таким образом,
g[xAt)-x
3
],
(5.104)
где
g —
константа,
х
3
—
значение
х
3
при
потенциале покоя.
Из
(5.102)
и
(5.104)
легко найти стационарное напряжение
Рос.
Вводя безразмерные переменные
n^P^KJgN,
ф
=
ф^"//?Г,
получаем
я
=
т-г—L
г
—т-г—-,
г-
(5.105)
1
+
ехр
(—
2ф)
I +
ехр
(—
гфо)
'
где
фо
—
потенциал покоя
в
единицах
RT/@~.
Находим
dn
—
-,
ех
Р (—
г
Ф>
/
В точке перегиба кривой я(ф) имеем
ехр(—zq>)
— 1, т. е.
dn
г
—г.
(5.107)
перегиб
4
Опыт дает значение этой величины, близкое
к 1
[132]. Следова-
тельно,
z fv 4, что
может означать перемещение двухвалентных
ионов
Са
++
парами. При длительной фиксации потенциала пара-
метры системы меняются один раз —в начале деполяризации.
Начальные условия имеют
вид
х,(0)
=
*,, *,(0)
=
je
3
.
(5.108)
Константы
а и Р во
втором уравнении
(5.101)
равны
а
TJ—~—\-*зоо
—
-*з/>
Р
—ц—v^noo
—
*з)'
^,o.luy^
Из
(5.102),
(5.104)
и
(5.109)
следует
(5.110)
Такой характер зависимости
P(t) при
фиксированном потен-
циале согласуется
с
опытом.
При
кратковременной деполяризации внешние параметры
ме-
няются дважды:
в
момент наложения потенциала
( = 0 и в мо-
мент
его
снятия
t = т.
Допустим
для
простоты,
что
|у1,г|т<С
1.
На
отрезке времени
0
—
т
переменные
ведут
себя согласно
си-
стеме уравнений
(5.100)
при
начальных условиях (5.108). Огра-
ничиваясь линейными членами
по
\1,2Т, находим
х\
(т)
«
xi
+
(х
х
—
х
н
) J
[
(5.111)