Berg J.M., Tymoczko J.L., Stryer L. Biochemistry
Подождите немного. Документ загружается.


direct repair
base-excision repair
nucleotide-excision repair
trinucleotide repeat
Ames test
III. Synthesizing the Molecules of Life 27. DNA Replication, Recombination, and Repair
Problems
1.
Activated intermediates. DNA polymerase I, DNA ligase, and topoisomerase I catalyze the formation of
phosphodiester bonds. What is the activated intermediate in the linkage reaction catalyzed by each of these
enzymes? What is the leaving group?
See answer
2.
Fuel for a new ligase. Whether the joining of two DNA chains by known DNA ligases is driven by NAD
+
or ATP
depends on the species. Suppose that a new DNA ligase requiring a different energy donor is found. Propose a
plausible substitute for NAD
+
or ATP in this reaction.
See answer
3.
Life in a hot tub. An archaeon (Sulfolobus acidocaldarius) found in acidic hot springs contains a topoisomerase that
catalyzes the ATP-driven introduction of positive supercoils into DNA. How might this enzyme be advantageous to
this unusual organism?
See answer
4.
A cooperative transition. The transition from B-DNA to Z-DNA occurs over a small change in the superhelix
density, which shows that the transition is highly cooperative.
(a) Consider a DNA molecule at the midpoint of this transition. Are B- and Z-DNA regions frequently intermingled
or are there long stretches of each?
(b) What does this finding reveal about the energetics of forming a junction between the two kinds of helices?
(c) Would you expect the transition from B- to A-DNA to be more or less cooperative than the one from B- to Z-
DNA? Why?
See answer
5.
Molecular motors in replication. (a) How fast does template DNA spin (expressed in revolutions per second) at an
E. coli replication fork? (b) What is the velocity of movement (in micrometers per second) of DNA polymerase III
holoenzyme relative to the template?
See answer

6.
Wound tighter than a drum. Why would replication come to a halt in the absence of topoisomerase II?
See answer
7.
Telomeres and cancer. Telomerase is not active in most human cells. Some cancer biologists have suggested that
activation of the telomerase gene would be a requirement for a cell to become cancerous. Explain why this might be
the case.
See answer
8.
Nick translation. Suppose that you wish to make a sample of DNA duplex highly radioactive to use as a DNA probe.
You have a DNA endonuclease that cleaves the DNA internally to generate 3
-OH and 5 -phosphate groups, intact
DNA polymerase I, and radioactive dNTPs. Suggest a means for making the DNA radioactive.
See answer
9.
Revealing tracks. Suppose that replication is initiated in a medium containing moderately radioactive tritiated
thymine. After a few minutes of incubation, the bacteria are transferred to a medium containing highly radioactive
tritiated thymidine. Sketch the autoradiographic pattern that would be seen for (a) undirectional replication and (b)
bidirectional replication, from a single origin.
See answer
10.
Mutagenic trail. Suppose that the single-stranded RNA from tobacco mosaic virus was treated with a chemical
mutagen, that mutants were obtained having serine or leucine instead of proline at a specific position, and that
further treatment of these mutants with the same mutagen yielded phenylalanine at this position.
(a) What are the plausible codon assignments for these four amino acids?
(b) Was the mutagen 5-bromouracil, nitrous acid, or an acridine dye?
See answer
11.
Induced spectrum. DNA photolyases convert the energy of light in the near ultraviolet or visible region (300 500
nm) into chemical energy to break the cyclobutane ring of pyrimidine dimers. In the absence of substrate, these
photoreactivating enzymes do not absorb light of wavelengths longer than 300 nm. Why is the substrate-induced
absorption band advantageous?
See answer
Mechanism Problems
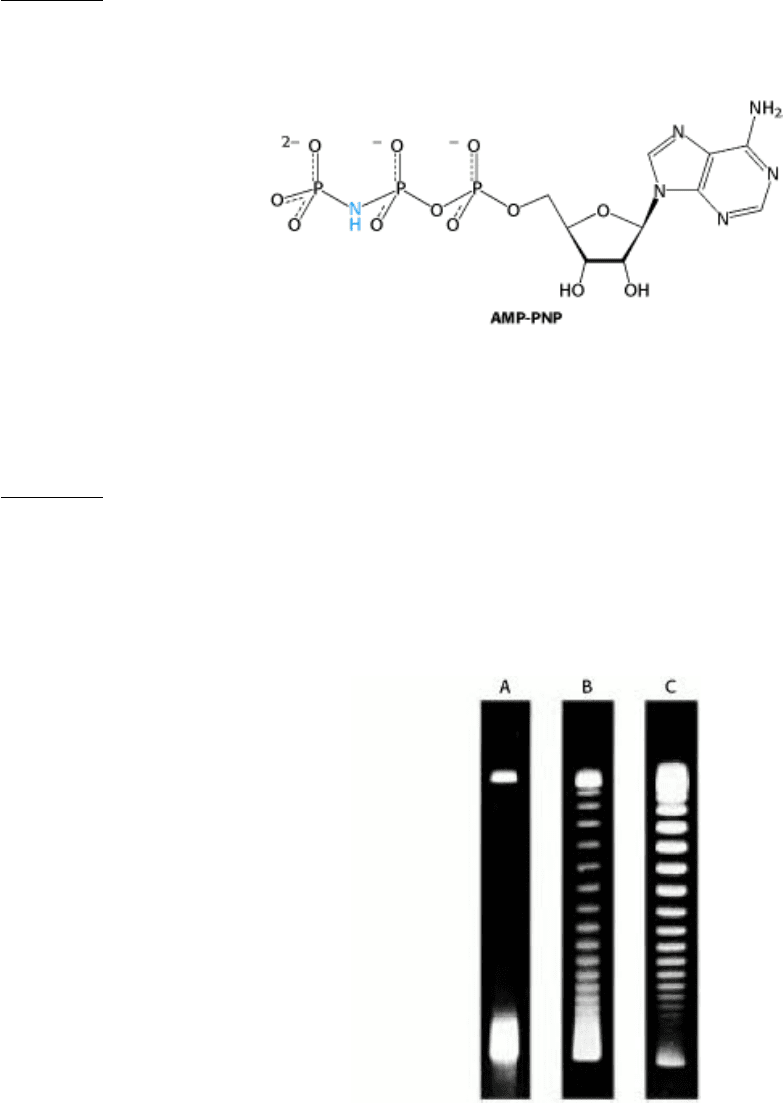
12.
AMP-induced relaxation. DNA ligase from E. coli relaxes supercoiled circular DNA in the presence of AMP but
not in its absence. What is the mechanism of this reaction, and why is it dependent on AMP?
See answer
13.
A revealing analog. AMP-PNP, the β,γ-imido analog of ATP, is hydrolyzed very slowly by most ATPases.
The addition of AMP-PNP to topoisomerase II and circular DNA leads to the negative supercoiling of a single
molecule of DNA per enzyme. DNA remains bound to the enzyme in the presence of this analog. What does this
finding reveal about the catalytic mechanism?
See answer
Data Interpretation and Chapter Integration Problems
14.
Like a ladder. Circular DNA from SV40 virus was isolated and subjected to gel electrophoresis. The results are
shown in lane A (the control) of the adjoining gel patterns.
(a) Why does the DNA separate in agarose gel electrophoresis? How does the DNA in each band differ?
The DNA was then incubated with topoisomerase I for 5 minutes and again analyzed by gel electrophoresis with
the results shown in lane B.
(b) What types of DNA do the various bands represent?
Another sample of DNA was incubated with topoisomerase I for 30 minutes and again analyzed as shown in lane C.
(c) What is the significance of the fact that more of the DNA is in slower moving forms?
See answer
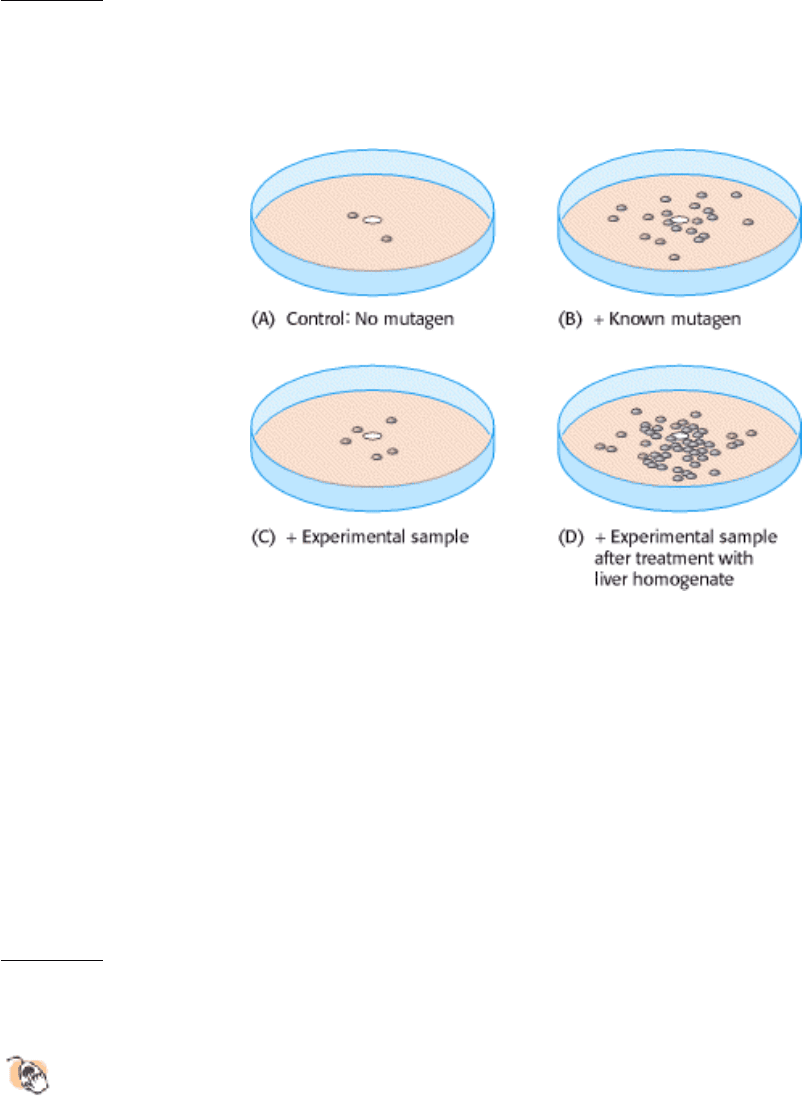
15.
Ames test. The adjoining illustration shows four petri plates used for the Ames test. A piece of filter paper (white
circle in the center of each plate) was soaked in one of four preparations and then placed on a petri plate. The four
preparations contained
(A) purified water (control), (B) a known mutagen, (C) a chemical whose mutagenicity is under investigation, and
(D) the same chemical after treatment with liver homogenate. The number of revertants, visible as colonies on the
petri plates, was determined in each case.
(a) What was the purpose of the control plate, which was exposed only to water?
(b) Why was it wise to use a known mutagen in the experimental system?
(c) How would you interpret the results obtained with the experimental compound?
(d) What liver components would you think are responsible for the effects observed part D?
See answer
Media Problem
16.
Cre-ative duplexes. Site specific recombinases like Cre require that the sequence between the recombinase
binding sites be identical in the two DNA molecules that are to be recombined. Examine the structures of the
various Cre-DNA complexes in the Structural Insights module on Cre recombinase. How might the
recombination mechanism check these sequences to ensure they are identical in the parent DNA molecules?

III. Synthesizing the Molecules of Life 27. DNA Replication, Recombination, and Repair
Selected Readings
Where to begin
A. Kornberg. 1988. DNA replication J. Biol. Chem. 263: 1-4. (PubMed)
R.E. Dickerson. 1983. The DNA helix and how it is read Sci. Am. 249: (6) 94-111.
J.C. Wang. 1982. DNA topoisomerases Sci. Am. 247: (1) 94-109. (PubMed)
T. Lindahl. 1993. Instability and decay of the primary structure of DNA Nature 362: 709-715. (PubMed)
C.W. Greider and E.H. Blackburn. 1996. Telomeres, telomerase, and cancer Sci. Am. 274: (2) 92-97. (PubMed)
Books
Kornberg, A., and Baker, T. A., 1992. DNA Replication (2d ed.).W. H. Freeman and Company.
Bloomfield, V. A., Crothers, D., Tinoco, I., and Hearst, J., 2000. Nucleic Acids: Structures, Properties and Functions.
University Science Books.
Friedberg, E. C., Walker, G. C., Siede, W., 1995. DNA Repair and Mutagenesis. American Society for Microbiology.
Cozzarelli, N. R., and Wang, J. C. (Eds.), 1990. DNA Topology and Its Biological Effects. Cold Spring Harbor
Laboratory Press.
DNA structure
T.K. Chiu and R.E. Dickerson. 2000. 1 Å crystal structures of B-DNA reveal sequence-specific binding and groove-
specific bending of DNA by magnesium and calcium J. Mol. Biol. 301: 915-945. (PubMed)
A. Herbert and A. Rich. 1999. Left-handed Z-DNA: Structure and Function Genetica 106: 37-47. (PubMed)
R.E. Dickerson. 1992. DNA Structure from A to Z Methods Enzymol 211: 67-111. (PubMed)
J.R. Quintana, K. Grzeskowiak, K. Yanagi, and R.E. Dickerson. 1992. Structure of a B-DNA decamer with a central T-A
step: C-G-A-T-T-A-A-T-C-G J. Mol. Biol. 225: 379-395. (PubMed)
N. Verdaguer, J. Aymami, F.D. Fernandez, I. Fita, M. Coll, D.T. Huynh, J. Igolen, and J.A. Subirana. 1991. Molecular
structure of a complete turn of A-DNA J. Mol. Biol. 221: 623-635. (PubMed)
DNA topology and topoisomerases
D. Sikder, S. Unniraman, T. Bhaduri, and V. Nagaraja. 2001. Functional cooperation between topoisomerase I and single
strand DNA-binding protein J. Mol. Biol. 306: 669-679. (PubMed)
Z. Yang and J.J. Champoux. 2001. The role of histidine 632 in catalysis by human topoisomerase I J. Biol. Chem. 276:
677-685. (PubMed)
J.M. Fortune and N. Osheroff. 2000. Topoisomerase II as a target for anticancer drugs: When enzymes stop being nice
Prog. Nucleic Acid Res. Mol. Biol. 64: 221-253. (PubMed)
R.J. Isaacs, S.L. Davies, M.I. Sandri, C. Redwood, N.J. Wells, and I.D. Hickson. 1998. Physiological regulation of
eukaryotic topoisomerase II Biochim. Biophys. Acta 1400: 121-137. (PubMed)
J.C. Wang. 1996. DNA topoisomerases Annu. Rev. Biochem. 65: 635-692. (PubMed)
J.C. Wang. 1998. Moving one DNA double helix through another by a type II DNA topoisomerase: The story of a
simple molecular machine Q. Rev. Biophys. 31: 107-144. (PubMed)
C.L. Baird, T.T. Harkins, S.K. Morris, and J.E. Lindsley. 1999. Topoisomerase II drives DNA transport by hydrolyzing
one ATP Proc. Natl. Acad. Sci. USA 96: 13685-13690. (PubMed) (Full Text in PMC)
A.V. Vologodskii, S.D. Levene, K.V. Klenin, K.M. Frank, and N.R. Cozzarelli. 1992. Conformational and
thermodynamic properties of supercoiled DNA J. Mol. Biol. 227: 1224-1243. (PubMed)
L.M. Fisher, C.A. Austin, R. Hopewell, M. Margerrison, M. Oram, S. Patel, D.B. Wigley, G.J. Davies, E.J. Dodson, A.
Maxwell, and G. Dodson. 1991. Crystal structure of an N-terminal fragment of the DNA gyrase B protein Nature 351:
624-629. (PubMed)
Mechanism of replication
M.J. Davey and M. O'Donnell. 2000. Mechanisms of DNA replication Curr. Opin. Chem. Biol. 4: 581-586. (PubMed)
J.L. Keck and J.M. Berger. 2000. DNA replication at high resolution Chem. Biol. 7: R63-R71. (PubMed)
T.A. Kunkel and K. Bebenek. 2000. DNA replication fidelity Annu. Rev. Biochem. 69: 497-529. (PubMed)
S. Waga and B. Stillman. 1998. The DNA replication fork in eukaryotic cells Annu. Rev. Biochem. 67: 721-751.
(PubMed)
K.J. Marians. 1992. Prokaryotic DNA replication Annu. Rev. Biochem. 61: 673-719. (PubMed)
DNA polymerases and other enzymes of replication
U. Hubscher, H.P. Nasheuer, and J.E. Syvaoja. 2000. Eukaryotic DNA polymerases: A growing family Trends Biochem.
Sci. 25: 143-147. (PubMed)
S. Doublié, S. Tabor, A.M. Long, C.C. Richardson, and T. Ellen-berger. 1998. Crystal structure of a bacteriophage T7
DNA replication complex at 2 2 Å resolution Nature 391: 251-258. (PubMed)
B. Arezi and R.D. Kuchta. 2000. Eukaryotic DNA primase Trends Biochem. Sci. 25: 572-576. (PubMed)
J. Jager and J.D. Pata. 1999. Getting a grip: Polymerases and their substrate complexes Curr. Opin. Struct. Biol. 9: 21-
28. (PubMed)
T.A. Steitz. 1999. DNA polymerases: Structural diversity and common mechanisms J. Biol. Chem. 274: 17395-17398.
(PubMed)
L.S. Beese, V. Derbyshire, and T.A. Steitz. 1993. Structure of DNA polymerase I Klenow fragment bound to duplex
DNA Science 260: 352-355. (PubMed)
C.S. McHenry. 1991. DNA polymerase III holoenzyme: Components, structure, and mechanism of a true replicative
complex J. Biol. Chem. 266: 19127-19130. (PubMed)
X.P. Kong, R. Onrust, M. O'Donnell, and J. Kuriyan. 1992. Three-dimensional structure of the beta subunit of E. coli
DNA polymerase III holoenzyme: A sliding DNA clamp Cell 69: 425-437. (PubMed)
A.H. Polesky, T.A. Steitz, N.D. Grindley, and C.M. Joyce. 1990. Identification of residues critical for the polymerase
activity of the Klenow fragment of DNA polymerase I from Escherichia coli J. Biol. Chem. 265: 14579-14591.
(PubMed)
J.Y. Lee, C. Chang, H.K. Song, J. Moon, J.K. Yang, H.K. Kim, S.T. Kwon, and S.W. Suh. 2000. Crystal structure of
NAD(+)-dependent DNA ligase: Modular architecture and functional implications EMBO J. 19: 1119-1129. (PubMed)
D.J. Timson and D.B. Wigley. 1999. Functional domains of an NAD
+
-dependent DNA ligase J. Mol. Biol. 285: 73-83.
(PubMed)
A.J. Doherty and D.B. Wigley. 1999. Functional domains of an ATP-dependent DNA ligase J. Mol. Biol. 285: 63-71.
(PubMed)
P.H. von Hippel and E. Delagoutte. 2001. A general model for nucleic acid helicases and their "coupling" within
macromolecular machines Cell 104: 177-190. (PubMed)
B.K. Tye and S. Sawyer. 2000. The hexameric eukaryotic MCM helicase: Building symmetry from nonidentical parts J.
Biol. Chem. 275: 34833-34836. (PubMed)
K.J. Marians. 2000. Crawling and wiggling on DNA: Structural insights to the mechanism of DNA unwinding by
helicases Structure Fold Des. 5: R227-R235.
P. Soultanas and D.B. Wigley. 2000. DNA helicases: 'Inching forward' Curr. Opin. Struct. Biol. 10: 124-128. (PubMed)
F. Bachand and C. Autexier. 2001. Functional regions of human telomerase reverse transcriptase and human telomerase
RNA required for telomerase activity and RNA-protein interactions Mol. Cell Biol. 21: 1888-1897. (PubMed) (Full Text
in PMC)
T.M. Bryan and T.R. Cech. 1999. Telomerase and the maintenance of chromosome ends Curr. Opin. Cell Biol. 11: 318-
324. (PubMed)
J.D. Griffith, L. Comeau, S. Rosenfield, R.M. Stansel, A. Bianchi, H. Moss, and T. de Lange. 1999. Mammalian
telomeres end in a large duplex loop Cell 97: 503-514. (PubMed)
M.J. McEachern, A. Krauskopf, and E.H. Blackburn. 2000. Telomeres and their control Annu. Rev. Genet. 34: 331-358.
(PubMed)
Recombinases
G.D. Van Duyne. 2001. A structural view of cre-loxp site-specific recombination Annu. Rev. Biophys. Biomol. Struct.
30: 87-104. (PubMed)
Y. Chen, U. Narendra, L.E. Iype, M.M. Cox, and P.A. Rice. 2000. Crystal structure of a Flp recombinase-Holliday
junction complex: Assembly of an active oligomer by helix swapping Mol. Cell 6: 885-897. (PubMed)
N.L. Craig. 1997. Target site selection in transposition Annu. Rev. Biochem. 66: 437-474. (PubMed)
D.N. Gopaul, F. Guo, and G.D. Van Duyne. 1998. Structure of the Holliday junction intermediate in Cre-loxP site-
specific recombination EMBO J. 17: 4175-4187. (PubMed)
D.N. Gopaul and G.D. Duyne. 1999. Structure and mechanism in site-specific recombination Curr. Opin. Struct. Biol. 9:
14-20. (PubMed)
Mutations and DNA repair
R.J. Michelson and T. Weinert. 2000. Closing the gaps among a web of DNA repair disorders Bioessays 22: 966-969.
(PubMed)
L. Aravind, D.R. Walker, and E.V. Koonin. 1999. Conserved domains in DNA repair proteins and evolution of repair
systems Nucleic Acids Res. 27: 1223-1242. (PubMed) (Full Text in PMC)
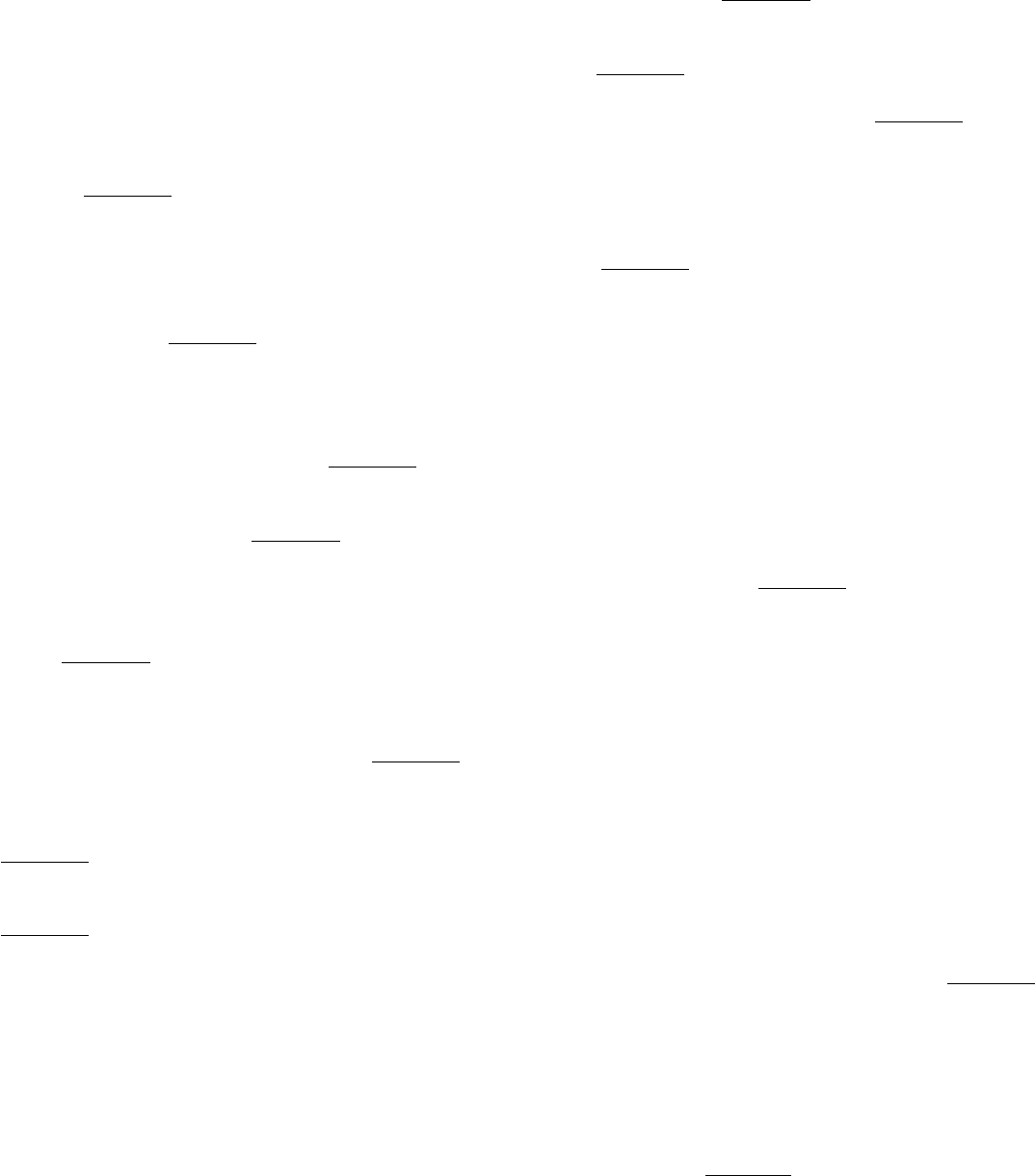
C.D. Mol, S.S. Parikh, C.D. Putnam, T.P. Lo, and J.A. Tainer. 1999. DNA repair mechanisms for the recognition and
removal of damaged DNA bases Annu. Rev. Biophys. Biomol. Struct. 28: 101-128. (PubMed)
S.S. Parikh, C.D. Mol, and J.A. Tainer. 1997. Base excision repair enzyme family portrait: Integrating the structure and
chemistry of an entire DNA repair pathway Structure 5: 1543-1550. (PubMed)
D.G. Vassylyev and K. Morikawa. 1997. DNA-repair enzymes Curr. Opin. Struct. Biol. 7: 103-109. (PubMed)
G.L. Verdine and S.D. Bruner. 1997. How do DNA repair proteins locate damaged bases in the genome? Chem. Biol. 4:
329-334. (PubMed)
R.P. Bowater and R.D. Wells. 2000. The intrinsically unstable life of DNA triplet repeats associated with human
hereditary disorders Prog. Nucleic Acid Res. Mol. Biol. 66: 159-202. (PubMed)
C.J. Cummings and H.Y. Zoghbi. 2000. Fourteen and counting: Unraveling trinucleotide repeat diseases Hum. Mol.
Genet. 9: 909-916. (PubMed)
Defective DNA repair and cancer
M. Berneburg and A.R. Lehmann. 2001. Xeroderma pigmentosum and related disorders: Defects in DNA repair and
transcription Adv. Genet. 43: 71-102. (PubMed)
M.W. Lambert and W.C. Lambert. 1999. DNA repair and chromatin structure in genetic diseases Prog. Nucleic Acid
Res. Mol. Biol. 63: 257-310. (PubMed)
C.H. Buys. 2000. Telomeres, telomerase, and cancer N. Engl. J. Med. 342: 1282-1283. (PubMed)
V. Urquidi, D. Tarin, and S. Goodison. 2000. Role of telomerase in cell senescence and oncogenesis Annu. Rev. Med. 51:
65-79. (PubMed)
H.T. Lynch, T.C. Smyrk, P. Watson, S.J. Lanspa, J.F. Lynch, P.M. Lynch, R.J. Cavalieri, and C.R. Boland. 1993.
Genetics, natural history, tumor spectrum, and pathology of hereditary nonpolyposis colorectal cancer: An updated
review Gastroenterology 104: 1535-1549. (PubMed)
R. Fishel, M.K. Lescoe, M.R.S. Rao, N.G. Copeland, N.A. Jenkins, J. Garber, M. Kane, and R. Kolodner. 1993. The
human mutator gene homolog MSH2 and its association with hereditary nonpolyposis colon cancer Cell 75: 1027-1038.
(PubMed)
B.N. Ames and L.S. Gold. 1991. Endogenous mutagens and the causes of aging and cancer Mutat. Res. 250: 3-16.
(PubMed)
B.N. Ames. 1979. Identifying environmental chemicals causing mutations and cancer Science 204: 587-593. (PubMed)
III. Synthesizing the Molecules of Life
28. RNA Synthesis and Splicing
DNA stores genetic information in a stable form that can be readily replicated. However, the expression of this genetic
information requires its flow from DNA to RNA to protein, as was introduced in Chapter 5. The present chapter deals
with how RNA is synthesized and spliced. We begin with transcription in Escherichia coli and focus on three questions:
What are the properties of promoters (the DNA sites at which RNA transcription is initiated), and how do the promoters
function? How do RNA polymerase, the DNA template, and the nascent RNA chain interact with one another? How is
transcription terminated?
We then turn to transcription in eukaryotes, beginning with promoter structure and the transcription-factor proteins that
regulate promoter activity. A distinctive feature of eukaryotic DNA templates is the presence of enhancer sequences that

can stimulate transcriptional initiation more than a thousand base pairs away from the start site. Primary transcripts in
eukaryotes are extensively modified, as exemplified by the capping of the 5
end of an mRNA precursor and the addition
of a long poly(A) tail to its 3
end. Most striking is the splicing of mRNA precursors, which is catalyzed by spliceosomes
consisting of small nuclear ribonucleoprotein particles (snRNPs). The small nuclear RNA (snRNA) molecules in these
complexes play a key role in directing the alignment of splice sites and in mediating catalysis. Indeed, some RNA
molecules can splice themselves in the absence of protein. This landmark discovery by Thomas Cech and Sidney Altman
revealed that RNA molecules can serve as catalysts and greatly influenced our view of molecular evolution.
RNA splicing is not merely a curiosity. Approximately 15% of all genetic diseases are caused by mutations that affect
RNA splicing. Morever, the same pre-mRNA can be spliced differently in various cell types, at different stages of
development, or in response to other biological signals. In addition, individual bases in some pre-mRNA molecules are
changed, in a process called RNA editing. One of the biggest surprises of the sequencing of the human genome was that
only approximately 40,000 genes were identified compared with previous estimates of 100,000 or more. The ability of
one gene to encode more than one distinct mRNA and, hence, more than one protein may play a key role in expanding
the repertoire of our genomes.
28.0.1. An Overview of RNA Synthesis
RNA synthesis, or transcription, is the process of transcribing DNA nucleotide sequence information into RNA
sequence information. RNA synthesis is catalyzed by a large enzyme called RNA polymerase. The basic biochemistry of
RNA synthesis is common to prokaryotes and eukaryotes, although its regulation is more complex in eukaryotes. The
close connection between prokaryotic and eukaryotic transcription has been beautifully illustrated by the recently
determined three-dimensional structures of representative RNA polymerases from prokaryotes and eukaryotes (Figure
28.1). Despite substantial differences in size and number of polypeptide subunits, the overall structures of these enzymes
are quite similar, revealing a common evolutionary origin.
RNA synthesis, like nearly all biological polymerization reactions, takes place in three stages: initiation, elongation, and
termination. RNA polymerase performs multiple functions in this process:
1. It searches DNA for initiation sites, also called promoter sites or simply promoters. For instance, E. coli DNA has
about 2000 promoter sites in its 4.8 × 10
6
bp genome. Because these sequences are on the same molecule of DNA as the
genes being transcribed, they are called cis-acting elements.
2. It unwinds a short stretch of double-helical DNA to produce a single-stranded DNA template from which it takes
instructions.
3. It selects the correct ribonucleoside triphosphate and catalyzes the formation of a phosphodiester bond. This process is
repeated many times as the enzyme moves unidirectionally along the DNA template. RNA polymerase is completely
processive a transcript is synthesized from start to end by a single RNA polymerase molecule.
4. It detects termination signals that specify where a transcript ends.
5. It interacts with activator and repressor proteins that modulate the rate of transcription initiation over a wide dynamic
range. These proteins, which play a more prominent role in eukaryotes than in prokaryotes, are called transcrip-tion
factors or trans-acting elements. Gene expression is controlled mainly at the level of transcription, as will be discussed
in detail in Chapter 31.
The fundamental reaction of RNA synthesis is the formation of a phosphodiester bond. The 3
-hydroxyl group of the last
nucleotide in the chain nucleophilically attacks the α-phosphate group of the incoming nucleoside triphosphate with the
concomitant release of a pyrophosphate (see Figure 5.25). This reaction is thermodynamically favorable, and the
subsequent degradation of the pyrophosphate to orthophosphate locks the reaction in the direction of RNA synthesis.
The chemistry of RNA synthesis is identical for all forms of RNA, including messenger RNA, transfer RNA, and
ribosomal RNA. The basic steps just outlined also apply to all forms. Their synthetic processes differ mainly in
regulation, posttranscriptional processing, and the specific polymerase that participates.
III. Synthesizing the Molecules of Life 28. RNA Synthesis and Splicing
Figure 28.1. RNA Polymerase Structures.
The three-dimensional structures of RNA polymerases from a prokaryote
(Thermus aquaticus) and a eukaryote (Saccharoromyces cerevisiae). The two largest subunits for each structure
are shown in dark red and dark blue. The similarity of these structures reveals that these enzymes have the same
evolutionary origin and have many mechanistic features in common.
III. Synthesizing the Molecules of Life 28. RNA Synthesis and Splicing
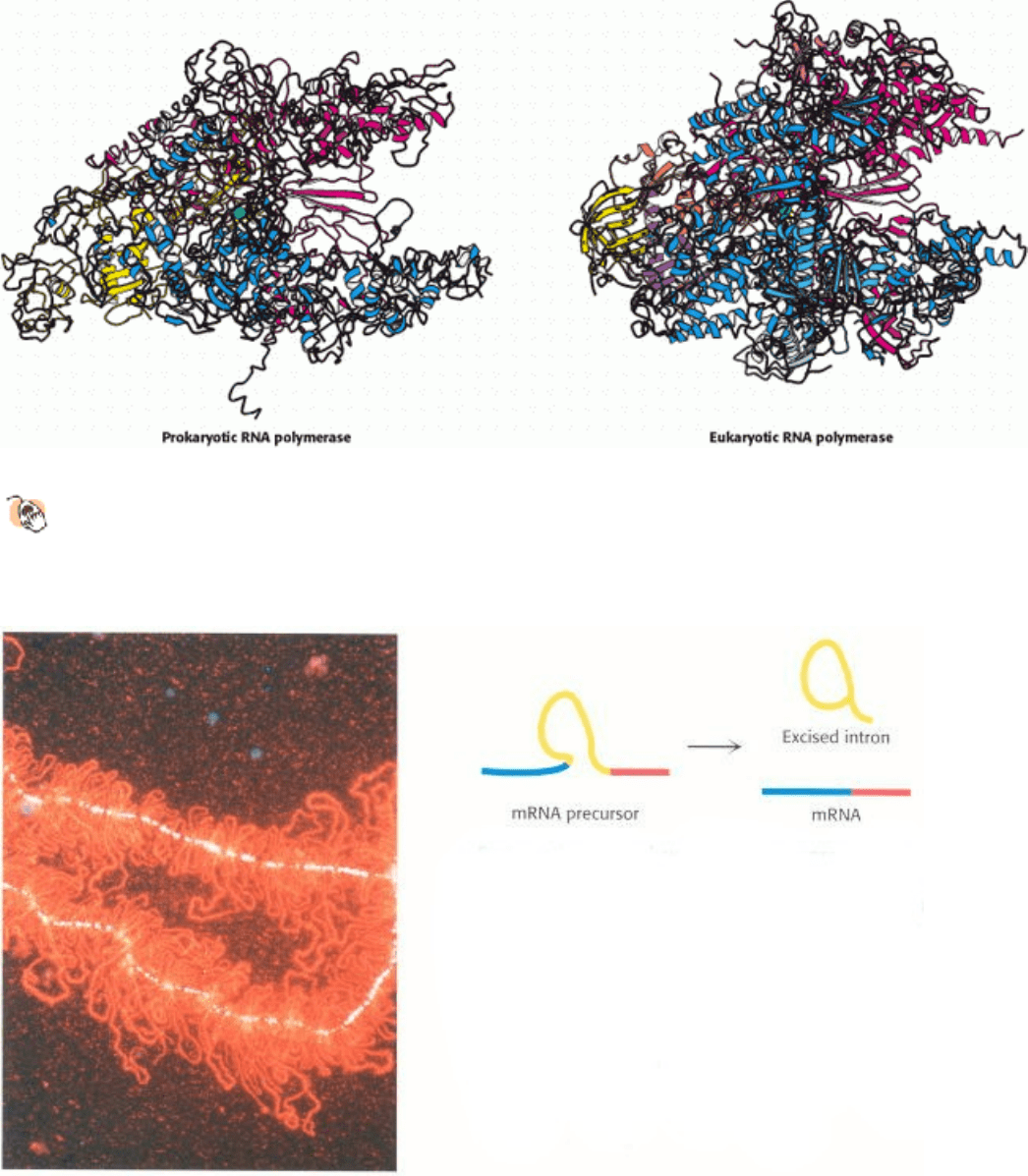
RNA synthesis is a key step in the expression of genetic information. For eukaryotic cells, the initial RNA transcript
(the mRNA precursor) is often spliced, removing introns that do not encode protein sequences. Often, the same pre-
mRNA is spliced differently in different cell types or at different developmental stages. In the image at the left, proteins