Berg J.M., Tymoczko J.L., Stryer L. Biochemistry
Подождите немного. Документ загружается.


associated with RNA splicing (stained with a fluorescent antibody) highlight regions of the newt genome that are being
actively transcribed. [(Left) courtesy of Mark B. Roth and Joseph G. Gall.]
III. Synthesizing the Molecules of Life 28. RNA Synthesis and Splicing
28.1. Transcription Is Catalyzed by RNA Polymerase
We begin our consideration of transcription by examining the process in bacteria such as E. coli. RNA polymerase from
E. coli is a very large (~400 kd) and complex enzyme consisting of four kinds of subunits (Table 28.1). The subunit
composition of the entire enzyme, called the holoenzyme, is α
2
β β σ. The σ subunit helps find a promoter site where
transcription begins, participates in the initiation of RNA synthesis, and then dissociates from the rest of the enzyme.
RNA polymerase without this subunit (α
2
β β ) is called the core enzyme. The core enzyme contains the catalytic site.
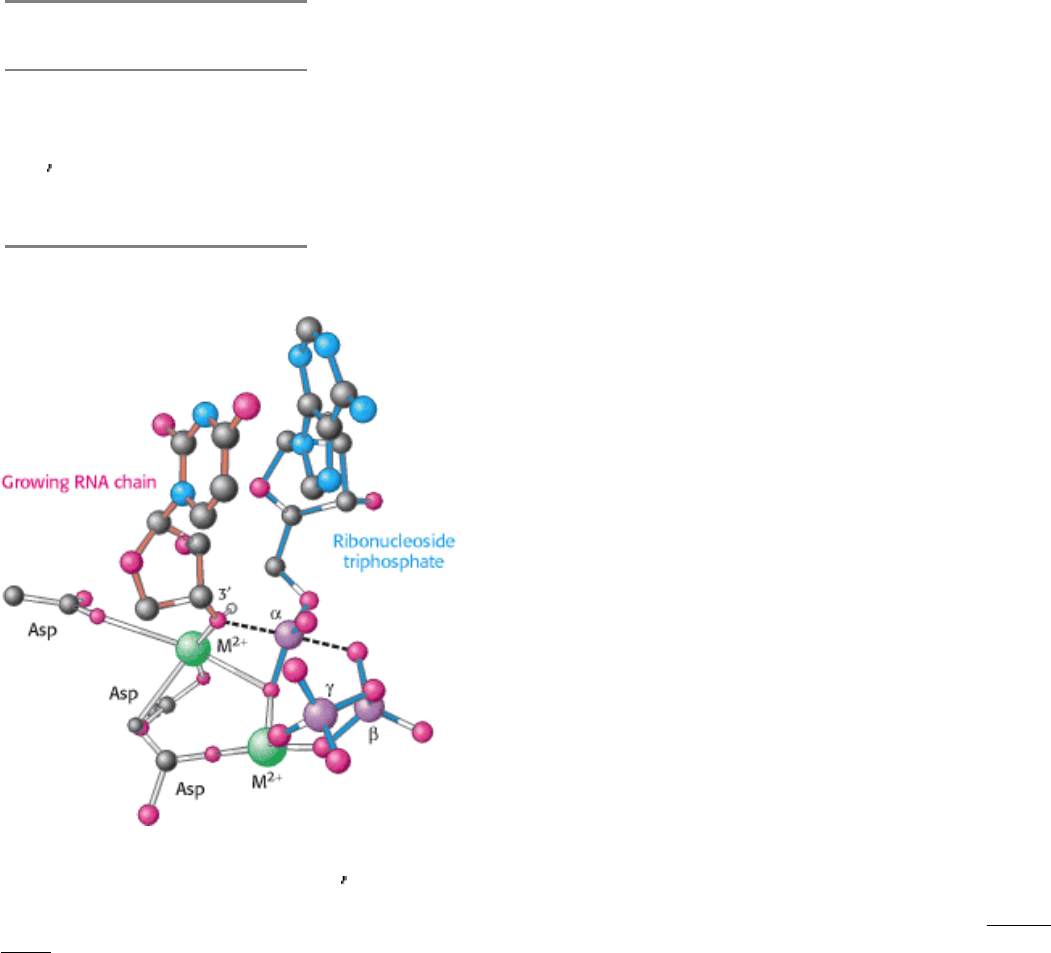
This catalytic site resembles that of DNA polymerase (Secion 27.2.2) in that it includes two metal ions in its active form
(Figure 28.2). One metal ion remains bound to the enzyme, whereas the other appears to come in with the nucleoside
triphosphate and leave with the pyrophosphate. Three conserved aspartate residues of the enzyme participate in binding
these metal ions. Note that the overall structures of DNA polymerase and RNA polymerase are quite different; their
similar active sites are the products of convergent evolution.
28.1.1. Transcription Is Initiated at Promoter Sites on the DNA Template
Transcription starts at promoters on the DNA template. Promoters are sequences of DNA that direct the RNA
polymerase to the proper initiation site for transcription. Promoter sites can be identified and characterized by a
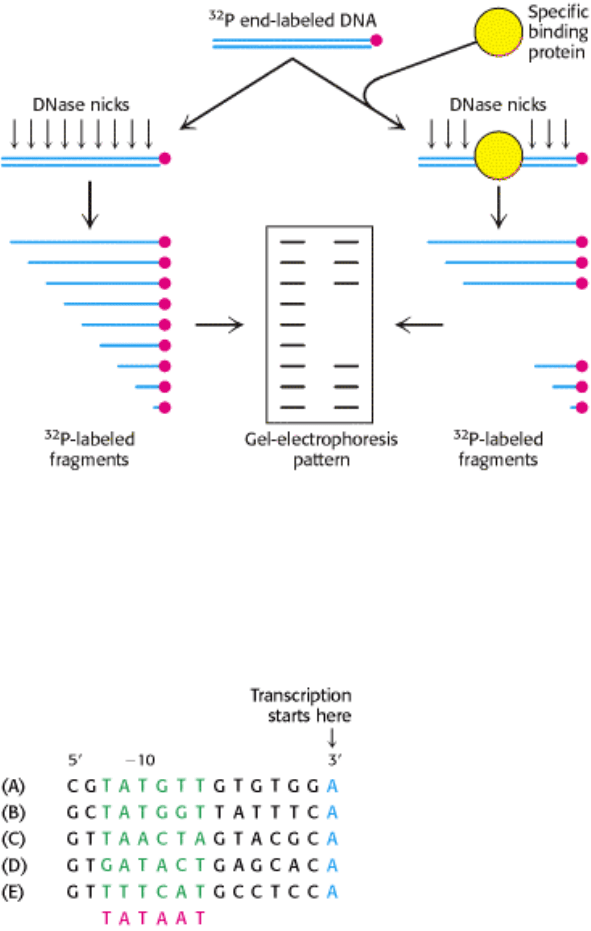
combination of techniques. One powerful technique for characterizing these and other protein-binding sites on DNA is
called footprinting (Figure 28.3). First, one of the strands of a DNA fragment under investigation is labeled on one end
with
32
P. RNA polymerase is added to the labeled DNA, and the complex is digested with DNAse just long enough to
make an average of one cut in each chain. A part of the radioactive DNA is treated in the same way but without the
addition of RNA polymerase to serve as a control. The resulting DNA fragments are separated according to size by
electrophoresis. The gel pattern is highly revealing: a series of bands present in the control sample is absent from the
sample containing RNA polymerase. These bands are missing because RNA polymerase shields DNA from cleavages
that would give rise to the corresponding fragments.
A striking pattern is evident when the sequences of many prokaryotic promoters are compared. Two common motifs are
present on the 5
(upstream) side of the start site. They are known as the -10 sequence and the -35 sequence because
they are centered at about 10 and 35 nucleotides upstream of the start site. These sequences are each 6 bp long. Their
consensus (average) sequences, deduced from analyses of many promoters (Figure 28.4), are
The first nucleotide (the start site) of a transcribed DNA sequence is denoted as +1 and the second one as +2; the
nucleotide preceding the start site is denoted as -1. These designations refer to the coding strand of DNA. Recall that the
sequence of the template strand of DNA is the complement of that of the RNA transcript (see Figure 5.26). In contrast,
the coding strand of DNA has the same sequence as that of the RNA transcript except for thymine (T) in place of uracil
(U). The coding strand is also known as the sense (+) strand, and the template strand as the antisense (-) strand.
Promoters differ markedly in their efficacy. Genes with strong promoters are transcribed frequently
as often as every
2 seconds in E. coli. In contrast, genes with very weak promoters are transcribed about once in 10 minutes. The -10 and -

35 regions of most strong promoters have sequences that correspond closely to the consensus sequences, whereas weak
promoters tend to have multiple substitutions at these sites. Indeed, mutation of a single base in either the -10 sequence
or the -35 sequence can diminish promoter activity. The distance between these conserved sequences also is important; a
separation of 17 nucleotides is optimal. Thus, the efficiency or strength of a promoter sequence serves to regulate
transcription. Regulatory proteins that bind to specific sequences near promoter sites and interact with RNA polymerase
(Chapter 31) also markedly influence the frequency of transcription of many genes.
28.1.2. Sigma Subunits of RNA Polymerase Recognize Promoter Sites
The α
2
β β core of RNA polymerase is unable to start transcription at promoter sites. Rather, the complete α
2
β β σ
holoenzyme is essential for initiation at the correct start site. The σ subunit contributes to specific initiation in two ways.
First, it decreases the affinity of RNA polymerase for general regions of DNA by a factor of 10
4
. In its absence, the core
enzyme binds DNA indiscriminately and tightly. Second, the σ subunit enables RNA polymerase to recognize promoter
sites. A large fragment of a σ subunit was found to have an α helix on its surface; this helix has been implicated in
recognizing the 5
-TATAAT sequence of the -10 region (Figure 28.5). The holoenzyme binds to duplex DNA and moves
along the double helix in search of a promoter, forming transient hydrogen bonds with exposed hydrogen-donor and -
acceptor groups on the base pairs. The search is rapid because RNA polymerase slides along DNA instead of repeatedly
binding and dissociating from it. In other words, the promoter site is encountered by a random walk in one dimension
rather than in three dimensions. The observed rate constant for the binding of RNA polymerase holoenzyme to promoter
sequences is 10
10
M
-1
s
-1
, more than 100 times as fast as could be accomplished by repeated encounters moving on and
off the DNA. The σ subunit is released when the nascent RNA chain reaches nine or ten nucleotides in length. After its
release, it can assist initiation by another core enzyme. Thus, the σ subunit acts catalytically.
E. coli contains multiple σ factors to recognize several types of promoter sequences contained in E. coli DNA. The type
that recognizes the consensus sequences described earlier is called σ
70
because it has a mass of 70 kd. A different σ
factor comes into play when the temperature is raised abruptly. E. coli responds by synthesizing σ
32
, which recognizes
the promoters of heat-shock genes. These promoters exhibit -10 sequences that are somewhat different from the -10
sequence for standard promoters (Figure 28.6). The increased transcription of heat-shock genes leads to the coordinated
synthesis of a series of protective proteins. Other σ factors respond to environmental conditions, such as nitrogen
starvation. These findings demonstrate that σ plays a key role in determining where RNA polymerase initiates
transcription.
28.1.3. RNA Polymerase Must Unwind the Template Double Helix for Transcription to
Take Place
Although RNA polymerase can search for promoter sites when bound to double-helical DNA, a segment of the helix
must be unwound before synthesis can begin. A region of duplex DNA must be unpaired so that nucleotides on one of its
strands become accessible for base-pairing with incoming ribonucleoside triphosphates. The DNA template strand
selects the correct ribonucleoside triphosphate by forming a Watson-Crick base pair with it (Section 5.2.1), as in DNA
synthesis.
How much of the template DNA is unwound by the polymerase? Because unwinding increases the negative supercoiling
of the DNA (Section 27.3.2), this question was answered by analyzing the supercoiling of a circular duplex DNA
exposed to varying amounts of RNA polymerase. Topoisomerase I, an enzyme catalyzing the concerted cleavage and
resealing of duplex DNA (Section 27.3.3), was then added to relax the part of circular DNA not in contact with
polymerase molecules. These DNA samples were analyzed by gel electrophoresis after the removal of bound protein.
The degree of negative supercoiling increased in proportion to the number of RNA polymerase molecules bound per
template DNA, showing that the enzyme unwinds DNA. Each bound polymerase molecule unwinds a 17-bp segment of
DNA, which corresponds to 1.6 turns of B-DNA helix (Figure 28.7).
Negative supercoiling of circular DNA favors the transcription of genes because it facilitates unwinding (Section 27.3.2).
Thus, the introduction of negative supercoils into DNA by topoisomerase II can increase the efficiency of promoters
located at distant sites. However, not all promoter sites are stimulated by negative supercoiling. The promoter site for
topoisomerase II itself is a noteworthy exception. Negative supercoiling decreases the rate of transcription of this gene,
an elegant feedback control ensuring that DNA does not become excessively supercoiled. Negative supercoiling could
decrease the efficiency of this promoter by changing the structural relation of the -10 and -35 regions.
The transition from the closed promoter complex (in which DNA is double helical) to the open promoter complex (in
which a DNA segment is unwound) is an essential event in transcription. The stage is now set for the formation of the
first phosphodiester bond of the new RNA chain.
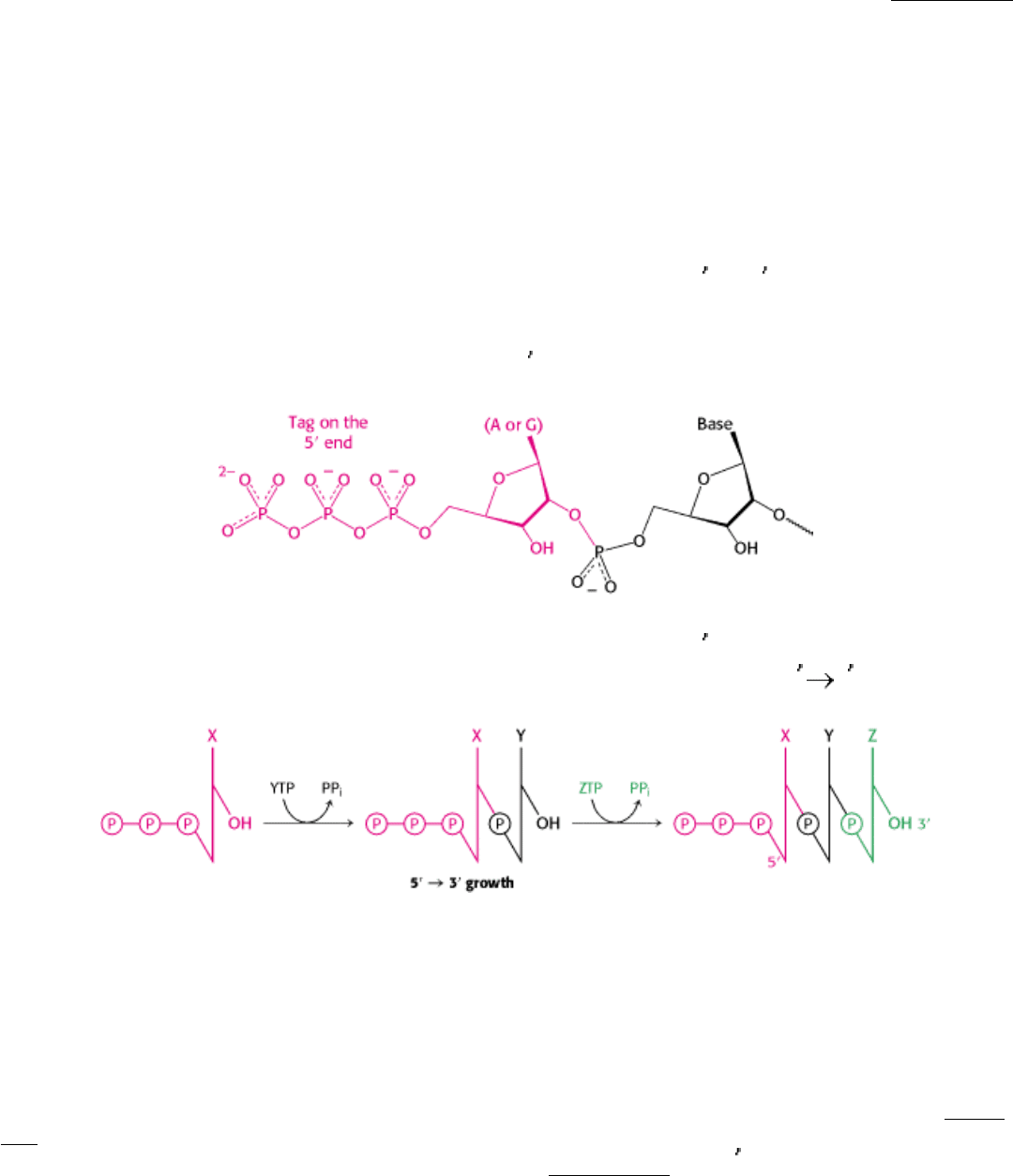
28.1.4. RNA Chains Are Formed de Novo and Grow in the 5-to-3 Direction
In contrast with DNA synthesis, RNA synthesis can start de novo, without the requirement for a primer. Most newly
synthesized RNA chains carry a highly distinctive tag on the 5
end: the first base at that end is either pppG or pppA.
The presence of the triphosphate moiety suggests that RNA synthesis starts at the 5
end. The results of labeling
experiments with γ-
32
P substrates confirmed that RNA chains, like DNA chains, grow in the 5 3 direction.
28.1.5. Elongation Takes Place at Transcription Bubbles That Move Along the DNA
Template
The elongation phase of RNA synthesis begins after the formation of the first phosphodiester bond. An important change
is the loss of σ; recall that the core enzyme without σ binds more strongly to the DNA template. Indeed, RNA
polymerase stays bound to its template until a termination signal is reached. The region containing RNA polymerase,
DNA, and nascent RNA is called a transcription bubble because it contains a locally melted "bubble" of DNA (Figure
28.8). The newly synthesized RNA forms a hybrid helix with the template DNA strand. This RNA-DNA helix is about 8
bp long, which corresponds to nearly one turn of a double helix (Section 27.1.3). The 3 -hydroxyl group of the RNA in
this hybrid helix is positioned so that it can attack the α-phosphorus atom of an incoming ribonucleoside triphosphate.
The core enzyme also contains a binding site for the other DNA strand. About 17 bp of DNA are unwound throughout
the elongation phase, as in the initiation phase. The transcription bubble moves a distance of 170 Å (17 nm) in a second,
which corresponds to a rate of elongation of about 50 nucleotides per second. Although rapid, it is much slower than the
rate of DNA synthesis, which is 800 nucleotides per second.

The lengths of the RNA-DNA hybrid and of the unwound region of DNA stay rather constant as RNA polymerase
moves along the DNA template. This finding indicates that DNA is rewound at about the same rate at the rear of RNA
polymerase as it is unwound at the front of the enzyme. The RNA-DNA hybrid must also rotate each time a nucleotide is
added so that the 3
-OH end of the RNA stays at the catalytic site. The length of the RNA-DNA hybrid is determined by
a structure within the enzyme that forces the RNA-DNA hybrid to separate, allowing the RNA chain to exit from the
enzyme and the DNA chain to rejoin its DNA partner (Figure 28.9).
It is noteworthy that RNA polymerase lacks nuclease activity. Thus, in contrast with DNA polymerase, RNA polymerase
does not correct the nascent polynucleotide chain. Consequently, the fidelity of transcription is much lower than that of
replication. The error rate of RNA synthesis is of the order of one mistake per 10
4
or 10
5
nucleotides, about 10
5
times as
high as that of DNA synthesis. The much lower fidelity of RNA synthesis can be tolerated because mistakes are not
transmitted to progeny. For most genes, many RNA transcripts are synthesized; a few defective transcripts are unlikely
to be harmful.
28.1.6. An RNA Hairpin Followed by Several Uracil Residues Terminates the
Transcription of Some Genes
The termination of transcription is as precisely controlled as its initiation. In the termination phase of transcription, the
formation of phosphodiester bonds ceases, the RNA-DNA hybrid dissociates, the melted region of DNA rewinds, and
RNA polymerase releases the DNA. What determines where transcription is terminated? The transcribed regions of
DNA templates contain stop signals. The simplest one is a palindromic GC-rich region followed by an AT-rich region.
The RNA transcript of this DNA palindrome is self-complementary (Figure 28.10). Hence, its bases can pair to form a
hairpin structure with a stem and loop, a structure favored by its high content of G and C residues. Guanine-cytosine
base pairs are more stable than adenine-thymine pairs because of the extra hydrogen bond in the base pair. This stable
hairpin is followed by a sequence of four or more uracil residues, which also are crucial for termination. The RNA
transcript ends within or just after them.
How does this combination hairpin-oligo(U) structure terminate transcription? First, it seems likely that RNA
polymerase pauses immediately after it has synthesized a stretch of RNA that folds into a hairpin. Furthermore, the RNA-
DNA hybrid helix produced after the hairpin is unstable because its rU-dA base pairs are the weakest of the four kinds.
Hence, the pause in transcription caused by the hairpin permits the weakly bound na-scent RNA to dissociate from the
DNA template and then from the enzyme. The solitary DNA template strand rejoins its partner to re-form the DNA
duplex, and the transcription bubble closes.
28.1.7. The Rho Protein Helps Terminate the Transcription of Some Genes
RNA polymerase needs no help to terminate transcription at a hairpin followed by several U residues. At other sites,
however, termination requires the participation of an additional factor. This discovery was prompted by the observation
that some RNA molecules synthesized in vitro by RNA polymerase acting alone are longer than those made in vivo. The
missing factor, a protein that caused the correct termination, was isolated and named rho (ρ). Additional information
about the action of ρ was obtained by adding this termination factor to an incubation mixture at various times after the
initiation of RNA synthesis (Figure 28.11). RNAs with sedimentation coefficients of 10S, 13S, and 17S were obtained
when ρ was added at initiation, a few seconds after initiation, and 2 minutes after initiation, respectively. If no ρ was
added, transcription yielded a 23S RNA product. It is evident that the template contains at least three termination sites
that respond to ρ (yielding 10S, 13S, and 17S RNA) and one termination site that does not (yielding 23S RNA). Thus,
specific termination at a site producing 23S RNA can occur in the absence of ρ. However, ρ detects additional
termination signals that are not recognized by RNA polymerase alone.
How does ρ provoke the termination of RNA synthesis? A key clue is the finding that ρ hydrolyzes ATP in the presence
of single-stranded RNA but not in the presence of DNA or duplex RNA. Hexameric ρ, which is structurally similar to and
homologous to ATP synthase (Section 18.4.1), specifically binds single-stranded RNA; a stretch of 72 nucleotides is
bound in such a way that the RNA passes through the center of the structure (Figure 28.12). Rho is brought into action
by sequences located in the nascent RNA that are rich in cytosine and poor in guanine. The ATPase activity of ρ enables
the protein to pull the nascent RNA while pursuing RNA polymerase. When ρ catches RNA polymerase at the
transcription bubble, it breaks the RNA-DNA hybrid helix by functioning as an RNA-DNA helicase. Given the structural
and evolutionary connection, it is possible that the mechanism of action of ρ is similar to that of ATP synthase, with the
single-stranded RNA playing the role of the γ subunit.
Proteins in addition to ρ mediate and modulate termination. For example, the nusA protein enables RNA polymerase in
E. coli to recognize a characteristic class of termination sites. In E. coli, specialized termination signals called
attenuators are regulated to meet the nutritional needs of the cell (Section 31.4.1). A common feature of protein-
independent and proteindependent termination is that the functioning signals lie in newly synthesized RNA rather than in
the DNA template.
28.1.8. Precursors of Transfer and Ribosomal RNA Are Cleaved and Chemically
Modified After Transcription
In prokaryotes, messenger RNA molecules undergo little or no modification after synthesis by RNA polymerase. Indeed,
many mRNA molecules are translated while they are being transcribed. In contrast, transfer RNA and ribosomal RNA
molecules are generated by cleavage and other modifications of nascent RNA chains. For example, in E. coli, three kinds
of rRNA molecules and a tRNA molecule are excised from a single primary RNA transcript that also contains spacer
regions (Figure 28.13). Other transcripts contain arrays of several kinds of tRNA or of several copies of the same tRNA.
The nucleases that cleave and trim these precursors of rRNA and tRNA are highly precise. Ribonuclease P, for example,
generates the correct 5
terminus of all tRNA molecules in E. coli. This interesting enzyme contains a catalytically active
RNA molecule. Ribonuclease III excises 5S, 16S, and 23S rRNA precursors from the primary transcript by cleaving
double-helical hairpin regions at specific sites.
A second type of processing is the addition of nucleotides to the termini of some RNA chains. For example, CCA, a
terminal sequence required for the function of all tRNAs, is added to the 3
ends of tRNA molecules that do not already
possess this terminal sequence. A third type of processing is the modification of bases and ribose units of ribosomal
RNAs. In prokaryotes, some bases of rRNA are methylated. Unusual bases are found in all tRNA molecules (Section
29.1.2). They are formed by the enzymatic modification of a standard ribonucleotide in a tRNA precursor. For example,
uridylate residues are modified after transcription to form ribothymidylate and pseudouridylate. These modifications
generate diversity, allowing greater structural and functional versatility.
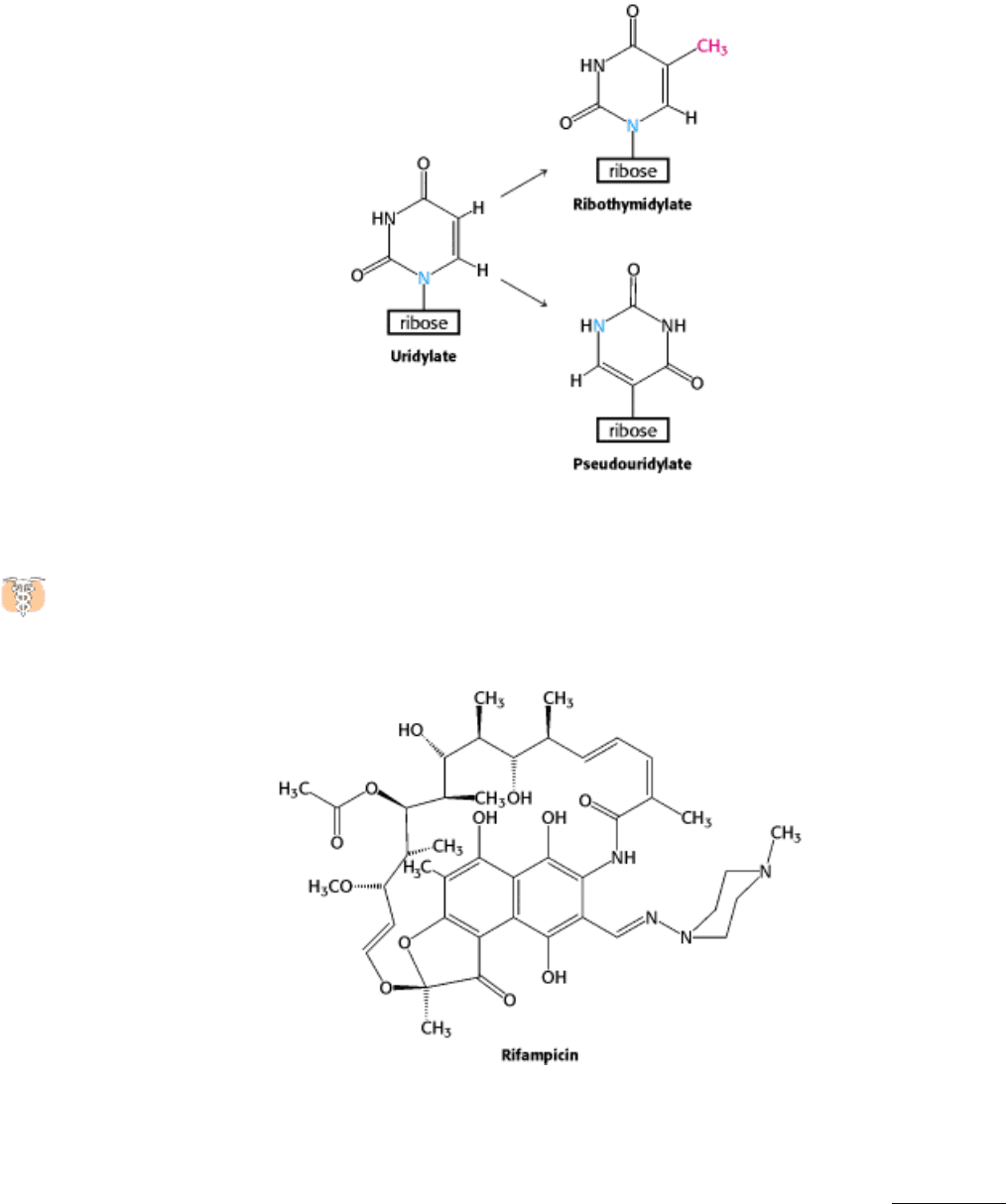
28.1.9. Antibiotic Inhibitors of Transcription
Many antibiotics are highly specific inhibitors of biological processes. Rifampicin and actinomycin are two
antibiotics that inhibit transcription, although in quite different ways. Rifampicin is a semisynthetic derivative of
rifamycins, which are derived from a strain of Streptomyces.
This antibiotic specifically inhibits the initiation of RNA synthesis. Rifampicin does not block the binding of RNA
polymerase to the DNA template; rather, it interferes with the formation of the first few phosphodiester bonds in the
RNA chain. The structure of a complex between a prokaryotic RNA polymerase and rifampicin reveals that the
antibiotic blocks the channel into which the RNA-DNA hybrid generated by the enzyme must pass (Figure 28.14). The
binding site is 12 Å from the active site itself. Rifampicin does not hinder chain elongation once initiated, because the
RNA-DNA hybrid present in the enzyme prevents the antibiotic from binding.
Actinomycin D, a polypeptide-containing antibiotic from a different strain of Streptomyces, inhibits transcription by an
entirely different mechanism. Actinomycin D binds tightly and specifically to double-helical DNA and thereby prevents it
from being an effective template for RNA synthesis. It does not bind to single-stranded DNA or RNA, double-stranded
RNA, or RNA-DNA hybrids. The results of spectroscopic and hydrodynamic studies of complexes of actinomycin D and
DNA suggested that the phenoxazone ring of actinomycin slips in between neighboring base pairs in DNA. This mode of
binding is called intercalation. At low concentrations, actinomycin D inhibits transcription without significantly
affecting DNA replication or protein synthesis. Hence, actinomycin D is extensively used as a highly specific inhibitor of
the formation of new RNA in both prokaryotic and eukaryotic cells. Its ability to inhibit the growth of rapidly dividing
cells makes it an effective therapeutic agent in the treatment of some cancers.
III. Synthesizing the Molecules of Life 28. RNA Synthesis and Splicing 28.1. Transcription Is Catalyzed by RNA Polymerase
Table 28.1. Subunits of RNA polymerase from E. coli
Subunit Gene Number Mass(kd)
α
rpoA 2 37
β
rpoB 1 151
β
rpoC 1 155
σ
70
rpoD 1 70
III. Synthesizing the Molecules of Life 28. RNA Synthesis and Splicing 28.1. Transcription Is Catalyzed by RNA Polymerase
Figure 28.2. RNA Polymerase Active Site. A model of the transition state for phosphodiester-bond formation in the
active site of RNA polymerase. The 3
-hydroxyl group of the growing RNA chain attacks the α -phosphate of the
incoming nucleoside triphosphate. This transition state is structurally similar to that in DNA polymerase (see Figure
27.12).
III. Synthesizing the Molecules of Life 28. RNA Synthesis and Splicing 28.1. Transcription Is Catalyzed by RNA Polymerase
Figure 28.3. Footprinting. One end of a DNA chain is labeled with
32
P (shown as a red circle). This labeled DNA is
then treated with DNAse I such that each fragment is cut only once. The same cleavage is carried out after a protein that
binds to specific sites on the DNA has been added. The bound protein protects a segment on the DNA from the action of
DNAse I. Hence, certain fragments present in the reaction without protein will be missing. These missing bands in the
gel pattern identify the binding site on DNA.
III. Synthesizing the Molecules of Life 28. RNA Synthesis and Splicing 28.1. Transcription Is Catalyzed by RNA Polymerase
Figure 28.4. Prokaryotic Promoter Sequences. A comparison of five sequences from prokaryotic promoters reveals a
recurring sequence of TATAAT centered on position -10. The -10 consensus sequence (in red) was deduced from a large
number of promoter sequences. The sequences are from the (A) lac, (B) gal, and (C) trp operons of E. coli; (D) from λ
phage; and (E) from Φ Χ174 phage.
III. Synthesizing the Molecules of Life 28. RNA Synthesis and Splicing 28.1. Transcription Is Catalyzed by RNA Polymerase
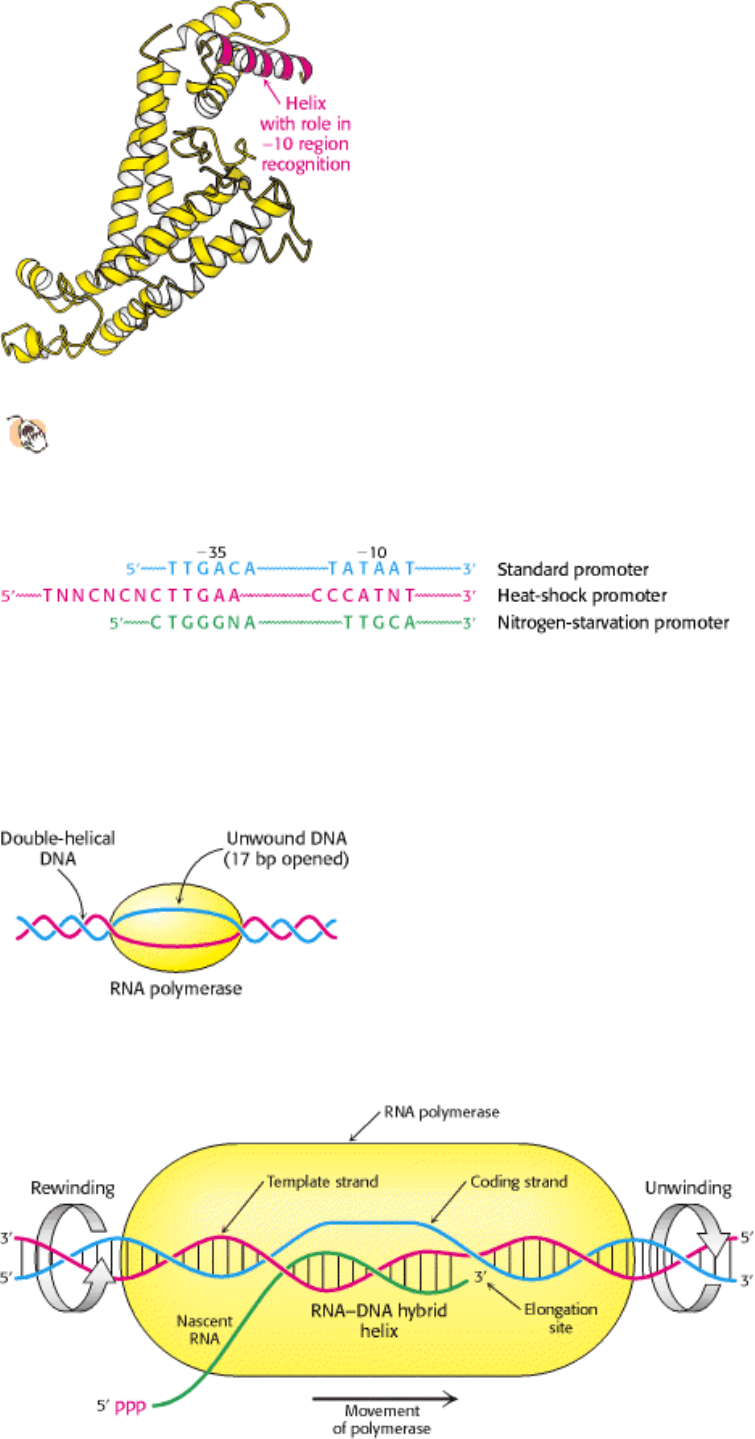
Figure 28.5. Structure of the σ Subunit. The structure of a fragment from the E. coli subunit σ
70
reveals the position
of an α helix on the protein surface; this helix plays an important role in binding to the -10 TATAAT sequence.
III. Synthesizing the Molecules of Life 28. RNA Synthesis and Splicing 28.1. Transcription Is Catalyzed by RNA Polymerase
Figure 28.6. Alternative Promoter Sequences. A comparison of the consensus sequences of standard promoters, heat-
shock promoters, and nitrogen-starvation promoters of E. coli. These promoters are recognized by σ
70
, σ
32
, and σ
54
,
respectively.
III. Synthesizing the Molecules of Life 28. RNA Synthesis and Splicing 28.1. Transcription Is Catalyzed by RNA Polymerase
Figure 28.7. DNA Unwinding. RNA polymerase unwinds about 17 base pairs of template DNA.
III. Synthesizing the Molecules of Life 28. RNA Synthesis and Splicing 28.1. Transcription Is Catalyzed by RNA Polymerase
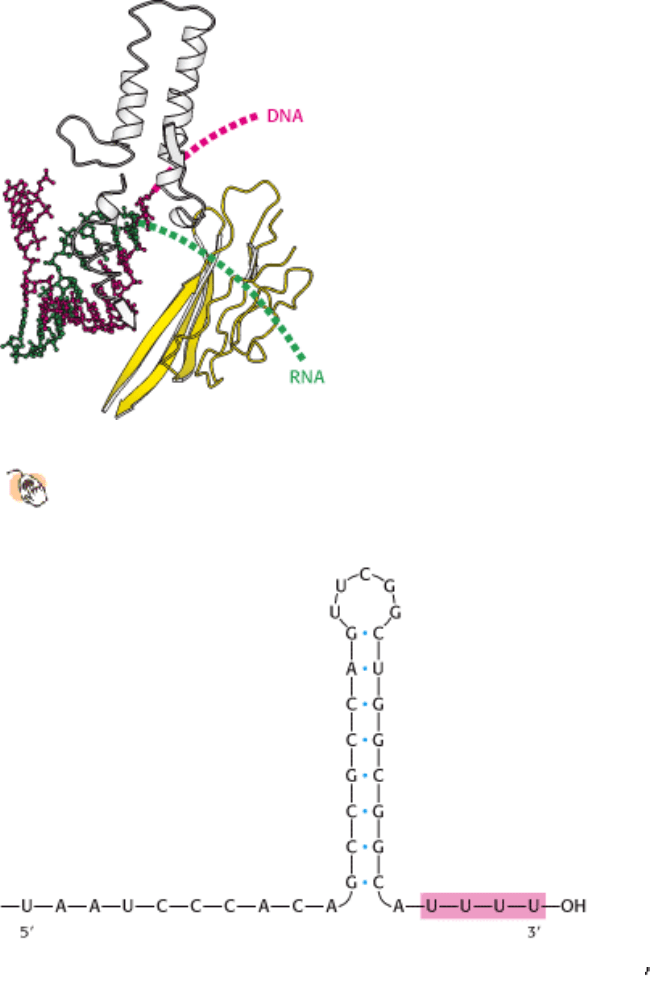
Figure 28.8. Transcription Bubble. A schematic representation of a transcription bubble in the elongation of an RNA
transcript. Duplex DNA is unwound at the forward end of RNA polymerase and rewound at its rear end. The RNA-DNA
hybrid rotates during elongation.
III. Synthesizing the Molecules of Life 28. RNA Synthesis and Splicing 28.1. Transcription Is Catalyzed by RNA Polymerase
Figure 28.9. RNA-DNA Hybrid Separation.
A structure within RNA polymerase forces the separation of the RNA-
DNA hybrid, allowing the DNA strand to exit in one direction and the RNA product to exit in another.
III. Synthesizing the Molecules of Life 28. RNA Synthesis and Splicing 28.1. Transcription Is Catalyzed by RNA Polymerase
Figure 28.10. Termination Signal. A termination signal found at the 3
end of an mRNA transcript consists of a series
of bases that form a stable stem-loop structure and a series of U residues.