Проничев И.В. Лекции по физиологии центральной нервной системы
Подождите немного. Документ загружается.

В процессе развития ПД действуют многие факторы , связанные прямыми и обратными
связями.
Например, на восходящей фазе ПД действует система факторов с положительной
обратной связью:
Здесь деполяризация увеличивает Р
Nа
, а последняя порождает I
Na
, усиливающий
деполяризацию (пока и поскольку INa больше I
к
+I утечки). В силу этих отношений I
Na
в
ПД и восходящая фаза самого ПД развивается быстро, с ускорением, а амплитуда ПД
всегда достигает некоторого максимума, более или менее приближенного к E
Na
(в
зависимости от степени компенсации натриевого тока токами К
+
и утечки). На спаде ПД
при реполяризации для INa действует та же система факторов, но с обратными знаками
эффектов. А для калиевого тока действует система факторов с отрицательной обратной
связью:
В итоге спад I
к
несколько затягивается.
Если скрупулезно учесть все эти факторы и их взаимодействие, то можно из величин
токов, получаемых при фиксации мембранного потенциала на разных уровнях, рассчитать
форму нормального ПД, что и сделали А. Хаксли и А. Ходжкин. Было показано
совпадение расчетных параметров ПД и параметров ПД, регистрируемых в опыте.
Расход ионов на пик одного проводящегося ПД даже в гигантском аксоне кальмара очень
мал. Например, расход внутриаксонального К при этом приблизительно равен 1-й
миллионной доле внутреннего калиевого запаса. Таким образом, пик ПД - весьма
экономичный сигнал, практически не нарушающий ионных градиентов на мембране,
энергией которых он питается. Ионные градиенты на мембране - это "пружина", энергии
которой может хватить на 5 • 10
5
ПД без подзарядки. Но для длительной работы волокна
ионные градиенты нужно восстанавливать, что и обеспечивает работа натрий-калиевого
насоса мембраны.
Несколько слов о механизме подпорогового локального ответа (ЛО) в нервном волокне.
Этот ответ имеет в общем тот же механизм, что и ПД. Его восходящая фаза определяется
входящим током Na
+
, а нисходящая - выходящим калиевым током. Но амплитуда ЛО (и
его натриевый ток Na) пропорциональна силе подпорогового раздражения, а не
стандартна, как у ПД, т. е. не подчиняется правилу "все или ничего". ЛО не вызывает
абсолютной рефрактерности, хотя обнаруживает относительную рефрактерность из-за
инактивации части натриевых каналов.
Необходимо остановиться на некоторых особенностях потенциалов действия в телах
нейронов моллюсков.
Тела гигантских нейронов брюхоногих моллюсков - это крупные образования, не
имеющие на себе синапсов. У этих нейронов синапсы находятся на так называемом
центральном отростке.
Большие размеры и однородность поверхности позволяют исследовать тела этих нейронов
с применением самых точных современных электрофизиологических методов. Так, с
помощью двух или большего числа микроэлектродов, вводимых в эти клетки, производят
электрическое раздражение клетки, регистрируют ее МПП и ПД, исследуют R
м
, С
τ
;
производят фиксацию мембранного потенциала и на этом фоне изучают трансмембранные
токи, лежащие в основе возбуждения.
В лаборатории П. Г. Костюка разработан метод внутриклеточного диализа одиночной
гигантской нервной клетки. Он состоит в следующем: механически изолированную
гигантскую нервную клетку помещают в соответствующем солевом растворе в камеру с
плоскими стенками, на дне которой имеется коническая пора; ее широкая часть
(обращенная в этот сосуд) может пропустить клетку, а узкая нет. Стенки поры покрыты
клейкой массой. Под камерой, содержащей клетку, находится другая аналогичная камера
(с тем же солевым раствором), имеющая сообщение с первой только через упомянутую
коническую пору. Клетку подводят к поре и "засасывают" в нее, понижая
гидростатическое давление в нижнем сосуде. При этом части клетки, вошедшие в пору,
прилипают к ее стенкам. Часть клетки, выступающая в нижнюю камеру, разрушается
пропусканием через нижнюю камеру безкальциевого раствора. После этого
гидростатическое давление в нижней камере и среду в ней возвращают к норме. В итоге
получается препарат соматической мембраны, омываемой снаружи жидкостью верхней
камеры, а изнутри жидкостью нижней камеры. Внутриклеточные структуры при этом
вымываются. На этом препарате соматической мембраны при его электрическом
раздражении, как показали опыты, могут быть получены полноценные ПД. Удалось
осуществить кламп потенциала такой мембраны, изучить трансмембранные токи при ее
активации, а при уменьшении площади препарата даже получить характеристики токов
отдельных мембранных каналов.
МПП сомы невелик (-40------50 мВ) и имеет механизм, в общем сходный с известным для
аксонов. ПД имеет овершут и достигает 100 и более милливольт (мВ). По форме он схож с
аксонным. Но ионные механизмы ПД мембраны сомы нейрона моллюска имеют
существенные отличия от известных для нервных волокон. Главное отличие состоит в
том, что ПД здесь генерируется не только за счет входящего натриевого тока (как в
аксоне), но и за счет входящего кальциевого тока. В соматической мембране имеются
сходные с натриевыми, но независимые от них специальные кальциевые каналы,
открываемые деполяризацией. Ток Са
2+
через них осуществляется по электрохимическому
градиенту. Эти каналы не блокируются тетро-дотоксином (ТТХ), но блокируются
кобальтом, верапамилом и препаратом Д-600. Кальциевый ток развивается медленнее
натриевого и гораздо медленнее инактивируется (много секунд!).
В соматической мембране нейрона моллюсков обнаружены, кроме того, особые
"быстрые" потенциалозависимые калиевые каналы (очень быстро активирующиеся и
инактивирующиеся). Эти каналы почти не участвуют в формировании МПП и ПД, ибо
они уже инактивированы при МП = 70 мВ, т. е. при МПП. Но эти каналы принимают
участие в регулировке МП на фоне следовой гиперполяризации, тем самым определяя в
какой-то мере частоту ритмического разряда в ответ на то или иное воздействие.
Калиевые каналы, участвующие в ПД, т. е. каналы "задержанного" выходящего тока, здесь
также имеют особенность - они инактивируются быстрее, чем в аксоне. За счет этого у
соматических ПД, следующих ритмически, прогрессивно удлиняются нисходящие фазы,
но удерживается величина амплитуды, ибо ослабление калиевого шунта компенсирует
ослабление натриевых токов от натриевой инактивации. Особенности ионных каналов
соматической мембраны, вероятно, имеют значение не только для электрогенеза. Можно
думать, что они существенны и для обеспечения интенсивно протекающих в соме
метаболических реакций, чувствительных к [Са
2+
], [К
+
]и [Na
+
] внутри клетки. Однако у
разных животных эти характеристики перикарионов не одинаковы. В телах нейронов
спинномозговых ганглиев крыс, исследованных по такой же методике, не обнаружилось
кальциевых каналов, участвующих в генерации ПД, вместо них там были найдены особые
"медленные" натриевые каналы.
Следует сказать несколько слов о ПД перехватов Ранвье нервных волокон позвоночных
животных. Потенциалы действия перехватов Ранвье имеют в общем такой же механизм,
как н у гигантского аксона кальмара. Однако плотность натриевого тока ПД здесь гораздо
выше. Нисходящая фаза ПД определяется в перехвате не только выходящим калиевым
током, но в значительной мере неспецифическим током утечки. Роль этого тока особенно
велика у птиц и млекопитающих. По последним данным у миелинизированных нервных
волокон млекопитающих в мембране перехватов очень мало или даже совсем нет
потенциалозависимых калиевых каналов. Но у этих волокон такие каналы имеются в
межперехватной (интернодальной) мембране. По-видимому, у всех позвоночных
неспецифический ток утечки, определяющий нисходящую фазу ПД в перехватах
миелинизированных волокон, протекает отчасти и через насечки миелиновой оболочки. В
генезе следового отрицательного потенциала неизолированного перехвата Ранвье очень
большую, если не основную роль играет накопление ионов К
+
в щели перехвата.
В заключение остановимся на так называемых шумах ионных каналов. Каждый из ионных
каналов мембраны, обладающих теми или иными воротными механизмами, даже при
постоянстве МП и химического состава среды не находится все время в каком-то одном
положении; он то открывается, то закрывается. Эти переходы от закрытого состояния к
открытому и обратно осуществляются по закону случая и практически мгновенно, что
обусловливает прямоугольную форму одиночного толчка тока в канале. Время открытого
состояния называют "временем жизни канала". Хаотическое открытие и закрытие каналов,
порождающее перемещения ионов, создает электрический шум. Для
потенциалозависимых каналов соответствующее электрическое поле является фактором
резко увеличивающим вероятность открытого состояния каналов, что и создает эффект
увеличения ионной проницаемости. Для исследования динамики отдельных каналов
определенного вида исследуют слабый электрический шум, создаваемый этими каналами
в каком-то небольшом участке мембраны в условиях клампа МП и химической блокады
прочих каналов. Например, в мембране гигантского аксона кальмара так изучили шумы
отдельно натриевых и калиевых каналов.
Анализ этих шумов позволил рассчитать плотность расположения каналов в мембране и
проводимость одиночного канала.
В гигантском аксоне кальмара плотность натриевых каналов оказалась равной 300/1 мкм
2
,
проводимость канала 4 рмО. В перехвате Ранвье амфибий плотность натриевых каналов -
2000/1 мкм
2
, а средняя проводимость канала равна 8 рмО.

Лекция 4. Распространение электрона и
проведение потенциала действия
Мы уже говорили о том, что если мембрана нервного волокна (нервной клетки или
мышечного волокна) в какой-то точке подвергается действию входящего или выходящего
постоянного электрического тока, то в этой точке МП оказывается постоянно измененным
- сниженным при выходящем токе и повышенным при входящем токе. Это состояние
измененного МП называют физическим электротоном (тонус, т. е. постоянное
напряжение).
Физический электротон даже при строго локальной подаче тока с помощью
внутриклеточного микроэлектрода на какую-либо точку мембраны не локализуется в этой
точке мембраны, расползается во все стороны от нее, ослабевая (декрементируя). Это
распространение физического электротона по мембране, хотя само по себе и не
обеспечивает дальней связи, тем не менее имеет очень большое физиологическое
значение. Оно обеспечивает ближнюю связь по мембране в объектах с малой
протяженностью (например, в телах нейронов, в некоторых мышечных волокнах, в
клетках глии, эпителия и др.). Это же распространение электротона является одной из
основ механизма бездекрементного проведения возбуждения (потенциала действия) в
протяженных нервных и мышечных волокнах.
Рассмотрим некоторые подробности распространения электротона и причины этого
распространения.
При включении, достаточно длительном действии, а затем выключении тока электротон
соответственно возникает (нарастает, расползается), далее существует как стационарное
распределение потенциалов вокруг электрода и, наконец, исчезает. Причина
распространения электротона заключена в кабельной структуре клеточных объектов,
например нервного (мышечного) волокна. Нервное волокно подобно кабелю имеет
хорошо проводящий электрический ток стержень - аксоплазму, и плохо проводящую
высокоомную оболочку - мембрану. При такой структуре электрический ток, вводимый
микроэлектродом в какую-либо точку кабеля (при наружном расположении другого
электрода) как бы стремится использовать проводящий стержень для подхода к возможно
большему числу точек высокоомной мембраны, сопротивление которой ему необходимо
преодолеть. Очевидно, если сопротивление аксоплазмы (R
i
) и сопротивление среды (R
e
) по
сравнению с сопротивлением мембраны (R
м
) пренебрежимо малы, то подаваемый ток
сможет использовать все точки мембраны волокна практически равномерно и элек-тротон
равномерно охватит все волокно. Наоборот, при сопротивлении мембраны (R
M
)
пренебрежимо малом по сравнению с сопротивлением аксоплазмы (R
i
) и среды (R
e
),
электротон локализовался бы практически в области микроэлектрода. При соизмеримости
R
M
и R
e
, стационарный электротон экспоненциально затухает в пространстве с константой
где R
M
, R
i
и R
e
рассчитаны на единицу длины (1 см).
Так как R
i
зависит от диаметра волокна (D), то при использовании тех же показателей,
рассчитанных на единицу площади (1 см
2
), имеем:
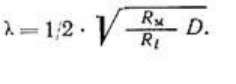
Постепенное, экспоненциальное развитие и исчезновение электротона во времени (в
каждой точке мембраны) объясняется наличием мембранной емкости С
м
. Скорость этого
развития определяется постоянной времени мембраны τ
м
. Чем больше τ, тем медленнее
возникает и исчезает электроток. Вдали от точки приложения электрода электроток
возникает и исчезает медленнее. Поскольку электроток при включении тока как бы
расползается по мембране, можно говорить о скорости распространения электротона V
эт
.
Обратимся теперь к феномену проведения возбуждения, т. е. распространения ПД,
имеющему место в нервном или фазном мышечном волокне, а также в соме и дендритах
нейрона.
Возбуждение (ПД) как и электротон, распространяется по мембране клеток всесторонне (в
волокнах, грубо говоря, "двухсторонне"). Но проведение возбуждения в отличие от
распространения электротона происходит (в однородном проводнике!) без снижения
амплитуды ПД и без снижения скорости (бездекрементно). Механизм проведения
возбуждения, как хорошо теперь известно, включает в себя два компонента: 1)
раздражающее действие катэлектротонического сигнала, порождаемого локальным ПД, на
соседний участок электровозбудимой мембраны и 2) возникновение ПД в этом соседнем
раздражаемом участке мембраны. Проведение ПД - это нечто вроде эстафеты, в которой
каждый участок вдоль волокна выступает как раздражаемый, а затем как раздражающий
последующий участок.
То, что ПД распространяется с помощью петли электротона, доказывается фактом
перескока нервного импульса через узкий участок инактивированной мембраны
(отравленной или охлажденной) . Хотя сам по себе этот участок не генерирует ПД, но ПД,
возникающий в доблоковой области, вызывает за участком блока катэлектротонический
сдвиг потенциала, на вершине которого можно определить ЛО. Иногда здесь получается и
ПД В последнем случае возбуждение как бы "перескакивает" через инактивированный
участок и распространяется дальше. Перескоку возбуждения через такой участок
отчетливо способствует снижение сопротивления внешней среды (R
e
), в частности,
шунтировка нервного кабеля с помощью металлических стерженьков и ртути,
применявшаяся Ходжкиным в опытах на гигантском аксоне кальмара (1939). Снижение R
e
делает электроток более распространенным в пространстве что и способствует перескоку
ПД.
Назовем реальные скорости в некоторых немиелинизированных нервных проводниках. В
гигантском аксоне кальмара V = 25 м/с (при D= 0,5 мм). В тонких волокнах позвоночных -
около 1 м/с (при D=l мкм).
Остановимся на понятии "протяженности возбужденного участка", т. е. участка, в котором
в данный момент генерируется пик ПД (та или иная его стадия) . Протяженность
возбужденного участка L=V
ПД
• t
ПД
. В гигантском аксоне кальмара L=l мс • 25 мм/мс = 25
мм. В тонких немиелинизированных волокнах при t
ПД
= 1 мс: V = 1 мм/мс; L=l мс •1
мм/мс=1 мм. Проведение возбуждения в миелинизированных нервных волокнах,
характерных для скоростных линий нервной связи у высших животных, осуществляется
тем же способом, что и в немиелинизированных, но здесь имеются некоторые
существенные особенности. Миелиновые муфты, каждая из которых формируется
намоткой одной шванновской клетки на аксон, являются хорошими электроизоляторами:
у миелина (R = 0,16 МОм•см
2
) С= 0,005 мкФ/см
2
. Поэтому в миелинизированном волокне
проводят электрические токи и генерируют ПД в ответ на них по существу только узкие
(∼ 1 мкм) оголенные участки, расположенные между муфтами - перехваты Ранвье, что
было показано Тасаки и Такеути в 1942 г. и Хаксли, Штемпфли в 1949 г. Распространение
ПД здесь осуществляется скачкообразно - "сальтаторно" - от перехвата к перехвату.
Поскольку миелиновые сегменты гораздо протяженнее перехватов (1000 - 2000 мкм
против 1 мкм), то такая форма функционирования проводника экономична в смысле
расхода ионов и нагрузки на ионный насос и обеспечивает существенно большие скорости
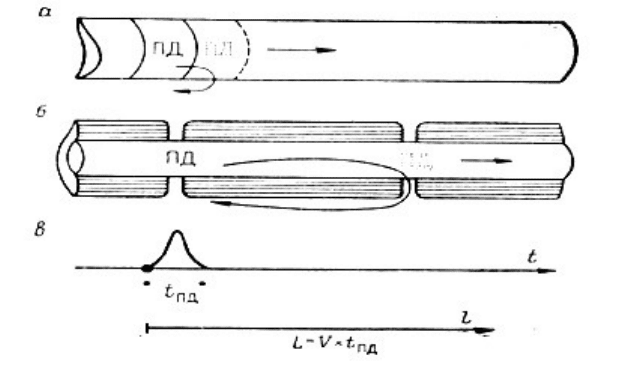
проведения возбуждения (рис.4.1. ).
Рис. 4.1. Сопоставление непрерывного и сальтаторного распространения потенциала
действия.
Схемы: а - непрерывного распространения ПД в немиелинизированном волокне ;
б - сальтаторного проведения в миелинизированном нервном волокне;
в - развитие одиночного пика ПД во времени и распространение его в пространстве.
V
ПД
здесь иначе зависит от диаметра волокна, чем в случае немиелинизированных
проводников. Эмпирически установлено, что в миелинизированных волокнах
позвоночных V
ПД
=КD; ν линейно зависит от D, потому что ν пропорциональна длине
межперехватного участка l, а l линейно зависит от D (при этом время на раздражение
очередного перехвата оказывается приблизительно постоянным). "К" варьирует от 6 до
1,73 для разных групп волокон. У млекопитающих в группе наиболее толстых
миелинизированных A-волокон К=6, т. е у волокон с D= 20 мкм V
ПД
=120 м/с (при этом L =
60 MM), a у волокон с D=4 мкм V
ПД
=24 м/с (L - 24 мм). Обратим внимание на то, что в
последнем случае скорость проведения приблизительно соответствует достигаемой в
гигантском аксоне кальмара (D = 500 мкм). Это наглядно демонстрирует прогрессивность
миелинизации, позволяющей достигать высоких скоростей нервнойсигнализации при
малой толщине нервных проводников. И понятно, что нервные болезни, при которых
происходит демиелинизация, характеризуются сильными расстройствами функций
нервной системы.
В заключение отметим особенности проведения ПД в участках резкого изменения свойств
нервного проводника. Таковым является, например, переход миелинизированного
нервного волокна в оголенное и разветвленное окончание. Площадь мембраны довольно
протяженного окончания обычно значительно (на 2 порядка) больше площади перехвата
Ранвье (хотя окончание тоньше магистрального волокна). При таком соотношении
площадей проведение ПД из перехвата в окончание (например, моторное) несколько
замедленно и имеет сниженный ГФ. Это объясняется падением плотности локального
раздражающего тока в окончании. Проведение же из окончания (например, сенсорного) в
перехват ускорено и облегчено из-за концентрации тока ПД окончания в перехвате.
Аналогичная ситуация наблюдается в участках разветвления магистральных нервных
волокон (а также проводящих ПД дендритов).
Поскольку рабочая площадь мембраны в неразветвленной части меньше, чем в
разветвленной, то проведение ПД из первой во вторую происходит с некоторым
замедлением и при сниженном ГФ]. Наоборот, синхронные ПД ветвей взаимно облегчают
себе вход в неразветвленную часть проводника. Наконец, сходные явления имеют место в
участках резкого изменения диаметра проводника, например, в районе перехода тела
нейрона в начальный сегмент аксона или дендрит. Здесь взаимодействующие соседние
участки мембраны также имеют существенно разную по величине рабочую площадь. И
это обстоятельство при прочих равных условиях способствует проведению ПД из
расширенного сегмента в суженный (например, из сомы в дендрит) и затрудняет
проведение в противоположном направлении.
Лекция 5. Электрофизиология нервного
ствола
Нервный ствол (или нерв) представляет собой пучок множества (тысячи) нервных
волокон, одетых общими эпителиальной и соединительнотканной оболочками. Обычно
нерв включает в себя волокна различного типа и разного диаметра. Например, в
седалищном нерве лягушки имеются двигательные волокна (аксоны мотонейронов),
чувствительные (аксоны чувствительных клеток) и вегетативные (аксоны нейронов
симпатических ганглиев). Моторные и наиболее толстые чувствительные волокна этого
нерва миелинизированы, симпатические - не имеют миелиновой оболочки, но покрыты
одним слоем шванновских клеток.
Все эти волокна в покое имеют поляризованную поверхностную мембрану. Как уже
говорилось, если нерв перерезать и соединить с электрорегистрирующим устройством его
поперечный разрез и продольную поверхность, то регистрируется потенциал повреждения
или иначе потенциал покоя (ПП) нормальной точки нерва относительно поврежденной.
ПП имеет обычно небольшую величину - 20-30 мВ, он меньше МПП волокон нерва, что
объясняется шунтирующим действием межволоконных щелевых пространств. Если в
сегменте нерва, расположенном между электродами, заменить (перфузией) межклеточную
жидкость на раствор сахарозы ("сахарозный мостик"), то ПП нерва поднимается до -70
мВ. Его величина, по существу, отражает величину МПП наиболее толстых ?-волокон
группы А, генерирующих основную часть внешнего тока.
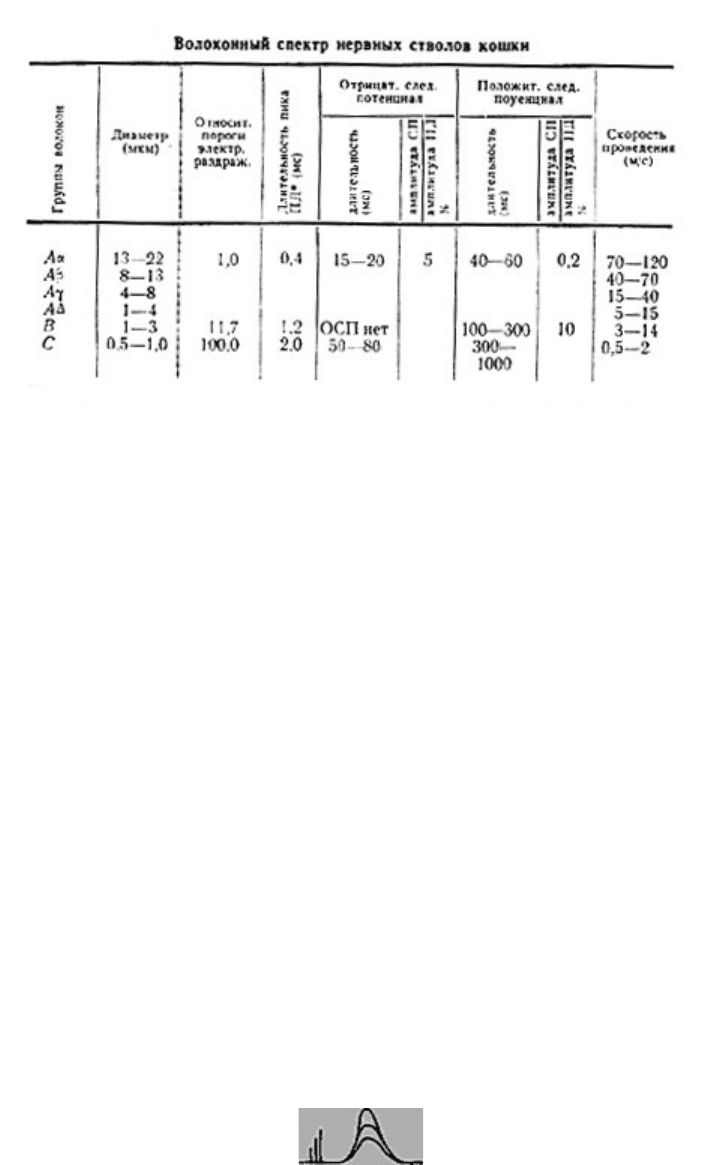
Потенциалы действия различных групп волокон нервных стволов позвоночных
исследованы в классических работах Эрлангера и Гассера (1937). Авторы установили, что
в нервах лягушки и кошки имеются три основные группы волокон (А, В и С),
различающиеся по электровозбудимости, по скорости развития и компенсации ПД и по
скорости проведения (все эти показатели в ряду А-В-С падают). Группа А включает
наиболее толстые хорошо миелинизированные моторные и чувствительные волокна; В -
слабо миелинизированные преганглио-нарные (парасимпатические) волокна; С -
немиелинизирован-ные постганглионарные (симпатические) волокна. Группа А
неоднородна, в ней по тем же показателям можно выделить подгруппы; в этом ряду
названные показатели тоже падают. Заметим, что указанные соотношения порогов
электрического раздражения групп волокон не отражают точного соотношения
электровозбудимости их мембран. Высокие относительные пороги тонких волокон при их
раздражении в нервном стволе определяются в основном тем обстоятельством, что тонкие
волокна по сравнению с толстыми обладают более высоким внутренним сопротивлением.
В них входит такая малая часть раздражающего тока, что при пороговой силе для Aα-
волокон, она совершенно недостаточна для создания на мембране более тонких волокон
сколько-нибудь существенной деполяризации. За вычетом этого фактора различия
электровозбудимости А, В, и С групп волокон, по-видимому, гораздо менее значительны.
Суммарная (многоволоконная) электрическая активность нерва (нервного ствола)
создается его волокнами, каждое из которых генерирует свой стандартный по амплитуде и
временным параметрам ПД, распространяющийся в обе стороны от точки, к которой
приложено раздражение. Суммарный электрический сигнал нерва зависит от числа
активных волокон, синхронности их активности, способа отведения и других
обстоятельств.
Рассмотрим случай так называемого однофазного отведения, при котором один
отводящий электрод (активный) стоит на нормальном участке ствола, а другой
(индифферентный) на поврежденном участке, где волокна деполяризованы полностью
(рис.5.1.). Пусть применяется искусственное раздражение и раздражающий электрод
(катод) расположен достаточно близко (l==3 мм) от активного отводящего электрода, а
нерв находится в непроводящей среде - масле или воздухе. В этой ситуации, когда
сильное одиночное раздражение приводит к синхронному возбуждению всех волокон,
активным отводящим электродом регистрируется суммарный ПД нерва, по форме
приближающийся к ПД отдельного α-волокна, но немного более затянутый во времени.
Этот ПД нерва, однако, не подчиняется правилу "все или ничего", но зависит от силы
раздражения: на пороге ПД ничтожно мало, с увеличением силы стимулов он постепенно
растет, достигая максимума, равного при обычных условиях нескольким милливольтам
(5-10 мВ), а в условиях сахарозного мостика 50-100 мВ. При дальнейшем увеличении
силы стимулов этот ПД несколько удлиняется во времени. Все эти изменения амплитуды
и длительности пика ПД нерва

Рис.5.1 . Развитие пика составного потенциала действия нервного ствола.
а - схема раздражения и однофазного отведения ПД нервного ствола,
перфузированного высокоомным раствором сахарозы, б - рост амплитуды
составного ПД при усилении стимулов. Схемы петления: в - тока одиночного
возбужденного волокна на окружающие его покоящиеся волокна, г - токов двух
соседних возбужденных волокон, д и е - эквивалентные схемы двух соседних волокон
при возбуждении одного из них (д) и при синхронной активации обоих в соседних
точках (е).
при усилении стимула определяются ростом числа активных волокон, подключением к
низкопороговым и быстрым а-волокнам, более высокопороговых медленных β, γ, Δ
волокон группы А, а затем В и, наконец С-группы.
Но рост амплитуды суммарного ("составного") ПД нерва при увеличении числа
синхронно активных волокон не имеет в своей основе истинного сложения их ЭДС.
Волокна в нерве подключены не последовательно (лишь тогда их ЭДС бы складывались),
а параллельно. Причину роста составного ПД при увеличении числа активных волокон
можно понять, рассмотрев схему на рис. , из которой видно, что неактивные волокна
шунтируют активные. При активизации неактивных волокон этот шунт снимается. Кроме
того, при этом падает суммарное внутреннее сопротивление источника .
Рассмотрим случай, когда при тех же прочих условиях существенно (до 80-100 мм)
увеличено расстояние между раздражающим электродом (катодом) и активным
отводящим электродом (рис.5.2 ). При этом на пороговые раздражения и