Проничев И.В. Лекции по физиологии центральной нервной системы
Подождите немного. Документ загружается.

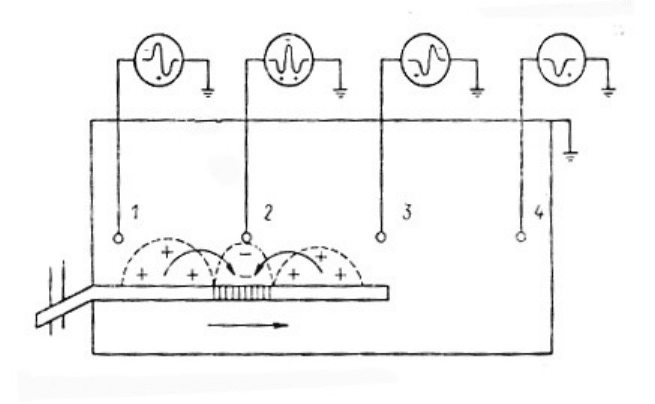
Рис.5.2. Схемы дисперсии компонентов составного однофазного потенциала
действия нерва (связанных с А (α, β, γ, Δ) В- и С-волокнами) и формирования
двухфазного потенциала действия.
а, б - варианты однофазного отведения ПД при малой (а) и большой (б) дистанциях
проведения, в, г - формы составных ПД при данных дистанциях и разных силах
раздражения (при большой дистанции и сверхпороговом раздражении пик
составного потенциала больше и разделен на ряд последовательных зубцов,
порождаемых волокнами с разными скоростями проведения), д, е - схемы
двухфазного отведения (d) и формирования асимметричной двухфазной кривой (е)
из однофазных сигналов двух отводимых точек (1 и 2).
при небольшом превышении порога получается в общем такой же Аα-пик. Но по мере
роста предъявляемых стимулов этот Аα--пик не просто увеличивается, а осложняется
рядом дополнительных более поздних пиков, следующих за Аα-пиком. Это Аβ-, Аγ и АΔ-,
В и С-пики. Такое их распределение -результат отставания от Аα--импульса импульсов
Аβ,γ и Δ, В и С волокон, называемое их дисперсией во времени. Степень ее у одиночного
залпа первоначально синхронных импульсов тем больше, чем больше дистанция
проведения и чем больше различия в скорости проведения у сигналов волокон разных
групп (рис.5.2.).
Рассмотрим вариант так называемого двухфазного отведения ПД, при котором оба
электрода расположены на нормальных неповрежденных участках нерва. Это отведение
применяют в том случае, если нервный проводник почему-либо надо сохранить в целости.
При двухфазном отведении картина сложна, даже если дистанция проведения мала и нет
существенной временной дисперсии волоконных сигналов. Дело в том, что каждый из
электродов при этом отводит свой составной ПД (ПД в своей точке) и эти два ПД
алгебраически складываются на входе регистрирующего прибора. На схеме (рис.5.2.)
видно, что при этом сложении получается двухфазная волна и что в силу
несимметричности пиков ПД первая фаза этой волны выше
Рис.5. 3 . Схема отведения распространяющего пика потенциала действия нерва на
некотором расстоянии от него в проводящей среде.
1-у левого края фильтровальной бумажки, 2 - на уровне средней части проводника, 3
- на конце нерва, 4 - у правого края фильтровальной бумажки.
второй, а вторая длиннее первой. При большей дистанции проведения эти фазы, кроме
того, осложняются соответствующими дополнительными пиками.
Наконец, рассмотрим так называемое фокальное отведение (рис.5.3.) пика ПД нервного
ствола, расположенного в проводящей среде. При таком отведении один электрод
приближен к нерву, а второй (индифферентный) "бесконечно" удален. Этот случай
моделирует регистрацию электрического сигнала нейрона или группы нейронов в мозге с
помощью внеклеточного фокального микроэлектрода. Лоренте де Но проанализировал
этот вариант отведения в опыте на седалищном нерве лягушки, расположенном на листе
фильтровальной бумаги, смоченном в растворе Рингера. Зарегистрировав фокальным
электродом потенциалы в разных точках проводящей среды в определенные моменты
возбуждения нерва (например, при достижении вершины а-пика ПД в средней части
нерва), автор смог получить картины электрического поля, возникающего в проводящей
среде вокруг активного нерва (см. рис.5.3.). Представленная на рис. схема - это
изображение электрического поля на плоскости. Но вращая рисунок вокруг оси нерва,
можно получить соответствующее объемное поле. Обратим внимание на то, что нерв в тот
момент, когда он оказывается возбужденным в средней части, имеет вид генератора с
тремя полюсами - двумя плюсами по концам и минусом в средней части. И электрическое
поле в среде имеет три соответственные зоны, разделенные на плоскости линиями (а в
объеме поверхностями) нулевых сдвигов U. Заметим, что по мере удаления от нерва в
любую сторону величины сдвигов U падают, на краях бумажки они близки к 0. Когда
возбуждение нерва (его составной ПД) распространяется слева направо, то движется и
порождаемое им электрическое поле в среде. При этом стоящий на средней линии
фокальный электрод, мимо которого движется ПД,
регистрирует трехфазный сигнал, тем более слабый,чем дальше этот электрод отнесен от
нерва (в поперечном к его оси направлении). Если отводящий фокальный электрод
помещается в месте зарождения поля ПД, т. е. на нашей схеме на левом краю
фильтровальной бумажки, где нерв на нее выходит из масляной ванночки (в которой он
контактирует с раздражающими электродами), то при пробеге ПД регистрируется
двухфазный сигнал . Если же фокальный электрод помещается на правом краю бумажки,
куда нерв не дотягивается и ПД соответственно не доходит, то при пробеге ПД
регистрируется однофазный сигнал положительной полярности. Заметим, что эти формы
фокального электрического сигнала, порождаемого полем проводящегося ПД,-
трехфазная, двухфазная и однофазная характерны соответственно для случаев прохода ПД
мимо электрода, зарождения ПД около электрода и ухода от него и, наконец, движения
ПД к электроду с остановкой "на полпути".
Все рассмотренные до сих пор варианты записи электронейрограмм получаются при
синхронной электрической активности всех волокон нервного ствола в условиях
искусственного электрического раздражения нерва.
Но в естественных условиях отдельные волокна нервного ствола получают раздражение
(тоже электрическое) из своих индивидуальных источников, например моторные волокна,
из тел мотонейронов, а чувствительные - из рецепторов. Волокна нервного ствола при
этом работают чаще всего асинхронно, а чувствительные и двигательные, кроме того,
проводят ПД в разных направлениях. При этом из-за действия шунтов суммарная
электрическая активность нервного ствола оказывается очень ослабленной, ее анализ
весьма труден.
В заключение коснемся вопроса об электрических взаимоотношениях работающих
соседних волокон в нервных стволах. На схеме (рис.5.1.) видно, что соседние волокна
подключены друг к другу, как шунты, через межклеточную щель. И если одно из этих
волокон работает, то внешние петли тока, порождаемого его ПД, затекают в соседние
волокна, используемые, таким образом, как часть внешней электропроводности
(проводимости). Сила тока этих петель при активности малого числа волокон не
достаточна для раздражения. Поэтому в условиях асинхронной активности волокна
функционируют вполне изолированно друг от друга. Но при синхронной активации
значительной части волокон нерва суммарное поле их ПД порождает более сильный ток,
приближающийся к пороговому для волокон. При этом возможно, например, ускорение
проведения импульсов по более медленным волокнам за счет добавления соседнего
электрического влияния от более быстро проводящих. Такой эффект отчетливо показали
Катц и Шмидт в 1940 г. на нерве ракообразного, содержащем всего 2 волокна. Если
искусственно повысить электровозбудимость волокон нерва, то, как показал еще в 1937 г.
Д. Г. Квасов, возбуждение части волокон влечет за собой "соседнее" электрическое
раздражение остальных. Такой эффект между прочим лежит в основе уравнительной
стадии парабиоза нерва, описанного Н. Е. Введенским.
Если синхронно и в одной точке нерва возбуждается большая часть его волокон, например
все А-волокна, то получается еще один эффект. Происходит взаимное вычитание внешних
петель биотоков соседних возбужденных волокон. Это приводит к снижению силы токов,
связывающих возбужденные участки этих волокон с покоящимися, подлежащими
раздражению. В итоге на несколько процентов падает скорость проведения их ПД и ГФ1
этого проведения. В нормальном проводнике такое снижение ГФ! практически не
сказывается на функции. Но если нерв локально обработан каким-либо повреждающим
агентом и в этом месте (участок парабиоза) его волокна исходно имеют ГФ1∼1, то
синхронный залп ПД совокупности его волокон, распространившись до участка
парабиоза, может не пройти через этот участок из-за рассмотренного дополнительного
снижения ГФ. И чем больше волокон синхронно возбуждено (т. е. чем сильнее
приложенное к нерву раздражение), тем вероятнее такой блок. В этом, по нашим данным,
состоит основная причина парадоксальной стадии парабиоза нерва.
Лекция 6. Синапсы.
Синапсами называют специализированные контакты между клетками, используемые для
передачи сигналов. Синапсы можно классифицировать, во-первых, по их
местоположению и принадлежности соответствующим клеткам (нервно-мышечные,
нейро-нейрональные, аксо-соматические, аксо-дендритические и т. д.). Во-вторых,
синапсы можно разделить по знаку их действия на возбуждающие и тормозящие. И
наконец, по способу передачи сигналов они разделяются на электрические, в которых
сигналы передаются электрическим током, и химические, в которых передатчиком
сигнала (трансмиттер) или иначе посредником (медиатор) является то или иное
физиологически активное вещество. Существуют и смешанные - электро-химические
синапсы. Заметим, что и в том, и в другом синапсе имеются такие компоненты, как
пресинаптическая мембрана, постсинаптическая мембрана и разделяющая их
синаптическая щель.
Синапсы классифицируются также в зависимости от того, чем они образованы. Например,
контакт, образуемый аксоном на теле (соме) клетки, называется аксо-соматическим
синапсом, а контакт на дендрите называется аксо-дендритным синапсом (рис.6.1.).
Подобно этому, контакт между двумя аксонами называется аксо-аксонным синапсом (В,
Д), а контакт между двумя дендритами - дендро-дендритным (Б).
В мозге редко встречаются изолированные одиночные синапсы. Обычно несколько
синапсов вместе складываются в тот или иной тип групповой синаптической связи.
Простейший из таких типов - когда два или несколько синапсов расположены рядом друг
с другом и ориентированы в одном направлении; все они бывают аксо-дендритными.
Более сложен тип, в котором отросток а образует синапс на отростке б, а отросток б на
отростке в. Такая ситуация схематически показана на рис.6.1.Д. Такие синапсы называют
последовательными; их примерами могут служить аксо-аксо-дендритные и аксо-дендро-
дендритные последовательности.
Еще в одном типе отросток а соединяется с отростком б, а последний - снова с отростком
а. Эта ситуация схематически показана на рис.6.1.Г. Такой синапс принято называть
реципрокным. Если два таких синапса расположены рядом, то их называют реципрокной
парой. Если же два синапса удалены один от другого, то возникает реципрокное
устройство. Наконец, есть такие типы синаптических соединений, когда тесно сближена
целая группа терминалей. Этот тип называют синаптической гломерулой (рис.6.1.Е).
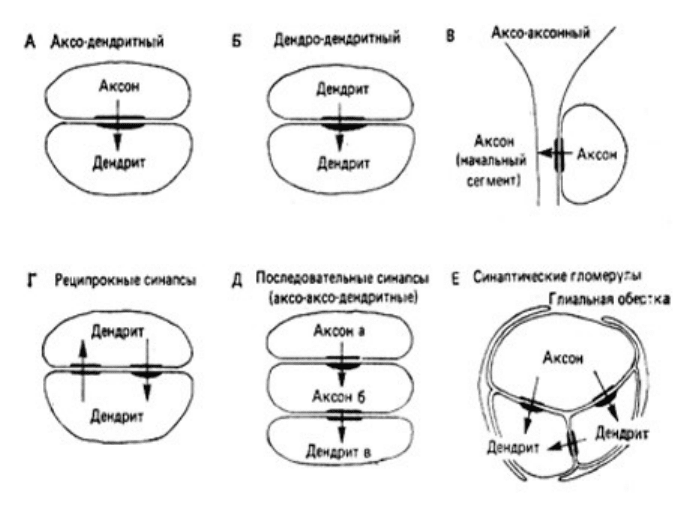
Рис.6.1. Типы синаптических связей (по Г.Шеперд,1987).
Первыми синапсами, которые удалось идентифицировать с помощью электронной
микроскопии, были простые контакты терминалей, относимые к аксо-соматическому и
аксо-дендритному типам. Поскольку эти простые типы соответствовали представлению о
"поляризованном" нейроне, их стали считать "классическими" синапсами. Позднее были
идентифицированы аксо-аксонные и дендро-дендритные типы синапсов. Тогда же были
обнаружены последовательные и реципрокные синапсы, а также различные типы
специализированных синаптических контактов и терминалей. Поскольку такие синапсы,
терминала и типы выходят за рамки классических представлений, то на практике простые
синапсы стали называть стандартными, а все остальные - нестандартными или даже
"необычными".
6.1. Электрофизиология синапсов
Электрический синапс
Начнем с рассмотрения более простого - электрического - синапса. Надо заметить, что
существование таких синапсов предполагалось давно, но выявлены и изучены они были
лишь в самое последнее время Электрические синапсы имеются в нервной системе как
беспозвоночных, так и позвоночных животных, но наиболее изученными являются такие
синапсы у беспозвоночных. Всем синапсам электрического типа свойственны а) очень
узкая синаптическая щель ("5 нм, иначе 50 А) и б) очень низкое удельное сопротивление
пре- и постсинаптических мембран. Последнее чаще всего связано с существованием
транссинапти-ческих каналов (D=l -1,5 нм), проходящих поперек синаптической щели в
специальных тельцах, связывающих пре- и постсинаптическую мембраны . Например, в
простейшем возбуждающем электрическом синапсе - в так называемом септальном
синапсе соседних сегментов гигантского аксона рака - удельное сопротивление
перегородки (септы) составляет 1,0 Ом • см
2
, в то время как сопротивление наружной
мембраны каждого сегмента - 1000-3000 Ом • см
2
. В таком синапсе ПД возбужденного
пресинаптического сегмента посредством петли электрического тока, входящего через
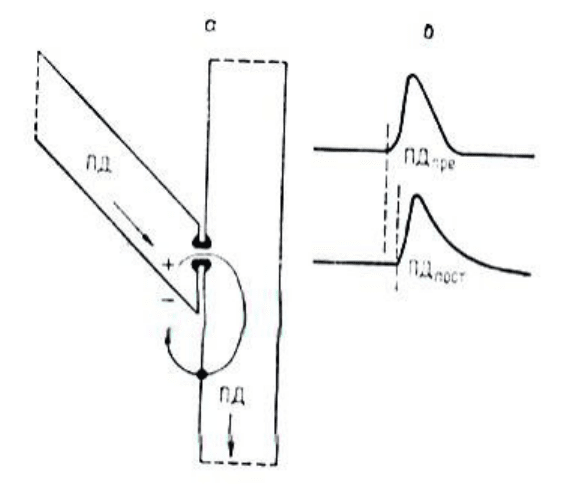
септальную мембрану и выходящего через наружную мембрану постсинаптического
сегмента, раздражает этот последний и вызывает ПД. При этом все же имеется некоторая
потеря силы раздражающего тока на перегородке, поскольку она имеет очень малую
площадь и ее общее сопротивление достигает 0,2-0,4 МОм. В результате проведение ПД
через
2. Схема работы возбуждающего электрического синапса (а) и временные
соотношения пресинаптического и постсинаптического ПД (б).
септальный синапс осуществляется с синаптической задержкой порядка 0,1 мс, которая,
однако, как увидим далее, и в этом и в других электрических синапсах гораздо короче,
чем задержка в химических синапсах.
В септальных синапсах, как и в непрерывном нервном проводнике, проведение
осуществляется в обе стороны.
Некоторые электрические возбуждающие синапсы работают как "вентильные"
механизмы, т. е. передают возбуждение, по существу, односторонне. Последнее
объясняется выпрямляющими свойствами их синаптических мембран, т. е., по-видимому,
тем, что их каналы открыты лишь для электрического тока одного определенного
направления. При этом сопротивления прямому и обратному токам могут различаться в 50
раз. Так, например, работает гигантский синапс рака - электрический синапс между
латеральным гигантским аксоном и гигантским моторным волокном . Из латерального
аксона в моторный импульс проходит с задержкой равной 0,1 мс, а в противоположном
направлении - совсем не проходит . В последнем случае за синапсом оказывается лишь
электротонический потенциал менее 0,3 мВ.
Заметим, что электрическим синапсам свойственно чрезвычайное быстродействие и
высокая надежность передачи. Однако эти синапсы как не включающие никакого
инерционного звена мало приспособлены для интегрирования серии импульсов
возбуждения.
Специализированные тормозящие электрические синапсы встречаются крайне редко. Они
описаны Фурукавой, Фуршпаном на маутнеровских клетках рыб. Здесь пресинаптическое
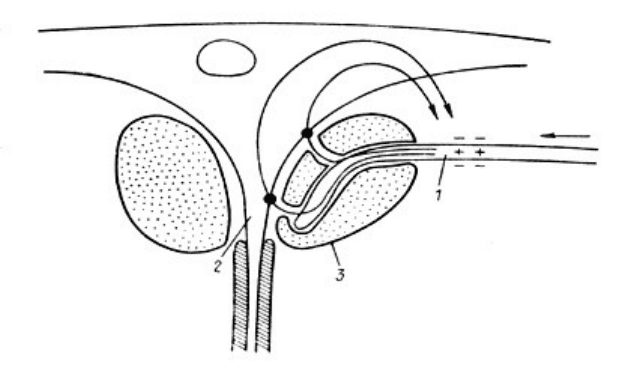
волокно не образует контакта, а лишь близко подходит к аксонному холмику - выходному
участку маутнеровской клетки, будучи окруженным относительно высокоомным
материалом аксонной чашечки ПД волокна, направляющийся в терминаль, видимо,
останавливаясь в начале терминали, создает ток, входящий в аксонный холмик. Этим
током мембрана аксонного холмика, наиболее возбудимая зона маутнеровской клетки,
гиперполяризуется, чем и достигается очень быстрый кратковременный тормозящий
эффект (рис.6.3.).
Рис.6.3. Вероятная схема работы тормозного электрического синапса на
маутнеровском нейроне рыбы.
1 - нервное окончание, 2 - начальный сегмент аксона маутнеровской клетки, 3 -
глиальная "чашечка". Стрелкой показано направление тока, порождаемого ПД
нервного окончания.
Химический синапс
Обратимся к химическим синапсам. Классическим модельным объектом здесь является
возбуждающий нервно-мышечный синапс скелетной мускулатуры позвоночных,
действующий с помощью медиатора ацетилхолина (АХ). Электрофизиология этого
синапса подробно проанализирована в работах А. Ф. Самойлова, Куффлера, А. Г.
Гинецинского и Катца. Этот и все известные химические синапсы в отличие от
электрических характеризуются, а) относительно широкой синаптической щелью (20-80
им, т. е. 200-800 А) и б) относительно высоким (обычным) удельным электрическим
сопротивлением контактирующих мембран. Их морфологической особенностью, кроме
того, являются пресинаптические везикулы с медиатором (рис.6.4.). При таких структуре
и свойствах синапса пресинаптический ПД не может петлей своего тока возбудить
постсинаптическую клетку.
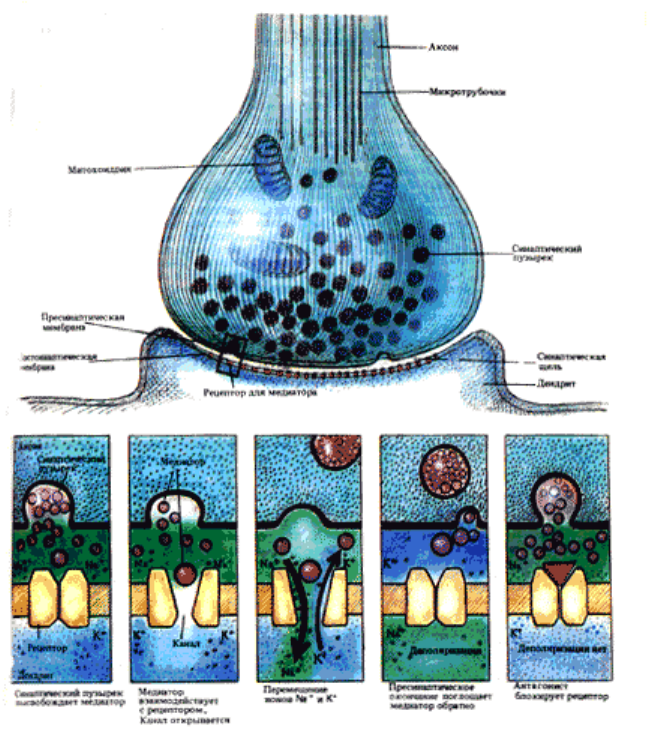
Рис.6.4. Схема работы возбуждающего химического синапса .
Дело в том, что ток, выходящий через пресинаптическую мембрану, здесь почти целиком
уходит через широкую синаптическую щель мимо постсинаптической клетки,
обладающей значительным входным сопротивлением . А очень небольшая часть этого
тока, которая все же входит в постсинаптическую клетку, вызывает лишь ничтожное
изменение ее МП. Суть работы химического синапса состоит в следующем.
Пресинаптический ПД здесь работает как инициатор нейро-секреторного акта. При
развитии ПД терминали (а также и при искусственной ее деполяризации) в нее из среды
входят ионы кальция. А это стимулирует выброс медиатора из 100-200 пресинаптических
везикул в синаптическую щель. Заметим, что выход медиатора очень сильно зависит от
величины деполяризации терминали. Эта зависимость объясняет резкое снижение выхода
медиатора при падении амплитуды ПД терминали. В нормальных условиях в ответ на
нервный импульс освобождается около миллиона молекул АХ. Медиатор диффундирует к
постсинаптической мембране, где для него имеются рецепторы (холинорецепторы- ХР).
При взаимодействии АХ и ХР в постсинаптической мембране открываются
многочисленные ионные каналы (D∼6,5A). Сопротивление этой мембраны падает в 4000
раз, что приводит к ее частичной деполяризации, т. е. к развитию возбуждающего
постсинаптического потенциала (ВПСП). В нервно-мышечном синапсе этот ВПСП
называется потенциалом концевой пластинки (ПКП). ВПСП (ПКП), в свою очередь,
создает ток, раздражающий соседнюю с постсинаптической злектровозбудимую мембрану
мышечного волокна, что и порождает в ней ПД.
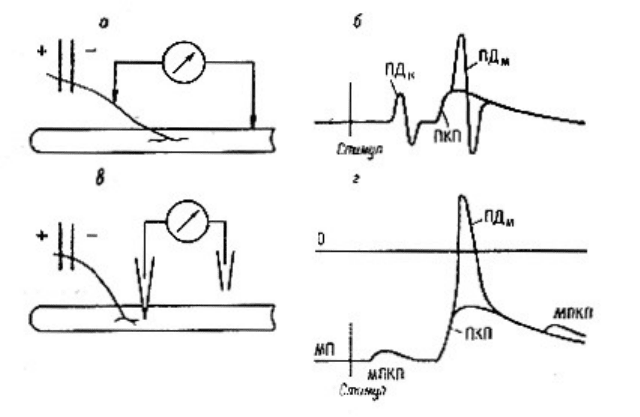
Рис.6.5. Формы основных электрических феноменов, регистрируемых в районе
нервно-мышечного синапса при различных способах отведения.
а - схема внеклеточного отведения (по А . Ф. Самойлову),
б - форма записи ПД нерва ПКП и П мышцы при таком отведении,
в - схема внутриклеточного отведения,
Г - форма записи внутриклеточных ПКП и ПД мышцы.
Таким образом, химическое звено в синапсе выполняет функцию усилителя. Но
синаптическая задержка в химическом синапсе составляет около 0,5 мс. Причем основная
часть этого времени тратится на процесс секреции медиатора.
Химический синапс - это вентильный механизм, рабочий сигнал в нем передается
односторонне (что не исключает обратных связей) (рис.6.5.).
ПКП - локальный потенциал, который злектротонически расползается по мембране с
декрементом. С помощью внутриклеточных микроэлектродов и методики фиксации
потенциала можно изучить токи, текущие через активируемую постсинаптическую
мембрану (рис.6.6.). В подобных экспериментах, варьируя ионный состав среды, А.
Такеучи и Н. Такеучи установили, что АХ открывает в постсинаптической мембране
каналы, пропускающие катионы (Na
+
, K
+
), но не пропускающие анионы.
Позже, изучая ацетилхолиновый шум каналов постсинаптической мембраны, рассчитали
проводимость одиночного канала (20-30 рмО), а также среднее время его жизни (∼1 мс).
Ток концевой пластинки (ТКП) и в клампе, и в обычных условиях представляет собой
результат движения Na
+
и К? по их электрохимическим градиентам.
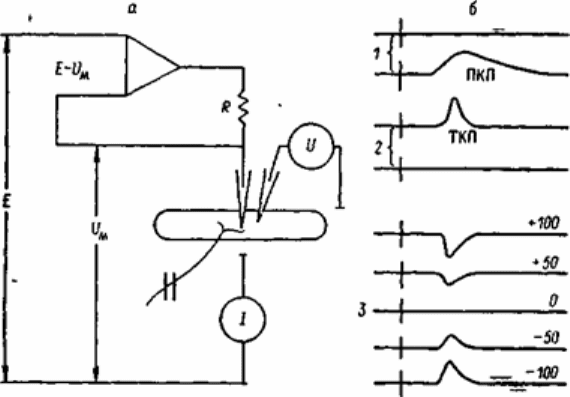
Рис.6.6. Схема методики фиксации потенциала постсинаптической мышечной
мембраны (а) и формы токов концевой пластинки при разных потенциалах
мембраны (б).
1 - запись потенциала концевой пластинки - ПКП - до включения схемы фиксации
потенциала на мембране, 2 - запись тока концевой пластинки (ТПК) при фиксации
потенциала на уровне покоя, 3 - запись ТКП при разных Uм.
При обычных значениях МП (-80, -60 мВ) ток Na
+
направлен внутрь, а ток К
+
наружу,
причем первый больше второго и суммарный (здесь разностный) ток является входящим.
При МП = 0; I
к
=I
Na
и суммарный ток равен 0. А при МП= + 10 мВ, + 20 мВ и т. д. I
к
>I
Na
и
суммарный ток имеет выходящее направление.
МП, при котором I
K
= I
Na
и суммарный ток равен 0, называют потенциалом реверсии ТКП
(E
р
ТКП). По тем же причинам ПКП на фоне нормальных МП (-90, -70 и т. п.) развивается
как деполяризация (20-30 мВ). При МП = 0 ПКП отсутствует (E
р
ПКП), а при
положительных МП развивается как гиперполяризация.
Формы ТКП (в клампе) и ПКП (рис.6.6.) несколько различаются. ТКП короче, основная
часть его соответствует по длительности восходящей фазе ПКП. Соответственно у ПКП
лишь восходящая фаза создается трансмембранными ионными токами. Относительно
более медленный спад ПКП определяется постепенной зарядкой мембранной емкости и
пропорционален тм. При частом следовании ПКП могут суммироваться и создавать
стационарную деполяризацию.
Помимо рабочих ПКП (ВПСП), инициируемых нервным импульсом, в химических
синапсах имеются спонтанные обычно редкие миниатюрные ПКП (ВПСП), составляющие
доли милливольта и обозначаемые МПКП (МВПСП) . Они отражают спонтанный выброс
одиночных квантов медиатора (т. е. содержимого одиночных везикул) и реакцию на них
постсинаптической мембраны. Как уже указывалось, рабочий ПКП складывается из 100-
200 МПКП, это число называют квантовым составом ПКП. При ритмической активности
ПКП (ВПСП) могут постепенно возрастать по амплитуде (потенциация) из-за накопления
Са
2+
в пресинапсе или снижаться (депрессия) из-за истощения запаса квантов медиатора.
Возбуждающие химические синапсы