Townsend C.R., Begon M., Harper J.L. Essentials of Ecology
Подождите немного. Документ загружается.


to reproduction overall but large individual seeds. By contrast, in more disturbed,
open, r-selecting habitats, the plants tend to conform to the general syndrome of
r characteristics: a greater reproductive allocation, but smaller seeds, smaller size,
earlier reproduction and a shorter life (Figure 5.29).
On the other hand, there seem to be about as many examples that fail to fit the
r/K scheme as examples that correspond. One might regard this as a damning
criticism of the r/K concept, since it undoubtedly demonstrates that the explanatory
powers of the scheme are limited. But it is equally possible to regard it as very
satisfactory that a relatively simple concept can help make sense of a large pro-
portion of the multiplicity of life histories. Nobody, though, can regard the r/K
scheme as the whole story. Like all attempts to classify species and their character-
istics into pigeonholes, the distinction between r and K species has to be recognized
as a convenient (and useful) human creation rather than an all-encompassing
statement about the living world.
Chapter 5 Birth, death and movement
179
Counting individuals, births and deaths
Ecologists try to describe and understand the distribu-
tion and abundance of organisms. The processes that
change the size of populations are birth, death and
movement. A population is a number of individuals,
but for some kinds of organism, especially modular
organisms, it is not always clear what we mean by
an individual.
Ecologists face enormous problems when they
try to count what is happening to populations
in nature. They almost always estimate rather than
count. There are particular problems in counting
modular organisms and the numbers of births and
deaths.
Life cycles and reproduction
The life histories of all unitary organisms can be seen
as variations around a simple, sequential pattern.
Some organisms fit several or many generations within
a single year, some breed predictably just once each
year (annuals), and others (perennials) have a life
cycle extended over several or many years. Some,
iteroparous species, breed repeatedly; others, semel-
parous species, have a single reproductive episode
followed quickly by death.
Most annuals germinate or hatch in spring, grow
rapidly, reproduce and then die before the end of
summer. Most spend part of the year dormant. There
is a marked seasonal rhythm in the lives of many
long-lived species. Where there is very little seasonal
variation, some reproduce throughout the year; others
have a long non-reproductive phase and then one
lethal burst of reproductive activity.
Monitoring birth and death: life tables and
fecundity schedules
Life tables can be useful in identifying what in a
life cycle is apparently most instrumental in deter-
mining rates of increase or decline. A cohort life
table records the survivorship of members of a
single cohort. When we cannot follow cohorts, it
may be possible to construct a static life table, but
great care is required. The fecundity of individuals
also changes with age, described in age-specific
fecundity schedules.
Ecologists search for patterns of life and death that
we can see repeated in the lives of many species.
A useful set of survivorship curves (types I–III) has
been developed, but in practice patterns of survival
are usually more complex.
SUMMARY
Summary
s
9781405156585_4_005.qxd 11/5/07 14:49 Page 179

Part III Individuals, Populations, Communities and Ecosystems
180
Dispersal and migration
Dispersal is the way individuals spread away from
each other. Migration is the mass directional move-
ment of large numbers of a species from one location
to another. Movement and spatial distribution are
intimately related. Dispersal and migration can have a
profound effect on the dynamics of a population and
on its composition.
The impact of intraspecific competition on
populations
Over a sufficiently large density range, competition
between individuals generally reduces the birth rate
as density increases and increases the death rate
(i.e. is density-dependent). Intraspecific competition
therefore tends to keep density within certain limits
and may thus be said to play a part in regulating the
size of populations.
When populations are sparse and uncrowded
they tend to exhibit exponential growth, but the rate
of increase tends to become reduced by competition
as the population grows, giving rise to population
growth that is not exponential but S-shaped or logistic.
Intraspecific competition also affects net recruitment,
typically resulting in a humped curve.
Life history patterns
There is typically a limited total amount of energy or
some other resource available to an organism for
growth and reproduction. There may be an observable
cost of reproduction. But populations of individuals
that reproduce early in their life can grow extremely
fast.
The potential of a species to multiply rapidly is
favored by natural selection in environments that
are short-lived, allowing the organisms to colonize
new habitats quickly and exploit new resources. Such
species have been called r species. Where there is
often intense competition for limited resources, the
individuals that are successful in leaving descendants
are those that have captured the larger share of
resources, often because they were born larger and/or
have grown faster (rather than reproducing): so-called
K species. The r/K concept can be useful in interpret-
ing many of the differences in form and behavior of
organisms, but of course it is not the whole story.
s
REVIEW QUESTIONS
Review questions
Asterisks indicate challenge questions
1 Contrast the meaning of the word ‘individual’ for
unitary and modular organisms.
2 In a mark–recapture exercise during which a
population of butterflies remained constant in
size, an initial sample provided 70 individuals,
each of which was marked and then released
back into the population. Two days later,
a second sample was taken, totaling 123
individuals of which 47 bore a mark from the
first sample. Estimate the size of the population.
State any assumptions that you have had to
make in arriving at your estimate.
3* Define annual, perennial, semelparous and
iteroparous. Try to give an example of both an
animal and a plant for each of the four possible
combinations of these terms. In which cases is
it difficult (or impossible) to come up with an
example and why?
4 Contrast the derivation of cohort and static
life tables and discuss the problems of
constructing and/or interpreting each.
5 The following is an outline life table and
fecundity schedule for a cohort of a population
of sparrows. Fill in the missing values (wherever
there is a question mark).
9781405156585_4_005.qxd 11/5/07 14:49 Page 180

Chapter 5 Birth, death and movement
181
6 Describe what are meant by aggregated,
random and regular distributions of organisms
in space, and outline, with actual examples
where possible, some of the behavioral
processes that might lead to each type of
distribution.
7* Why is the average density of people in
the United States lower than the density
experienced by people, on average, in
the United States? Is a similar contrast
likely to apply to most species? Why?
Under what conditions might it not
apply?
8* Compare unitary and modular organisms in
terms of the effects of intraspecific competition
both on individuals and on populations.
9 What is meant by the carrying capacity of
a population? Describe where it appears,
and why, in: (i) S-shaped population growth;
(ii) the logistic equation; and (iii) dome-shaped
net recruitment curves.
10 Explain why an understanding of life history
trade-offs is central to an understanding of
life history evolution. Explain the contrasting
trade-offs expected to be exhibited by
r-selected and K-selected species.
R = ?
PROPORTION OF ORIGINAL MEAN NO. OF EGGS
NUMBERS AT START COHORT ALIVE AT START PRODUCED PER INDIVIDUAL
STAGE (X) OF STAGE (A
X
) OF STAGE (l
X
) IN STAGE (m
X
)
Eggs 173 ? 0
Nestlings 107 ? 0
Fledglings 64 ? 0
1-year-olds 31 ? 2.5
2-year-olds 23 ? 3.7
3-year-olds 8 ? 3.1
4-year-olds 2 ? 3.5
9781405156585_4_005.qxd 11/5/07 14:49 Page 181

182
Chapter 6
Interspecific competition
CHAPTER CONTENTS
6.1 Introduction
6.2 Ecological effects of interspecific competition
6.3 Evolutionary effects of interspecific competition
6.4 Interspecific competition and community structure
6.5 How significant is interspecific competition in practice?
Chapter contents
KEY CONCEPTS
In this chapter you will:
l
appreciate the difficulty of distinguishing between the power and
importance of interspecific competition in principle and in practice
l
distinguish between fundamental and realized niches
l
define the Competitive Exclusion Principle and understand its
limitations
l
appreciate the potential role of the evolutionary effects of competition
in species coexistence and the difficulty of proving that role
l
understand the nature and importance of niche complementarity
l
appreciate the difficulties of determining the prevalence of current
competition in nature, and of distinguishing between the effects of
competition and mere chance
Key concepts
9781405156585_4_006.qxd 11/5/07 14:51 Page 182

6.1 Introduction
Having been introduced to intraspecific competition in previous chapters, it is not
difficult to deduce what interspecific competition is. Its essence is that individuals
of one species suffer a reduction in fecundity, survivorship or growth as a result of
exploitation of resources or interference by individuals from another species. These
competitive effects on individuals are likely to affect the population dynamics of
the competing species. These, in turn, can influence the species’ distributions and
also their evolution. The distributions and abundances of species, of course, deter-
mine the compositions of the communities of which they are part. And evolution,
in its turn, can influence the species’ distributions and dynamics.
This chapter, then, is about both the ecological and the evolutionary effects of
interspecific competition on individuals, on populations and on communities. But
it also addresses a more general issue in ecology and indeed in science – that there
is a difference between what a process can do and what it does do: a difference
between what, in this case, interspecific competition is capable of doing and what
it actually does in practice. These are two separate questions, and we must be
careful to keep them separate.
The way these different questions can be asked and answered will be different,
too. To find out what interspecific competition is capable of doing is relatively easy.
Species can be forced to compete in experiments, or they can be examined in nature
in pairs or groups chosen precisely because they seem most likely to compete.
But it is much more difficult to discover how important interspecific competition
actually is. It will be necessary to ask how realistic our experiments were, how
typical they were of the way species interact in nature, and how typical of pairs
and groups of species generally were those singled out for special attention.
We begin, though, with some examples of what interspecific competition can do.
6.2 Ecological effects of interspecific
competition
6.2.1 Competition between diatoms for silicate
Competition was investigated in the laboratory between two species of fresh-
water diatoms (single-celled plants), Asterionella formosa and Synedra ulna, both
of which require silicate in the construction of their cell walls (see Section 3.5).
Chapter 6 Interspecific competition
183
Interspecific competition is one of the most fundamental phenomena in ecology,
affecting not only the current distribution and success of species but also their
evolution. Yet the existence and effects of interspecific competition are often
remarkably difficult to establish and demand an armory of observational,
experimental and modeling techniques.
two separate questions –
the possible and actual
consequences of competition
9781405156585_4_006.qxd 11/5/07 14:51 Page 183
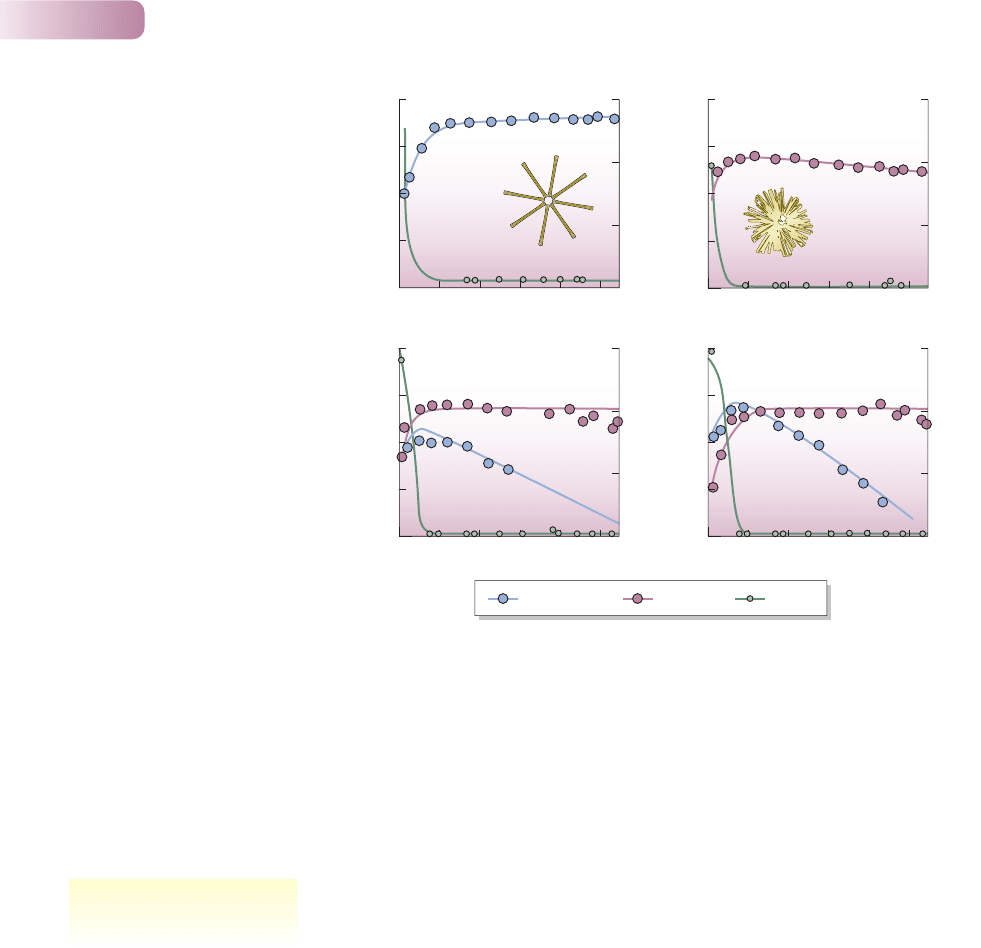
The population densities of the diatoms were monitored, but at the same time
their impact on their limiting resource (silicate) was also being recorded. When
either species was grown alone in a liquid medium to which resources were con-
tinuously being added, it established a steady population density while reducing
the silicate to a constant low concentration (Figure 6.1a, b). However, in exploit-
ing this resource, Synedra reduced the silicate concentration to a lower level than
did Asterionella. Hence, when the two species were grown together, Synedra
maintained the concentration at a level that was too low for the survival and
reproduction of Asterionella and only Synedra survived (Figure 6.1c).
Thus, although both species were capable of living alone in the laboratory
habitat, when they competed, Synedra excluded Asterionella because it was the
more effective exploiter of their shared, limiting resource. A similar result has
been obtained for the nocturnal, insectivorous gecko Hemidactylus frenatus,
an invader of urban habitats across the Pacific basin, where it is responsible
for population declines of the native gecko Lepidodactylus lugubris (Petren and
Case, 1996). The diets of the two geckos overlap substantially and insects are a
limiting resource for both. The invader is capable of depleting insect resources in
experimental enclosures to lower levels than the native gecko, and the latter suffers
reductions in body condition, fecundity and survivorship as a result.
6.2.2 Coexistence and exclusion of competing
salmonid fishes
Salvelinus malma (Dolly Varden charr) and S. leucomaenis (white-spotted charr)
are morphologically similar and closely related species of salmonid fish (see
Part III Individuals, Populations, Communities and Ecosystems
184
30
20
10
0
30
20
10
0
0 10 20 30 40 50 0 10 20 30 40 50
(c) Interspecific competition
(a) Asterionella alone
Population density (cells ml
–1
)
(b) Synedra alone
Silicate (µmol l
–1
)
Time (days)
10
5
10
4
10
3
10
2
10
1
10
5
10
4
10
3
10
2
10
1
30
20
10
0
30
20
10
0
0 10 20 30 40 50 0 10 20 30 40 50
10
5
10
4
10
3
10
2
10
1
10
5
10
4
10
3
10
2
10
1
Asterionella Synedra Silicate
Figure 6.1
Competition between diatoms.
(a) Asterionella formosa, when grown
alone in a culture flask, establishes a
stable population and maintains a
resource, silicate, at a constant low level.
(b) When Synedra ulna is grown alone it
does the same, but maintains silicate at
an even lower level. (c) When grown
together, in two replicates, Synedra
drives Asterionella to extinction.
AFTER TILMAN ET AL., 1981
more efficient exploiters exclude
less efficient ones
9781405156585_4_006.qxd 11/5/07 14:51 Page 184
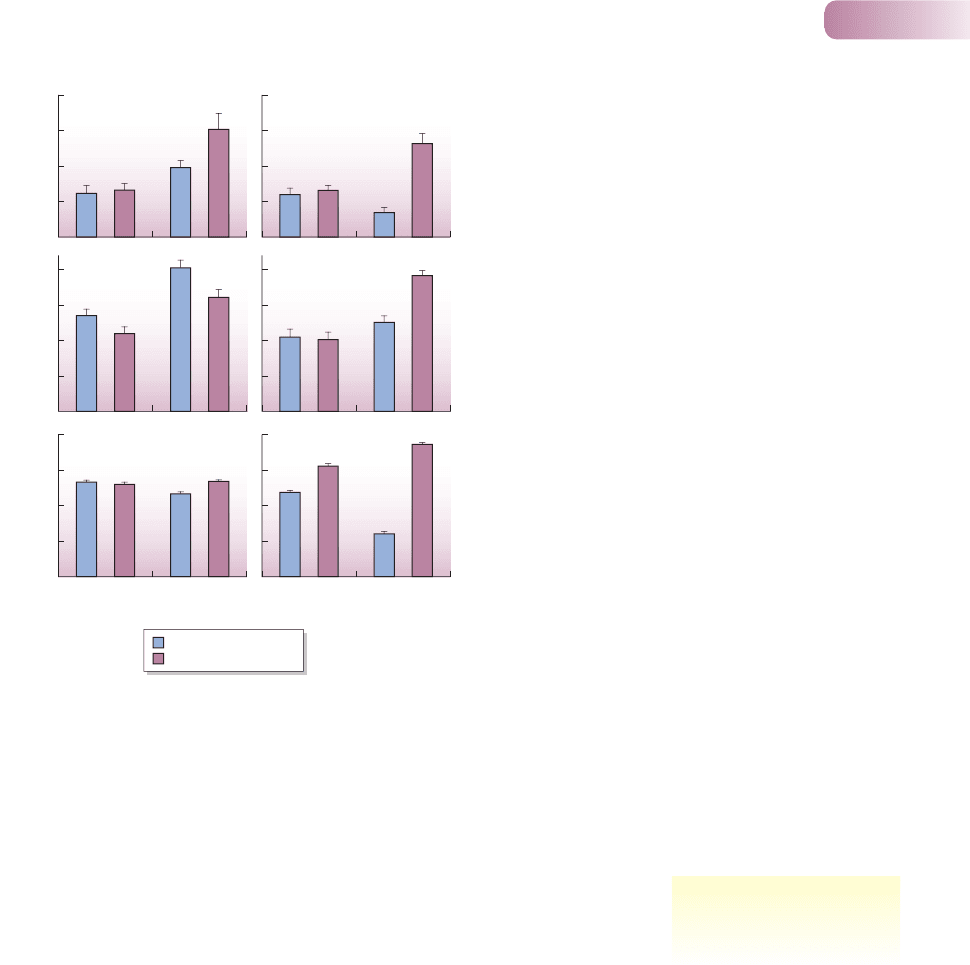
Section 3.2.4). They are found together in many streams on Hokkaido Island in
Japan, but Dolly Varden are distributed further upstream than white-spotted
charr, with a zone of overlap at intermediate altitudes. In streams where one
species is absent, the other expands its range. Water temperature, which has pro-
found consequences for fish ecology, increases downstream.
In laboratory streams, higher temperatures (12°C as compared to 6°C) led
to increased aggression in both species when they were tested alone. But this
effect was reversed for Dolly Varden when white-spotted charr were also pre-
sent (Figure 6.2a). Reflecting this, Dolly Varden charr were suppressed from
obtaining favorable foraging positions and so foraged far less effectively when
white-spotted charr were present at the higher temperature (Figure 6.2b). Also, when
alone, neither species’ growth rates were influenced by temperature, but when both
species were present, growth of Dolly Varden charr decreased with increasing
temperature, whereas that of white-spotted charr increased (Figure 6.2c), such
that the growth rate of Dolly Varden was much lower than that of white-spotted
charr at the higher temperature.
These results are consistent with the hypothesis that the lower altitudinal
boundary of Dolly Varden charr in the Japanese streams was due to temperature-
mediated competition favoring white-spotted charr: they were more aggressive,
foraged more effectively and grew far faster. But the results do not support the
contention that the upper boundary of white-spotted charr is also due to temperature-
mediated competitive difference; that is, Dolly Varden did not outcompete white-
spotted charr in any of the experiments, even at the lower temperatures. Further
Chapter 6 Interspecific competition
185
AFTER TANIGUCHI & NAKANO, 2000
Species togetherSpecies alone(a)
(b)
(c)
Specific growth rate
(day
–1
)
Foraging frequency
(number per 2 min)
Aggressive frequency
(number per 2 min)
2
1
0
2
1
0
0.2
0.1
0
Dolly Varden charr
White-spotted charr
Low High
Temperature treatment
Low High
Temperature treatment
a
a
a
a
a
a
a
a
ab
a
a
a
a
a
b
b
b
b
b
c
c
c
c
d
Figure 6.2
(a) Frequency of aggressive encounters initiated by individuals of
each fish species during a 72-day experiment in artificial stream
channels with two replicates each of 50 Dolly Varden (blue
histograms) or 50 white-spotted charr (maroon histograms) alone
(allopatry) or 25 of each species together (sympatry); (b) foraging
frequency; and (c) specific growth rate in length. Different letters
indicate means are significantly different from each other.
competitive advantage
determined by temperature-
dependent aggressive behaviour
9781405156585_4_006.qxd 11/5/07 14:51 Page 185

work will be needed to determine why Dolly Varden exclude white-spotted
charr upstream.
6.2.3 Some general observations
These two examples illustrate several points of general importance.
1 Competing species often coexist at one spatial scale but are found to
have distinct distributions at a finer scale of resolution. Here, the fishes
coexisted in the same stream, but each was more or less confined to its
own altitudinal zone.
2 Species are often excluded by interspecific competition from locations
at which they could exist perfectly well in the absence of interspecific
competition. Here, Dolly Varden charr can live in the white-spotted charr
zone – but only when there are no white-spotted charr there. Similarly,
Asterionella can live in laboratory cultures – but only when there were
no Synedra there.
3 We can describe this by saying that the conditions and resources provided
by the white-spotted charr zone are part of the fundamental niche of
Dolly Varden charr (see Section 3.6 for an explanation of ecological niches)
in that the basic requirements for the existence of Dolly Varden charr are
provided there. But the white-spotted charr zone does not provide a
realized niche for Dolly Varden when white-spotted charr are present.
Likewise, the laboratory cultures provided the requirements of the
fundamental niches of both Synedra and Asterionella, but those of the
realized niche for only Synedra.
4 Thus, a species’ fundamental niche is the combination of conditions
and resources that allow that species to exist, grow and reproduce when
considered in isolation from any other species that might be harmful to
its existence; whereas its realized niche is the combination of conditions
and resources that allow it to exist, grow and reproduce in the presence of
specified other species that might be harmful to its existence – especially
interspecific competitors.
5 Competing species can therefore coexist when both are provided with a
realized niche by their habitat (in the present case, the stream as a whole
provided a realized niche for both fishes); but even in locations that
provide a species with the requirements of its fundamental niche, that
species may be excluded by another, superior competitor that denies it
a realized niche there.
6 Finally, the fish study illustrates the importance of experimental
manipulation if we wish to discover what is really going on in a natural
population – ‘nature’ may need to be prodded to reveal its secrets.
6.2.4 Coexistence of competing diatoms
Another experimental study of competing diatoms looked at species coexisting
on not one but two shared, limiting resources. The two species were Asterionella
formosa (again) and Cyclotella meneghiniana, and the resources, which were both
Part III Individuals, Populations, Communities and Ecosystems
186
fundamental and realized niches
9781405156585_4_006.qxd 11/5/07 14:51 Page 186

capable of limiting the growth of both diatoms, were silicate and phosphate.
However, whereas Cyclotella was the more effective exploiter of silicate (reducing
its concentration to a lower level), Asterionella was the more effective exploiter of
phosphate. Thus, in cultures where there were especially low supplies of silicate,
Cyclotella excluded Asterionella (Figure 6.3): such cultures failed to provide a
realized niche for Asterionella, the inferior competitor there. Conversely, in
cultures where there were especially low supplies of phosphate, Asterionella
excluded Cyclotella. However, in cultures with relatively balanced supplies of
silicate and phosphate, the two diatoms coexisted (Figure 6.3): with two species,
both provided with sufficient supplies of a resource on which they were superior,
there was a realized niche for both.
6.2.5 Coexistence of competing birds
It is not always so easy to identify the ‘niche differentiation’ or ‘differential
resource utilization’ that allows competitors to coexist. Ornithologists, for
example, are well aware that closely related species of birds often coexist in the
same habitat. For example, five Parus species occur together in English broad-
leaved woodlands: the blue tit (Parus caeruleus), the great tit (P. major), the marsh
tit (P. palustris), the willow tit (P. montanus) and the coal tit (P. ater). All have
short beaks and hunt for food chiefly on leaves and twigs, but at times on the
ground; all eat insects throughout the year, and also seeds in winter; and all nest
in holes, normally in trees. Yet, the closer we look at the details of the ecology of
such coexisting species, the more likely we are to find ecological differences – for
example, in precisely where within the trees they feed, in the size of their insect
prey and the hardness of the seeds they take. We may be tempted to conclude that
such species compete but coexist by eating slightly different resources in slightly
different ways: ‘differential resource utilization’. But in complex natural environ-
ments, such conclusions, while plausible, are difficult to prove.
Indeed, it is often not easy to prove even that the species compete. To do so,
it is usually necessary to remove one or more of the species and monitor the
responses of those that remain. This was done, for example, in a study of two very
similar bird species: the orange-crowned warbler (Vermivora celata) and the
virginia’s warbler (V. virginiae), whose breeding territories overlap in central
Chapter 6 Interspecific competition
187
5
4
3
2
1
0
604020080100
Concentration of PO
4
(µmol I
–1
)
Concentration of SiO
2
(µmol I
–1
)
Cyclotella
excludes
Asterionella
Cyclotella and
Asterionella coexist
Asterionella
excludes Cyclotella
Figure 6.3
Asterionella formosa and Cyclotella meneghiniana coexist when
there are roughly balanced supplies of silicate (SiO
2
) and phosphate
(PO
4
), but Asterionella excludes Cyclotella when there are
especially low supplies of phosphate, whereas Cyclotella excludes
Asterionella when there are especially low supplies of silicate.
AFTER TILMAN, 1982
coexistence through niche
differentiation – and even
competition – may be
difficult to prove
9781405156585_4_006.qxd 11/5/07 14:51 Page 187
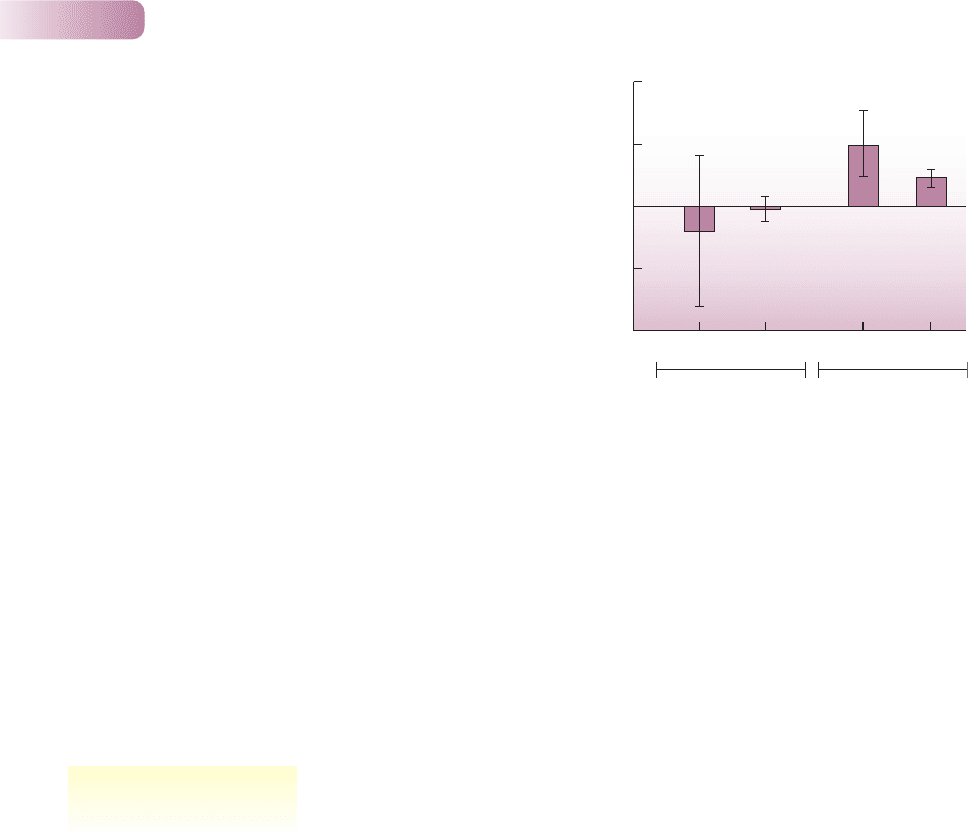
Arizona. On plots where one of the two species had been removed, the remain-
ing species fledged between 78% and 129% more young per nest. The enhanced
performance was due to improved access to preferred nest sites and consequent
decreases in the loss of nestlings to predators. In the case of virginia’s warblers,
but not orange-crowned warblers, feeding rate also increased in plots from which
the other species was removed (Figure 6.4).
6.2.6 Coexistence of competing rodents and ants
The examples described so far have all involved pairs of closely related species
– diatoms, salmonid fish or birds. This is potentially misleading in at least two
important respects. First, competition may occur amongst larger groups of species
than just a pair – where it is sometimes, therefore, called ‘diffuse’ competition.
And second, competition may occur between completely unrelated species.
Both points are illustrated by a study of interspecific competition involving
seed-eating ants and seed-eating rodents in deserts of the southwestern United
States. At the study sites, only two guilds (groups of species that feed on similar
foods in a similar fashion; Root, 1967) fed on seeds: the rodents and the ants.
By studying the size of the seeds harvested by each guild, it was apparent that the
two exhibited significant overlap in the size of the seeds they ate (Figure 6.5).
Ants did eat a larger proportion of the smallest seeds, but overall the potential for
resource competition between them was very high.
As already noted, however, the only true test for whether competition occurs
between them would be to manipulate the abundance of each competitor and
observe the response of its counterpart. Consequently, eight plots were estab-
lished in similar habitats. In two, rodents were trapped and excluded by fencing,
to ensure that only ants now had access to the seeds. In another two, ants were
eliminated by repeated applications of pesticides. In two further plots both ants and
rodents were excluded, and finally two plots were maintained as unmanipulated
controls.
When either rodents or ants were removed, there was a statistically significant
increase in the numbers of the other guild: the depressive effect of interspecific
competition from each guild on the abundance of the other was apparent. Also,
when rodents were removed, the ants ate as many seeds as the rodents and ants
Part III Individuals, Populations, Communities and Ecosystems
188
nstl
–200
inc
0
inc nstl
–100
100
200
% change when opposite species removed
Orange-crowned
warbler
Virginia’s
warbler
P = 0.77
P = 0.83
P = 0.02 P = 0.04
Figure 6.4
Percentage difference in feeding rates (mean ± SE) at orange-crowned
warbler and virginia’s warbler nests on plots where the other species had
been experimentally removed. Feeding rates (visits per hour to the nest with
food) were measured during incubation (inc; rates of male feeding of
incubating females on the nest) and during the nestling period (nstl; nestling
feeding rates by both parents combined). P-values are from t-tests of the
hypothesis that each species fed at higher rates on plots from which the other
had been removed. This hypothesis was supported for virginia’s warblers but
not orange-crowned warblers.
AFTER MARTIN & MARTIN, 2001
competition between groups
of unrelated species
9781405156585_4_006.qxd 11/5/07 14:51 Page 188