Townsend C.R., Begon M., Harper J.L. Essentials of Ecology
Подождите немного. Документ загружается.


had previously eaten between them – as did the rodents when the ants were
removed; only when both were removed did the amount of resource increase.
In other words, under normal circumstances both guilds eat less and achieve
lower levels of abundance than they would do if the other guild were absent. This
clearly indicates that rodents and ants, although they coexist in the same habitat,
compete interspecifically with one another.
6.2.7 The Competitive Exclusion Principle
The patterns that are apparent in these examples have also been uncovered in
many others, and have been elevated to the status of a principle: the Competitive
Exclusion Principle or Gause’s Principle (named after an eminent Russian ecologist).
It can be stated as follows:
l
If two competing species coexist in a stable environment, then they
do so as a result of niche differentiation, i.e. differentiation of their
realized niches.
l
If, however, there is no such differentiation, or if it is precluded by the
habitat, then one competing species will eliminate or exclude the other.
Although the principle has emerged here from a contemplation of patterns
evident in real sets of data, its establishment was – and many modern discussions
of interspecific competition still are – bound up with a simple mathematical model
of interspecific competition, usually known by the names of its two (independent)
originators: Lotka and Volterra (Box 6.1).
There is no question that there is some truth in the principle that competitor
species can coexist as a result of niche differentiation, and that one competitor
species may exclude another by denying it a realized niche. But it is crucial also
to be aware of what the Competitive Exclusion Principle does not say.
It does not say that whenever we see coexisting species with different niches
it is reasonable to jump to the conclusion that this is the principle in operation.
Each species, on close inspection, has its own unique niche. Niche differentiation
Chapter 6 Interspecific competition
189
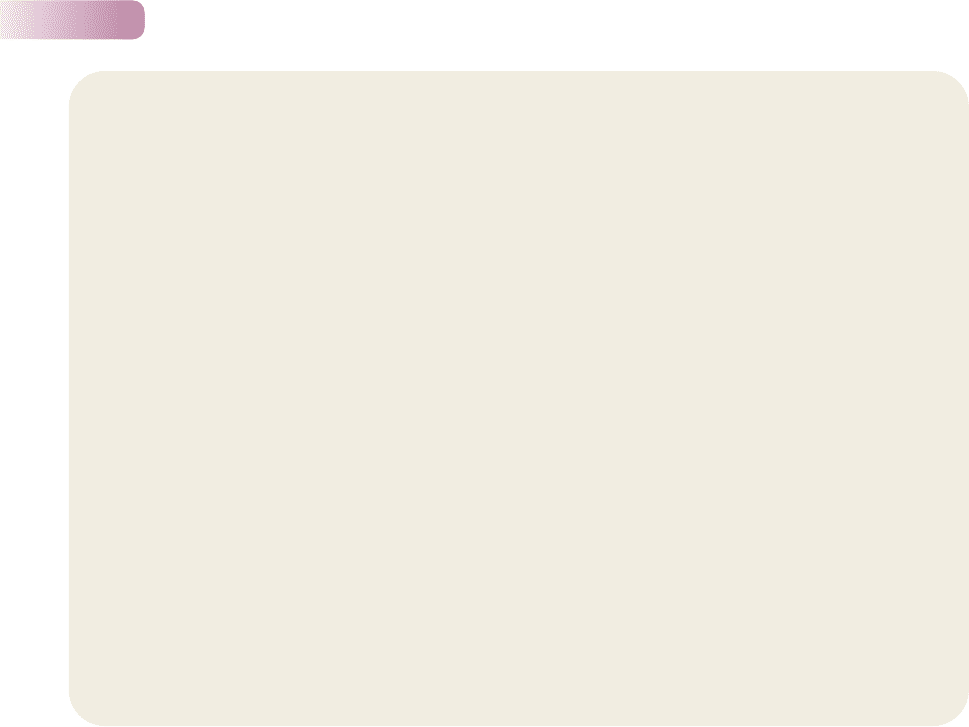
Figure 6.5
The diets of ants and rodents overlap:
sizes of seeds harvested by coexisting
ants and rodents near Portal, Arizona.
AFTER BROWN & DAVIDSON, 1977
0.3
0.2
0.1
0.0
0.3
0.2
0.1
0.0
<0.64
0.64–0.70
0.70–0.83
0.83–0.99
0.99–1.17
1.17–1.40
1.40–1.85
1.85–1.98
1.98–2.36
2.36–2.79
2.79–3.33
3.33–3.96
3.96–4.70
>4.70
Seed size (mm)
Frequency in diet
Rodents
Ants
The Competitive Exclusion
Principle – what it does and
does not say
9781405156585_4_006.qxd 11/5/07 14:51 Page 189

Part III Individuals, Populations, Communities and Ecosystems
190
(a) (b)
N
2
N
1
N
1
N
2
K
1
K
1
/α
21
K
1
α
12
K
2
6.1 QUANTITATIVE ASPECTS
6.1 Quantitative aspects
The most widely used model of interspecific competi-
tion is the Lotka–Volterra model (Volterra, 1926; Lotka,
1932). It is an extension of the logistic equation described
in Box 5.4. Its virtues are (like the logistic) its simplicity,
and its capacity to shed light on the factors that deter-
mine the outcome of a competitive interaction.
Within the logistic equation,
= rN
the particular term that models intraspecific competi-
tion is (K − N)/K. Within this term, the greater the value
of N (the bigger the population), the greater is the
strength of intraspecific competition. The basis of the
Lotka–Volterra model is the replacement of this term
by one that models both intra- and interspecific com-
petition. In the model, we call the population size of
the first species N
1
, and that of a second species N
2
.
Their carrying capacities and intrinsic rates of increase
are K
1
, K
2
, r
1
and r
2
.
By analogy with the logistic, we expect the total
competitive effect on, say, species 1 (intra- and inter-
specific) to be greater the larger the values of N
1
and
N
2
; but we cannot just add them together, since the
competitive effects of the two species on species 1
are unlikely to be the same. Suppose, though, that
10 individuals of species 2 have, between them, only
the same competitive effect on species 1 as does a
single individual of species 1. The total competitive
effect on species 1 will then be equivalent to the effect
of (N
1
+ N
2
*
1/10) species 1 individuals. The constant
(1/10 in the present case) is called a competition
coefficient and is denoted by α
12
(alpha one two).
Multiplying N
2
by α
12
converts it to a number of N
1
equivalents, and adding N
1
and α
12
N
2
together gives
us the total competitive effect on species 1. (Note
that α
12
< 1 means that individuals of species 2 have
less inhibitory effect on individuals of species 1 than
individuals of species 1 have on others of their own
species, and so on.)
(K − N)
K
dN
dt
The equation for species 1 can now be written:
= r
1
N
1
and for species 2 (with its own competition coefficient,
converting species 1 individuals into species 2
equivalents):
= r
2
N
2
These two equations constitute the Lotka–Volterra
model.
The best way to appreciate its properties is to ask
the question, ‘Under what circumstances does each
species increase or decrease in abundance?’ In order
to answer, it is necessary to construct diagrams in
which all possible combinations of N
1
and N
2
can be
displayed. This has been done in Figure 6.6. Certain
combinations (certain regions in Figure 6.6) give rise
to increases in species 1 and/or species 2, whereas
other combinations give rise to decreases. It follows
inevitably that there must also therefore be a so-called
(K
2
− [N
2
+ α
21
N
1
])
K
2
dN
2
dt
(K
1
− [N
1
+ α
12
N
2
])
K
1
dN
1
dt
The Lotka–Volterra model of interspecific competition
Figure 6.6
The zero isoclines generated by the Lotka–Volterra
competition equations. (a) The N
1
zero isocline: species 1
increases below and to the left of it, and decreases above
and to the right of it. (b) The equivalent N
2
isocline.
9781405156585_4_006.qxd 11/5/07 14:51 Page 190

Chapter 6 Interspecific competition
191
zero isocline for each species: that is, a line with com-
binations leading to increase on one side of it and
combinations leading to decrease on the other, but
along which there is neither increase nor decrease.
We can map out the regions of increase and
decrease in Figure 6.6 for species 1 if we can draw its
zero isocline, and we can do this by using the fact that
on the zero isocline, dN
1
/dt = 0 (the rate of change of
species 1 abundance is zero, by definition). Rearrang-
ing the equation, this gives us, as the zero isocline for
species 1:
N
1
= K
1
− α
21
N
2
Below and to the left of this, species 1 increases
in abundance (arrows in the figure, representing this
increase, point from left to right, since N
1
is on the
horizontal axis). It increases because numbers of
both species are relatively low, and species 1 is thus
subjected to only weak competition. Above and to the
right of the line, however, numbers are high, competi-
tion is strong and species 1 decreases in abundance
(arrows from right to left). Based on an equivalent
derivation, Figure 6.6b also shows the species 2 zero
isocline, with arrows, like the N
2
axis, running vertically.
In order to determine the outcome of competition
in this model, it is necessary to determine, at each
point on a figure, the behavior of the joint species
1–species 2 population, as indicated by the pair of
arrows. There are, in fact, four different ways in which
the two zero isoclines can be arranged relative to one
another, and these can be distinguished by the inter-
cepts of the zero isoclines (Figure 6.7). The outcome
of competition will be different in each case.
Looking at the intercepts in Figure 6.7a, for
instance,
> K
2
and K
1
>
Rearranging these slightly gives us:
K
1
> K
2
α
12
and K
1
α
21
> K
2
The first inequality (K
1
> K
2
α
12
) indicates that the
inhibitory intraspecific effects that species 1 can exert
on itself (denoted by K
1
) are greater than the inter-
specific effects that species 2 can exert on species 1
(K
2
α
12
). This means that species 2 is a weak inter-
specific competitor. The second inequality, however,
indicates that species 1 can exert more of an effect
K
2
α
21
K
1
α
12
s
N
2
N
1
N
1
K
1
K
1
K
2
/α
21
K
2
/α
21
K
1
α
12
K
2
K
2
K
1
α
12
N
2
N
2
N
2
N
1
N
1
K
1
K
1
K
2
/α
21
K
2
/α
21
K
1
α
12
K
2
K
2
K
1
α
12
(a) (b)
(c) (d)
Figure 6.7
The outcomes of competition generated by the
Lotka–Volterra competition equations for the four
possible arrangements of the N
1
and N
2
zero
isoclines. Black arrows refer to joint populations,
and are derived as indicated in (a). The solid circles
show stable equilibrium points. The open circle in
(d) is an unstable equilibrium point. For further
discussion, see box text.
9781405156585_4_006.qxd 11/5/07 14:51 Page 191
does not prove that there are coexisting competitors. The species may not be
competing at all and may never have done so in their evolutionary history. We
require proof of interspecific competition. In the examples above, this was provided
by experimental manipulation – remove one species (or one group of species)
and the other species increases its abundance or its survival. But most of even the
more plausible cases for competitors coexisting as a result of niche differentia-
tion have not been subjected to experimental proof. So just how important is
the Competitive Exclusion Principle in practice? We return to this question in
Section 6.5.
Part of the problem is that although species may not be competing now, their
ancestors may have competed in the past, so that the mark of interspecific com-
petition is left imprinted on the niches, the behavior or the morphology of their
present-day descendants. This particular question is taken up in Section 6.3.
Finally, the Competitive Exclusion Principle, as stated above, includes the
word ‘stable’. That is, in the habitats envisaged in the principle, conditions and
the supply of resources remain more or less constant – if species compete, then
that competition runs its course, either until one of the species is eliminated
or until the species settle into a pattern of coexistence within their realized
niches. Sometimes this is a realistic view of a habitat, especially in laboratory
Part III Individuals, Populations, Communities and Ecosystems
192
on species 2 than species 2 can on itself. Species 1
is thus a strong interspecific competitor; and as the
arrows in Figure 6.7a show, species 1 drives the weak
species 2 to extinction and attains its own carrying
capacity. The situation is reversed in Figure 6.7b.
Hence Figure 6.7a and b describe cases in which
the environment is such that one species invariably
outcompetes the other, because the first is a strong
interspecific competitor and the other weak.
In Figure 6.7c, by contrast:
K
1
> K
2
α
12
and K
2
> K
1
α
21
In this case, both species have less competitive
effect on the other species than those other species
have on themselves; in this sense, both are weak
competitors. This would happen, for example, if there
were niche differentiation between the species –
each competed mostly ‘within’ its own niche. The
outcome, as Figure 6.7c shows, is that all arrows point
towards a stable, equilibrium combination of the
two species, which all joint populations therefore tend
to approach: that is, the outcome of this type of com-
petition is the stable coexistence of the competitors.
Indeed, it is only this type of competition (both species
having more effect on themselves than on the other
species) that does lead to the stable coexistence of
competitors.
Finally, in Figure 6.7d:
K
2
α
12
> K
1
and K
1
α
21
> K
2
Thus individuals of both species have a greater com-
petitive effect on individuals of the other species than
those other species do on themselves. This will occur,
for instance, when each species is more aggressive
toward individuals of the other species than toward
individuals of its own species. The directions of the
arrows are rather more complicated in this case, but
eventually they always lead to one or other of two
alternative stable points. At the first, species 1 reaches
its carrying capacity with species 2 extinct; at the
second, species 2 reaches its carrying capacity with
species 1 extinct. In other words, both species are
capable of driving the other species to extinction,
but which actually does so cannot be predicted with
certainty. It depends on which species has the upper
hand in terms of densities, either because they start
with a higher density or because density fluctua-
tions in some other way give them that advantage.
Whichever species has this upper hand, capitalizes
on that and drives the other species to extinction.
s
9781405156585_4_006.qxd 11/5/07 14:51 Page 192

or other controlled environments where the experimenter holds conditions and
the supply of resources constant. However, most environments are not stable
for long periods of time. How does the outcome of competition change when
environmental heterogeneity in space and time are taken into consideration? This
is the subject of the next section.
6.2.8 Environmental heterogeneity
As explained in previous chapters, spatial and temporal variations in environ-
ments are the norm rather than the exception. Environments are usually a
patchwork of favorable and unfavorable habitats; patches are often only available
temporarily; and patches often appear at unpredictable times and in unpredict-
able places. Under such variable conditions, competition may only rarely ‘run
its course’, and the outcome cannot be predicted simply by application of the
Competitive Exclusion Principle. A species that is a ‘weak’ competitor in a con-
stant environment might, for example, be good at colonizing open gaps created
in a habitat by fire, or a storm, or the hoofprint of a cow in the mud – or may be
good at growing rapidly in such gaps immediately after they are colonized. It
may then coexist with a strong competitor, as long as new gaps occur frequently
enough. Thus, a realistic view of interspecific competition must acknowledge that
it often proceeds not in isolation, but under the influence of, and within the
constraints of, a patchy, impermanent or unpredictable world.
The following examples illustrate just two of the many ways in which environ-
mental heterogeneity ensures that the Competitive Exclusion Principle is very far
from being the whole story when it comes to determining the outcome of an
interaction between competing species.
The first concerns the coexistence of a superior competitor and a superior
colonizer: the sea palm Postelsia palmaeformis (a brown alga) and the mussel
Mytilus californianus on the coast of Washington, USA (Paine, 1979) (Figure 6.8).
Chapter 6 Interspecific competition
193
Time
Low
disturbance
shore
Regularly
disturbed
shore
Figure 6.8
On shores in which gaps are not created, mussels are able to exclude the brown alga Postelsia; but where gaps are created regularly enough the
two species coexist, even though Postelsia is eventually excluded by the mussels from each gap.
competition may only rarely
‘run its course’
mussels, sea palms and the
frequency of gap formation
9781405156585_4_006.qxd 11/5/07 14:51 Page 193

Postelsia is an annual plant that must re-establish itself each year in order to persist
at a site. It does so by attaching to the bare rock, usually in gaps in the mussel bed
created by wave action. However, the mussels themselves slowly encroach on
these gaps, gradually filling them and precluding colonization by Postelsia. In
other words, in a stable environment, the mussels would outcompete and exclude
Postelsia. But their environment is not stable – gaps are frequently being created.
It turns out that these species coexist only at sites in which there is a relatively
high average rate of gap formation (at least 7% of surface area per year), and in
which this rate is approximately the same each year. Where the average rate is
lower, or where it varies considerably from year to year, there is (either regularly
or occasionally) a lack of bare rock for colonization by Postelsia. At the sites of
coexistence, on the other hand, although Postelsia is eventually excluded from
each gap, these are created with sufficient frequency and regularity for there to
be coexistence in the site as a whole. In short, there is coexistence of competitors
– but not as a result of niche differentiation.
A perhaps more widespread path to the coexistence of a superior and an
inferior competitor is based on the idea that the two species may have independ-
ent, aggregated (i.e. clumped) distributions over the available habitat. This would
mean that the powers of the superior competitor were mostly directed against
members of its own species (in the high-density clumps), but that this aggregated
superior competitor would be absent from many areas – within which the inferior
competitor could escape competition. An inferior competitor may then be able
to coexist with a superior competitor that would rapidly exclude it from a con-
tinuous, homogeneous environment.
That such aggregated distributions are indeed a reality is illustrated by a field
study of two species of sand-dune plant, Aira praecox and Erodium cicutarium,
in northwest England. Both species were aggregated, and the smaller plant, Aira,
tended to be aggregated even at the smallest spatial scales (Figure 6.9a). The two
species, though, were negatively associated with one another at these smallest
scales (Figure 6.9b). Thus, Aira tended to occur in small single-species clumps
and was therefore much less liable to competition from Erodium than would have
been the case if they had been distributed at random.
The consequences of such aggregated distributions are illustrated by a
study of experimental communities of four annual terrestrial plants – Capsella
Part III Individuals, Populations, Communities and Ecosystems
194
Seashore with Postelsia
and Mytilus californianus.
coexistence as a result of
aggregated distributions
© GERALD AND BUFF CORSI, VISUALS UNLIMITED
9781405156585_4_006.qxd 11/5/07 14:51 Page 194
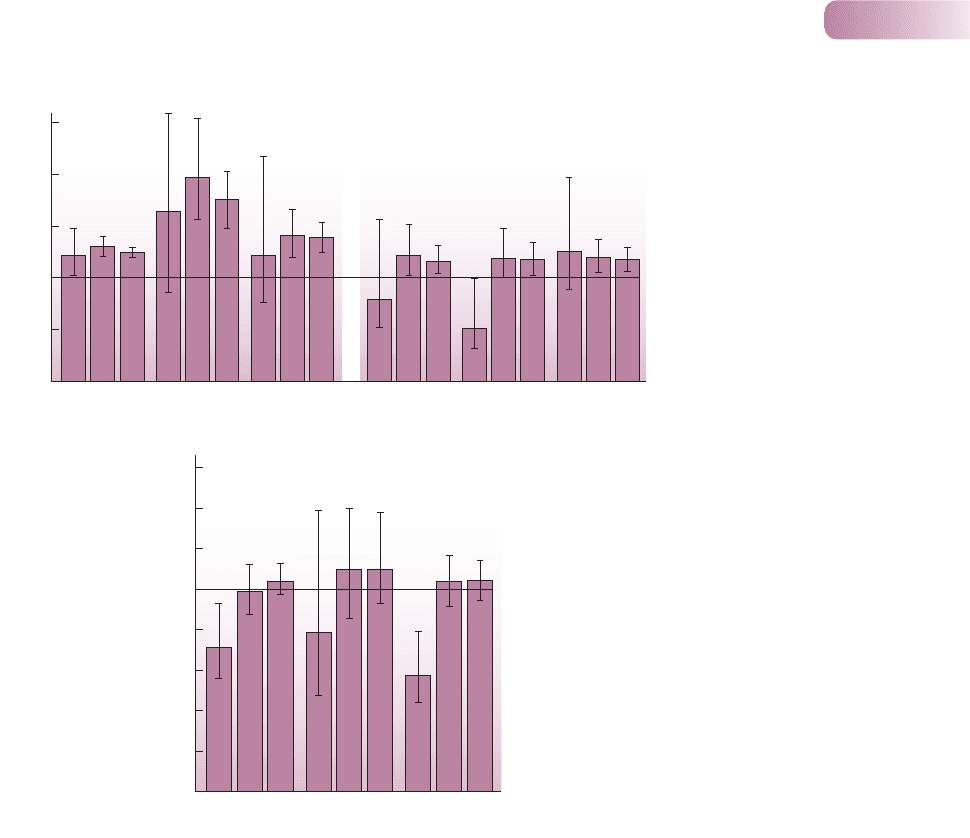
bursa-pastoris, Cardamine hirsuta, Poa annua and Stellaria media (Figure 6.10).
Stellaria is known to be the superior competitor among these species. Replicate
three- and four-species mixtures were sown at high density, and the seeds were
either placed completely at random, or seeds of each species were aggregated
in subplots within the experimental areas. Intraspecific aggregation harmed the
performance of the superior Stellaria in the mixtures, whereas in all but one
case aggregation improved the performance of the three inferior competitors.
Again, coexistence of competitors was favored not by niche differentiation but
simply by a type of heterogeneity that is typical of the natural world: aggregation
ensured that most individuals competed with members of their own and not of
another species.
These studies, and others like them, go a long way toward explaining the
co-occurrence of species that in constant, homogeneous environments would
probably exclude one another. The environment is rarely unvarying enough for
competitive exclusion to run its course or for the outcome to be the same across
the landscape.
Chapter 6 Interspecific competition
195
(b)
Association index
10 30 50 10 30 50 10 30 50
1995 1996 1997
Radius (mm)
(a)
Aggregation index
10
0.0
1.0
2.0
0.5
1.5
2.5
30 50 10 30 50 10 30 50
1995 1996 1997
Aira
Radius (mm)
10 30 50 10 30 50 10 30 50
1995 1996 1997
Erodium
1.6
1.4
1.2
1.0
0.8
0.6
0.4
0.2
0.0
Figure 6.9
(a) Spatial distribution of two
sand-dune species, Aira praecox
and Erodium cicutarium at a site in
northwest England. An aggregation
index of 1 indicates a random
distribution. Indices greater than
1 indicate aggregation (clumping)
within patches with the radius as
specified; values less than 1 indicate
a regular distribution. Bars represent
95% confidence intervals. (b) The
association between Aria and
Erodium in each of the 3 years.
An association index greater than 1
would indicate that the two species
tended to be found together more
than would be expected by chance
alone in patches with the radius as
specified; values less than 1 indicate
a tendency to find one species or
the other. Bars represent 95%
confidence intervals.
AFTER COOMES ET AL., 2002
9781405156585_4_006.qxd 11/5/07 14:51 Page 195
Part III Individuals, Populations, Communities and Ecosystems
196
0
900
600
300
(a)
Capsella bursa-pastoris
Random
Aggregated
0
100
50
(b)
Cardamine hirsuta
0
300
200
100
(c)
Poa annua
0
2000
1000
Mixtures
(d) Stellaria media
Above-ground biomass (g m
–2
)
Cbp
Ch
Pa
Cbp
Ch
Sm
Cbp
Pa
Sm
Ch
Pa
Sm
Cbp
Ch
Pa
Sm
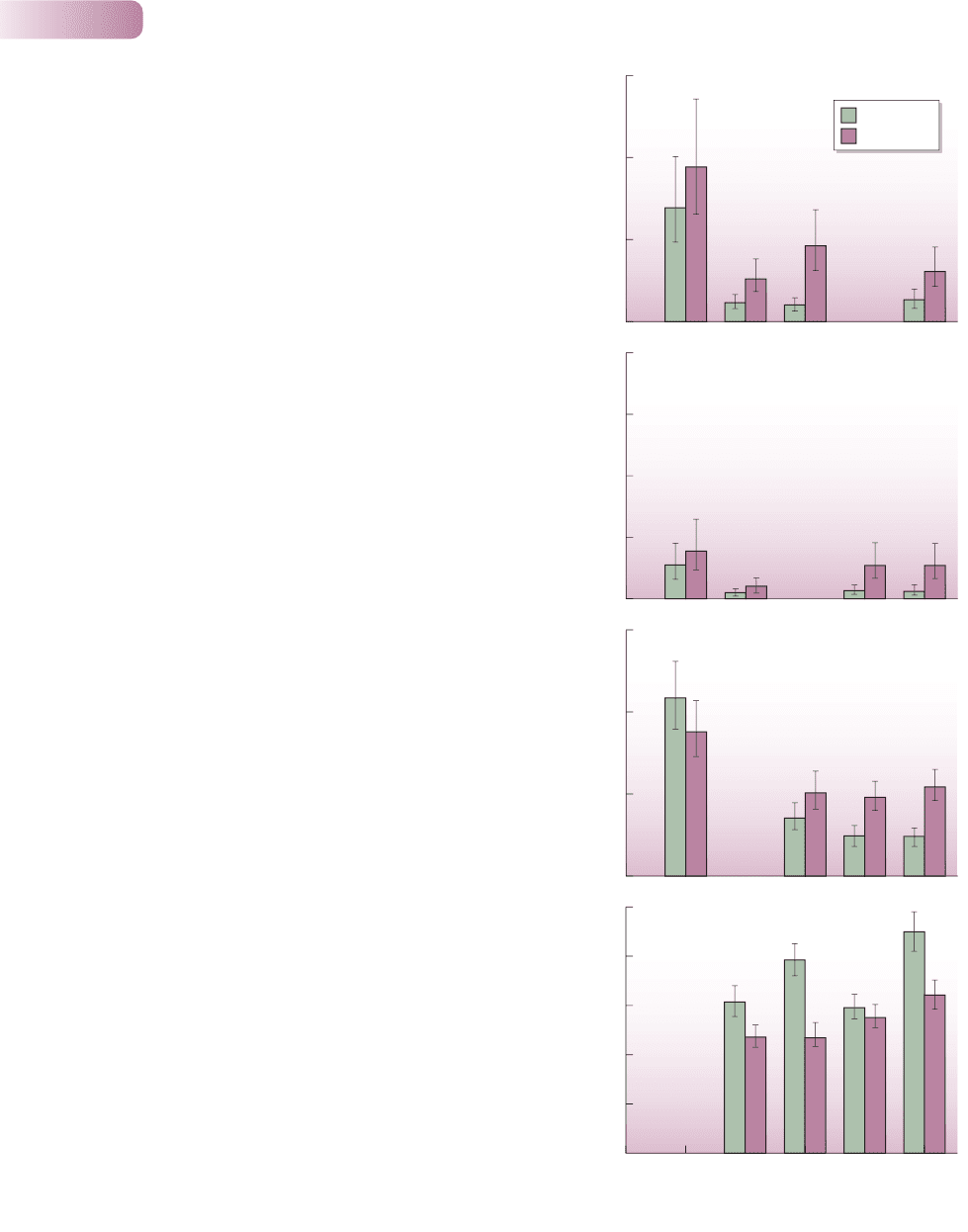
Figure 6.10
The effect of intraspecific aggregation on above-ground biomass
(mean ± SE) of four plant species grown for 6 weeks in three- and
four-species mixtures (four replicates of each). The normally competitively
superior Stellaria media (Sm) did consistently less well when seeds were
aggregated than when they were placed at random (d). In contrast, the
three competitively inferior species – Capsella bursa-pastoris (Cbp),
Cardamine hirsuta (Ch) and Poa annua (Pa) – almost always performed
better when seeds had been aggregated (a–c). Note the different scales
on the vertical axes, and that the compositions of the mixtures are given
only along the horizontal axis of (d).
AFTER STOLL & PRATI, 2001
9781405156585_4_006.qxd 11/5/07 14:51 Page 196
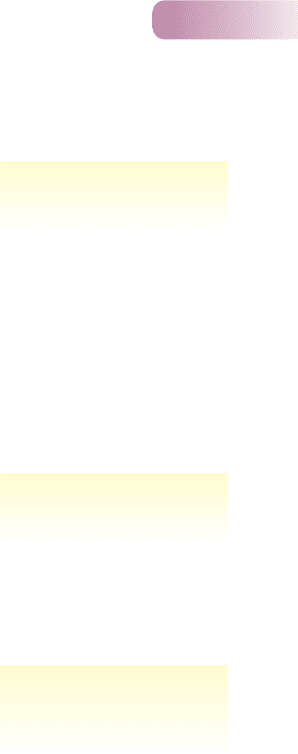
6.3 Evolutionary effects of interspecific
competition
Putting to one side the fact that environmental heterogeneity ensures that the
forces of interspecific competition are often much less profound than they would
otherwise be, it is nonetheless the case that the potential of interspecific com-
petition to adversely affect individuals is considerable. We have seen in Chapter 2
that natural selection in the past will have favored those individuals that, by their
behavior, physiology or morphology, have avoided adverse effects that act on
other individuals in the same population. The adverse effects of extreme cold, for
example, may have favored individuals with an enzyme capable of functioning
effectively at low temperatures. Similarly, in the present context, the adverse
effects of interspecific competition may have favored those individuals that
managed to avoid those competitive effects. We can, therefore, expect species to
have evolved characteristics that ensure that they compete less, or not at all, with
members of other species.
How will this look to us at the present time? Coexisting species, with an appar-
ent potential to compete, will exhibit differences in behavior, physiology or
morphology that ensure that they compete little or not at all. Connell has called
this line of reasoning ‘invoking the ghost of competition past’. Yet the pattern it
predicts is precisely the same as that supposed by the Competitive Exclusion
Principle to be a prerequisite for the coexistence of species that still compete.
Coexisting present-day competitors, and coexisting species that have evolved an
avoidance of competition, can look the same.
The question of how important either past or present competition are as
forces structuring natural communities will be addressed in the last section of this
chapter (Section 6.5). For now, we examine some examples of what interspecific
competition can do as an evolutionary force. Note, however, that by invoking
something that cannot be observed directly (evolution), it may be impossible
to prove an evolutionary effect of interspecific competition, in the strict sense
of ‘proof’ that can be applied to mathematical theorems or carefully controlled
experiments in the laboratory. Nonetheless, we consider some examples where
an evolutionary (rather than an ecological) effect of interspecific competition is
the most reasonable explanation for what is observed.
6.3.1 Character displacement and ecological release in
the Indian mongoose
In western parts of its range, the small Indian mongoose (Herpestes javanicus)
coexists with one or two slightly larger species in the same genus (H. edwardsii
and H. smithii), but these species are absent in the eastern part of its range
(Figure 6.11a). The upper canine teeth are the mongoose’s principal prey-killing
organ, and these vary in size within and between species and between the sexes
(female mongooses are smaller than males). In the east, where H. javanicus occurs
alone (area VII in Figure 6.11a), both males and females have larger canines
than in the western areas (III, V, VI) where it coexists with the larger species
(Figure 6.11b). This is consistent with the view that where similar but larger
mongoose species are present, the prey-catching apparatus of H. javanicus has
Chapter 6 Interspecific competition
197
evolutionary avoidance of
competition
invoking the ghost of
competition past
the difficulty of distinguishing
ecological and evolutionary
effects
9781405156585_4_006.qxd 11/5/07 14:51 Page 197
been selected for reduced size (referred to as ‘character displacement’), reducing
the strength of competition with other species in the genus because smaller
predators tend to take smaller prey. Where H. javanicus occurs in isolation,
since no character displacement has occurred, its canine teeth are much larger.
(Another strong candidate for the evolutionary effects of interspecific competi-
tion, especially because of its association with character displacement, is provided
by Darwin’s finches of the genus Geospiza living on the Galapagos islands, dis-
cussed in Section 2.4.2.)
In fact, H. javanicus was introduced about a century ago to many islands
outside its native range (often as part of a naive attempt to control introduced
rodents). In these places, the larger competitor mongoose species were absent.
Within 100–200 generations H. javanicus had increased in size (Figure 6.11b),
so that the sizes of island individuals are now intermediate between those in the
region of origin (where they coexisted with other species and were small) and
those in the east where they occur alone. Their size on the islands is consistent
with ‘ecological release’ from competition with larger species.
Part III Individuals, Populations, Communities and Ecosystems
198
(a)
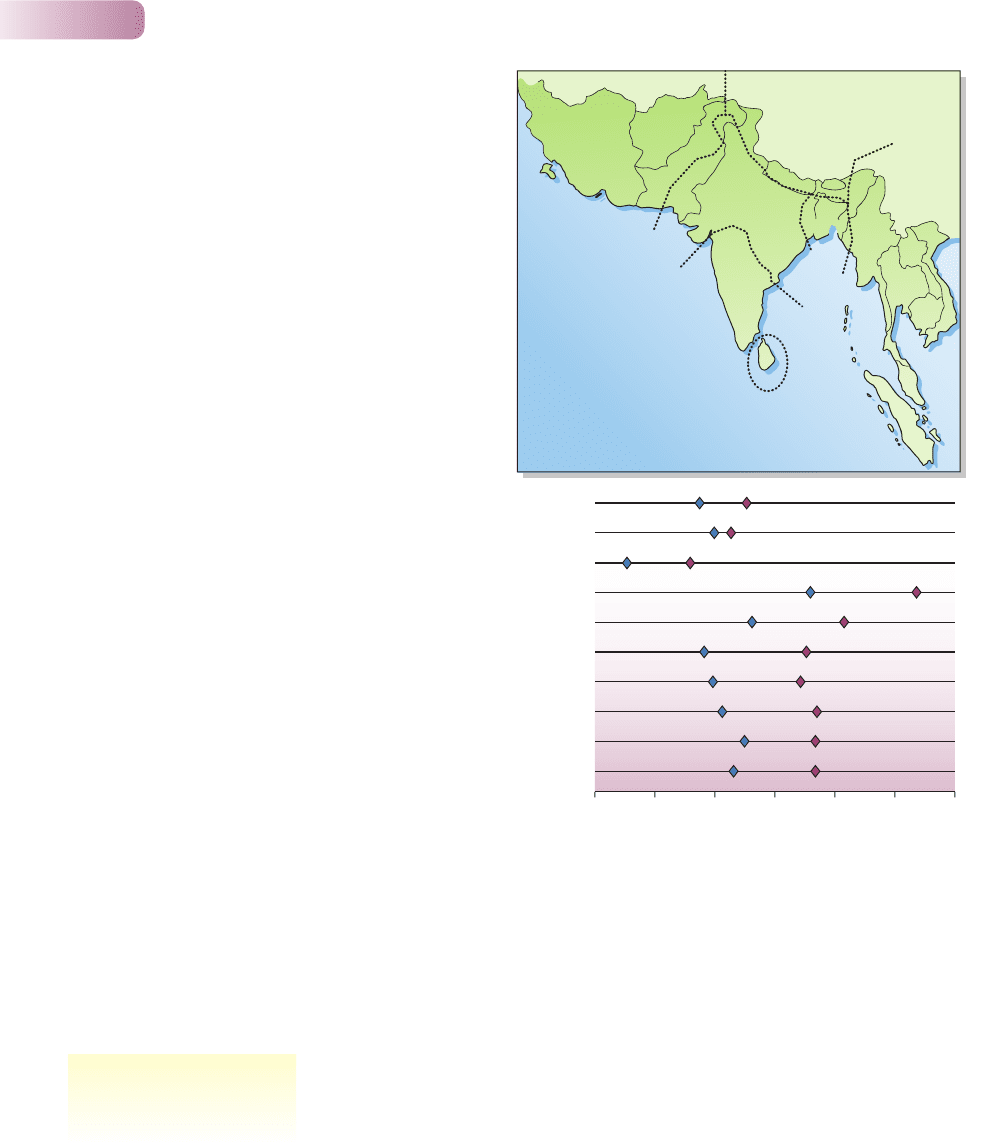
(b) Asia III
Asia VI
St Croix
Hawaii
Oahu
Mauritius
2.25 2.50 2.75 3.00 3.25 3.50 3.75
Asia V
Asia VII
Viti Levu
Okinawa
C
sup
L (mm)
IV
(e, j)
III
(e, j, s)
V
(e, j)
II
(e, s)
I
(e, s)
VI
(e, j, s)
VII
(j)
Figure 6.11
(a) Native geographic ranges of Herpestes javanicus (j),
H. edwardsii (e) and H. smithii (s). (b) Maximum diameter (mm)
of the upper canine (C
sup
L) for Herpestes javanicus in its native
range [data only for areas III, V, VI and VII from (a)] and islands on
which it has been introduced. Symbols in blue represent mean
female size and in maroon mean male size. Compared to area VII
(H. javanicus alone), animals in areas III, V and VI, where they
compete with the two larger species, are smaller. On the islands,
they have increased in size since their introduction, but are still not
as large as in area VII.
FROM SIMBERLOFF ET AL., 2000
smaller teeth in the small Indian
mongoose when larger
competitors are present
9781405156585_4_006.qxd 11/5/07 14:51 Page 198