Townsend C.R., Begon M., Harper J.L. Essentials of Ecology
Подождите немного. Документ загружается.

Chapter 8 Evolutionary ecology
279
Molecular ecology: differentiation within and
between species
For much of the time, it is entirely appropriate for eco-
logists to talk about ‘populations’ or ‘species’ as if they
were singular, homogeneous entities, but for some
purposes, knowing how much differentiation there is
within species, or between one species and another,
is critical for an understanding of their dynamics, and
ultimately for managing those dynamics. Molecular
genetic markers, of a variety of types, have massively
increased the resolution at which we can differentiate
between populations and even individuals.
Studies on albatrosses illustrate how even within a
species of conservation importance, separate popula-
tions even more threatened with extinction may be
hidden; while studies on salmon illustrate how mole-
cular markers can be used to detect, and to prosecute,
illegal fishermen. Molecular markers have also shown,
for example, that a threatened ‘species’, the red wolf,
may in fact be a hybrid between two other, relatively
common species, with implications for both the desir-
ability and the practicality of its conservation.
Coevolutionary arms races
A better defended food resource exerts a selection
pressure on consumers to overcome that defense. A
consumer that does so will steal a march on its com-
petitors, and so is likely to become relatively specialized
on that prey type – which is then under particular pres-
sure to defend itself against that particular consumer,
and so on: a coevolutionary ‘arms race’. Plants relying
on toxins are more prone to becoming involved in
arms races with their herbivores than those relying on
more ‘quantitative’ (digestion-reducing) chemicals.
The intimate association between parasites and
their hosts makes them especially prone to coevolu-
tionary arms races. However, the process is not
necessarily so straightforward, as illustrated by the
case of the myxoma virus and the European rabbit.
The evolution of resistance in the rabbit is easy to
understand, but the parasites favored by natural
selection are those with the greatest reproductive rate.
In the myxoma virus, this occurs at intermediate levels
of virulence because of increased transmissibility.
In other cases, host–parasite coevolution is more
definitely antagonistic: increasing resistance in the
host and increasing infectivity in the parasite. With
bacteria and their viruses, this process can be observed
in action, because generation times are so short.
Mutualistic interactions
No species lives in isolation, but often the association
with other species is especially close: for many organ-
isms, the habitat they occupy is an individual of another
species – a symbiosis. A mutualistic relationship is
one in which organisms of different species interact
to their mutual benefit. Current evolutionary thinking
views mutualisms as cases of reciprocal exploitation
where nonetheless each partner is a net beneficiary.
Mutualisms themselves have often been neglected in
the past compared to other types of interaction, yet
mutualists compose most of the world’s biomass.
Pairs of species from many taxa take part in mutual-
istic associations in which one species protects the other
from predators or competitors but gains privileged
access to a food resource on the protected species.
Some of the most dramatic mutualisms are those
of human agriculture, but similar ‘farming’ mutualisms
have developed in termite and especially ant societies.
Ants farm many species of aphids in return for sugar-
rich secretions of honeydew. The aphids benefit through
suffering lower mortality rates; but there are also costs:
where aphid predators are excluded experimentally,
aphids grow less well in the presence of ants.
Very many plant species use animals to disperse
their seeds and pollen, and many different kinds of
animals have entered into pollination liaisons with
flowering plants. The pollinators par excellence, though,
are the insects.
The gastrointestinal tracts of all vertebrates are
populated by a mutualistic microbiota. The microbes
receive a steady flow of substrates for growth in the
form of food that has been eaten, and they live within a
chamber in which pH and, in endotherms, temperature
SUMMARY
Summary
s
9781405156585_4_008.qxd 11/5/07 14:55 Page 279

Part III Individuals, Populations, Communities and Ecosystems
280
are regulated and anaerobic conditions are maintained.
The vertebrate hosts receive nutrition from food that
they would otherwise find, literally, indigestible.
Most higher plants do not have roots, they have
mycorrhizas – intimate mutualisms between fungi and
root tissue. In ectomycorrhizas (ECMs), fungi form a
sheath of varying thickness around the roots. These
fungi are effective in extracting the sparse and patchy
supplies of phosphorus and especially nitrogen from
the forest litter layer. Carbon flows from the plant to the
fungus (mostly hexose sugars). However, ECM growth
is directly related to the rate of flow of the sugars from
the plant. When the direct availability of nitrate to the
plants is high, plant metabolism is directed away from
hexose production. As a result the ECM degrades:
the plants seem to support just as much ECM as
they appear to need. Arbuscular mycorrhizas (AMs)
penetrate within the roots of the host. There has been
a tendency to emphasize facilitation of the uptake of
phosphorus as the main benefit to plants from AM
symbioses, but benefits have been demonstrated,
too, in nitrogen uptake, pathogen and herbivore
protection, and resistance to toxic metals.
The ability to fix nitrogen is widely distributed
amongst both the eubacteria and the Archaebacteria,
and many of these have been caught up in tight mutu-
alisms with distinct groups of eukaryotes. The best
known are the rhizobia, which fix nitrogen in the root
nodules of most leguminous plants. Nitrogen fixation is
often energetically inefficient, but energy may be much
more readily available for green plants than nitrogen.
On the other hand, when a nodulated legume is pro-
vided with nitrates, nitrogen fixation declines rapidly.
The mutualisms of rhizobia and legumes (like other
nitrogen-fixing mutualisms) must be seen in the con-
text of competition between legumes and non-legumes.
s
REVIEW QUESTIONS
Review questions
Asterisks indicate challenge questions
1 Explain why molecular (DNA) markers have
improved the ability of ecologists to study
degrees of differentiation within and between
species.
2* Review the range of molecular markers that
have been used in molecular ecology, stressing
their advantages and disadvantages at different
scales of resolution.
3 Should the red wolf be conserved, or would that
be a misguided waste of public money?
4 Why are some plants more likely than others
to be involved in arms races with their insect
herbivores?
5 Account for the decline in virulence of the
myxomatosis virus in European rabbits
after its initial introductions in Australia and
Europe.
6 Compare and contrast the mutualistic
associations of ants with plants they protect
and aphids they farm.
7* Discuss the following propositions: ‘Most
herbivores are not really herbivores but
consumers of the byproducts of the mutualists
living in their gut’ and ‘Most gut parasites are
not really parasites but competitors with their
hosts for food that the host has captured’.
8 Compare the roles of fruits and nectar in the
interactions between plants and the animals
that visit them.
9 What are mycorrhizas and what is their
significance?
10* Leguminous plants are a perfect example
of a mutualistic association that can only be
understood in the context of the ecological
community within which it normally exists.
Discuss.
9781405156585_4_008.qxd 11/5/07 14:55 Page 280

281
Chapter 9
From populations to
communities
CHAPTER CONTENTS
9.1 Introduction
9.2 Multiple determinants of the dynamics of populations
9.3 Dispersal, patches and metapopulation dynamics
9.4 Temporal patterns in community composition
9.5 Food webs
Chapter contents
KEY CONCEPTS
In this chapter you will:
l
appreciate the variety of interacting abiotic and biotic factors that
account for the dynamics of populations
l
distinguish between the determination and regulation of population
abundance
l
understand how patchiness and dispersal between patches influence
the dynamics of both populations and communities
l
recognize the influence of disturbance on community patterns and
understand the nature of community succession
l
appreciate the importance of direct and indirect effects and distinguish
between bottom-up and top-down control of food webs
l
understand the relationship between the structure and stability of
food webs
Key concepts
9781405156585_4_009.qxd 11/5/07 14:56 Page 281

9.1 Introduction
Single-species populations have been the focus for many of the questions posed
in previous chapters. In attempting to answer the most fundamental ecological
question of all – what determines a species’ abundance and distribution – we have
chosen to ask separately about the role of conditions and resources, of migration,
of competition (both intra- and interspecific), of mutualism, and of predation and
parasitism. In reality, the dynamics of any population reflect a combination of these
effects, though their relative importance varies from case to case. Now, therefore,
we need to view the population in the context of the whole community, since
each exists within a whole web of interactions (Figure 9.1), and each responds
differently to the prevailing abiotic conditions.
In Section 9.2 we consider how abiotic and biotic factors combine to deter-
mine the dynamics of species populations. Then, in Section 9.3, we revisit one
of the major themes of this book – the importance of patchiness and dispersal
between patches in ecological dynamics – and discuss especially the importance
of the concept of the metapopulation. Disturbances, such as forest fires and the
storm battering of seashores, also play an important role in the dynamics of many
populations and the composition of most communities. After each disturbance,
there is a pattern of re-establishment of species that is played out against a
background of changing conditions, resources and population interactions. We
deal with temporal patterns in community composition, including community
succession, in Section 9.4. Finally, in Section 9.5 we broaden our view further
to examine food webs, like the one illustrated in Figure 9.1, with usually at least
Part III Individuals, Populations, Communities and Ecosystems
282
In previous chapters, we generally dealt with individual species or pairs of species
in isolation, as ecologists often do. Ultimately, however, we must recognize that
every population exists within a web of interactions with myriad other
populations, across several trophic levels. Each population must be viewed
in the context of the whole community, and we need to understand that
populations occur in patchy and inconstant environments in which
disturbance and local extinction may be common.
6
2
7
54
31
Figure 9.1
Community matrix illustrating how each
species may interact with several others in
competitive interactions (among plant species
1, 2 and 3; or between grazers 4 and 5; or
between predators 6 and 7) and predator–prey
interactions (such as between 6 and 4, or 5
and 2).
9781405156585_4_009.qxd 11/5/07 14:56 Page 282

three trophic levels (plant–herbivore–predator), emphasizing the importance
not only of direct but also of indirect effects that a species may have on others
on the same trophic level or on levels below or above it.
9.2 Multiple determinants of the dynamics
of populations
Why are some species rare and others common? Why does a species occur at
low population densities in some places and at high densities in others? What
factors cause fluctuations in a species’ abundance? These are crucial questions
when we wish to conserve rare species, or control pests, or manage natural, living
resources, or when we wish simply to understand the patterns and dynamics of
the natural world. To provide complete answers for even a single species in
a single location, we need to know the physicochemical conditions, the level
of resources available, the organism’s life cycle and the influence of competitors,
predators, parasites and so on – and how all these factors influence abundance
through effects on birth, death, dispersal and migration. We now bring these
factors together and consider how we might discover which actually matter in
particular examples.
The raw material for the study of abundance is usually some estimate of the
numbers of individuals in a population. However, a record of numbers alone
can hide vital information. Picture three human populations, shown to contain
identical numbers of individuals. One is an old people’s residential area, the
second is a population of young children and the third is a population of mixed
age and sex. In the absence of information beyond mere numbers, it would not
be clear that the first population was doomed to extinction (unless maintained
by immigration), the second would grow fast but only after a delay and the
third would continue to grow steadily. The most satisfactory studies, therefore,
estimate not only the numbers of individuals (and their parts, in the case of
modular organisms) but also those of different age, sex and size.
The data that accumulate from estimates of abundance may be used to estab-
lish correlations with external factors like food or weather. Correlations may
be used to predict the future. For example, high intensities of the disease ‘late
blight’ in potato crops usually occur 15–22 days after a period in which the
minimum temperature is above 10°C and relative humidity is more than 75% for
two consecutive days. Such a correlation may alert the potato grower to the need
for protective spraying. Correlations may also suggest – but not prove – causal
relationships. For example, a correlation may be demonstrated between the size
of a population and its growth rate. But ultimately ‘cause’ requires a mechanism.
When the population is large, many individuals may starve to death, or may fail
to reproduce, or may become aggressive and drive out the weaker members.
A correlation cannot tell us which. Nonetheless, correlations can be informative.
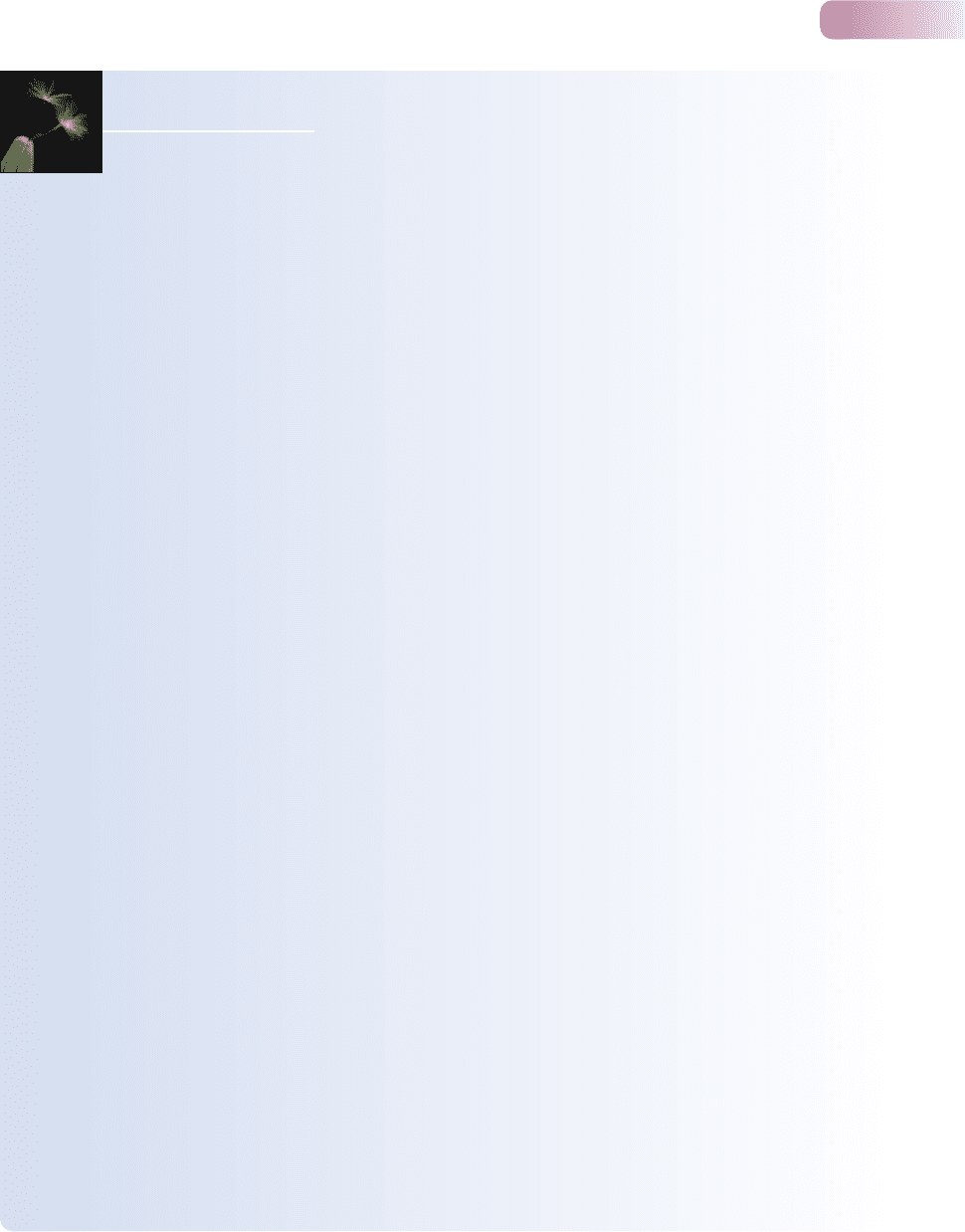
Figure 9.2, for example, shows four examples in which population growth rate
increases with the availability of food. It also suggests that in general, such rela-
tionships are likely to level off at the highest food levels where some other factor
or factors place an upper limit on abundance.
Chapter 9 From populations to communities
283
fluctuations in abundance are
caused by a wide variety of
biotic and abiotic factors
what total numbers can and
cannot tell us
what correlations can and
cannot tell us
9781405156585_4_009.qxd 11/5/07 14:56 Page 283
9.2.1 Fluctuation or stability?
Some populations appear to change very little in size. One study that covered an
extended timespan – though it was not necessarily the most scientific – examined
swifts (Micropus apus) in the village of Selborne in southern England over more
than 200 years. In one of the earliest published works on ecology, Gilbert White,
who lived in the village, wrote in 1778 (see White, 1789):
I am now confirmed in the opinion that we have every year the same number of
pairs invariably. ...The number that I constantly find are eight pairs, about half
of which reside in the church, and the rest in some of the lowest and meanest
thatched cottages.
More than 200 years later, Lawton and May (1984) visited the village and,
not surprisingly, found major changes. Swifts are unlikely to have nested in the
church for 50 years, and the thatched cottages have either disappeared or had
their roofs covered with wire. Yet the number of breeding pairs of swifts regularly
to be found in the village is now 12. In view of the many changes that have taken
place in the intervening centuries, this number is remarkably close to the eight
pairs so consistently found by White.
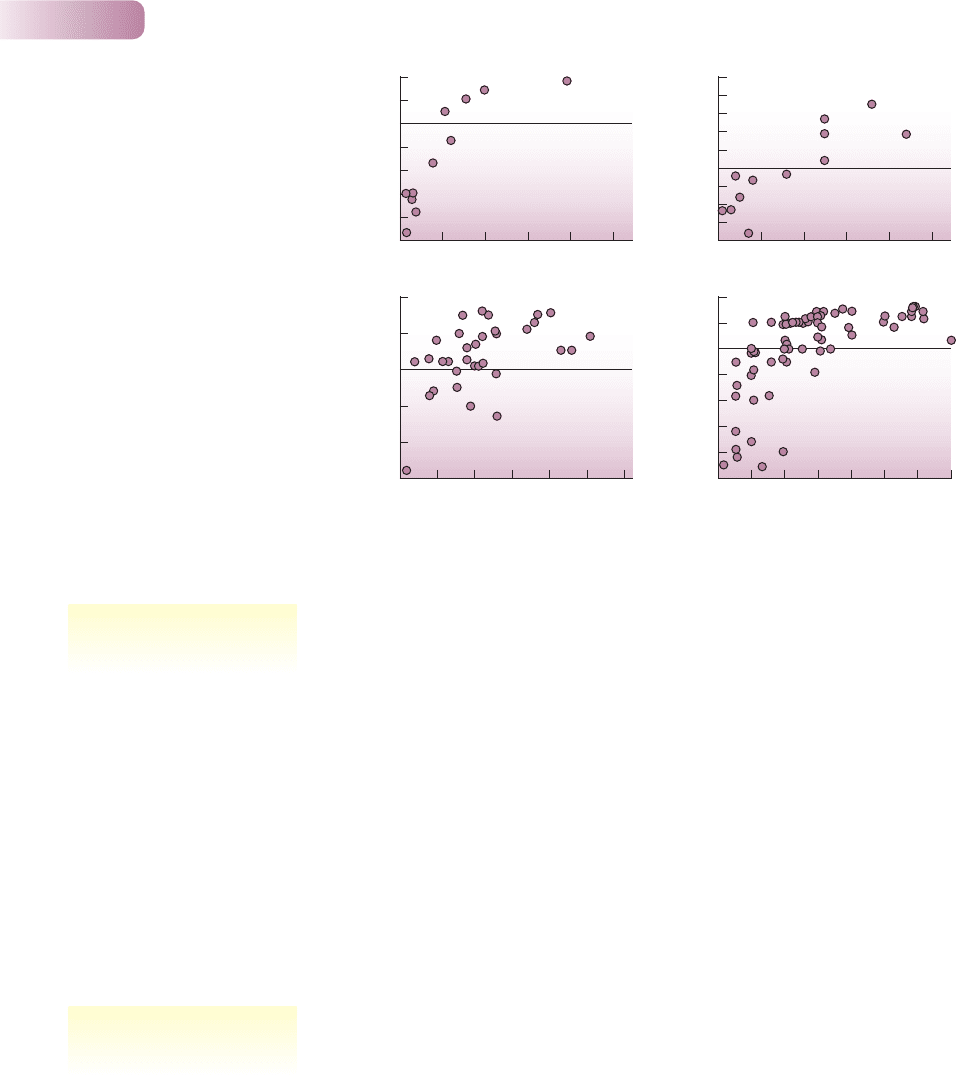
But the stability of a population may conceal complex underlying dynamics.
Another example of a population showing relatively little change in adult num-
bers from year to year is seen in an 8-year study in Poland of the small, annual
sand-dune plant Androsace septentrionalis (Figure 9.3a). Each year, however,
there was great flux within the population. Between 150 and 1000 new seedlings
per square meter appeared, but subsequent mortality reduced the population
by between 30% and 70%. Thus, the population appears to be kept within
bounds. At least 50 plants always survived to fruit and produce seeds for the next
season. By contrast, the mice in Figure 9.3b have extended periods of relatively
low abundance interrupted by sporadic and dramatic irruptions.
Part III Individuals, Populations, Communities and Ecosystems
284
3000
(c)
25020015010050
10000
(a)
0.2
–0.2
–0.4
–0.6
–0.8
–1.0
0
0.2
0.1
–0.2
–0.1
–0.3
0
0.4
0.8
0.6
0.4
0.2
–0.2
–0.4
–0.6
–0.8
0
1.0
0.5
–0.5
–1.5
–2.5
–1.0
–2.0
0
1.0
800600400200
1400
(d)
1200800600400200
500
(b)
40302010
1000
Food availability (kg ha
–1
)
Food availability (kg ha
–1
)
Food availability (per capita)
Food availability (voles)
Annual population
growth rate
Annual population
growth rate
Figure 9.2
Increases in annual population
growth rate with the availability of
food, measured as pasture biomass
(kg ha
−1
) in (a) and (c), as vole
abundance in (b) and as availability
of food per capita in (d). (a) Red
kangaroo (Bayliss, 1987). (b) Barn
owl (modified from Taylor, 1994).
(c) Wildebeest (Krebs et al., 1999).
(d) Feral pig (Choquenot, 1998).
Positive growth rates indicate
increasing abundance; negative
growth rates decreasing abundance.
AFTER SIBLY & HONE, 2002
many populations are
very stable...
. . . but stability need not mean
‘nothing changes’
9781405156585_4_009.qxd 11/5/07 14:56 Page 284
9.2.2 Determination and regulation of abundance
Is the move from eight to 12 pairs of swifts over 200 years an indication of con-
sistency or of change? Is the similarity between eight and 12 of most interest – or
the difference between them? Some investigators have emphasized the apparent
constancy of populations; others have emphasized the fluctuations.
Those who have emphasized constancy argue that we need to look for stabil-
izing forces within populations to explain why the populations do not exhibit
unfettered increase or a decline to extinction (generally, density-dependent forces:
for instance, competition between crowded individuals for limited resources).
Those who have emphasized fluctuations often look to external factors, weather
or disturbance, to explain the changes. Can the two sides be brought together to
form a consensus?
Chapter 9 From populations to communities
285
(a) AFTER SYMONIDES, 1979; (b) AFTER SINGLETON ET AL., 2001
Number of individuals per plot
0
1968
800
1000
(a)
600
400
200
19701969 1975197419731972
1971
Abundance index
0
(b)
100
50
20001998
150
200
250
300
199619941992199019881986
1984
Beginning of germination
Maximum germination
End of seedling phase
Flowering
Fruiting
Vegetative growth
Year
Year
Figure 9.3
(a) The population dynamics of
Androsace septentrionalis during an
8-year study. (b) Irregular irruptions
in the abundance of house mice
(Mus domesticus) in an agricultural
habitat in Victoria, Australia, where
the mice, when they irrupt, are
serious pests. The ‘abundance index’
is the number caught per 100 trap-
nights. In the fall of 1984 the index
exceeded 300.
9781405156585_4_009.qxd 11/5/07 14:56 Page 285
To do so, it is important to understand clearly the difference between ques-
tions about the ways in which abundance is determined and questions about the
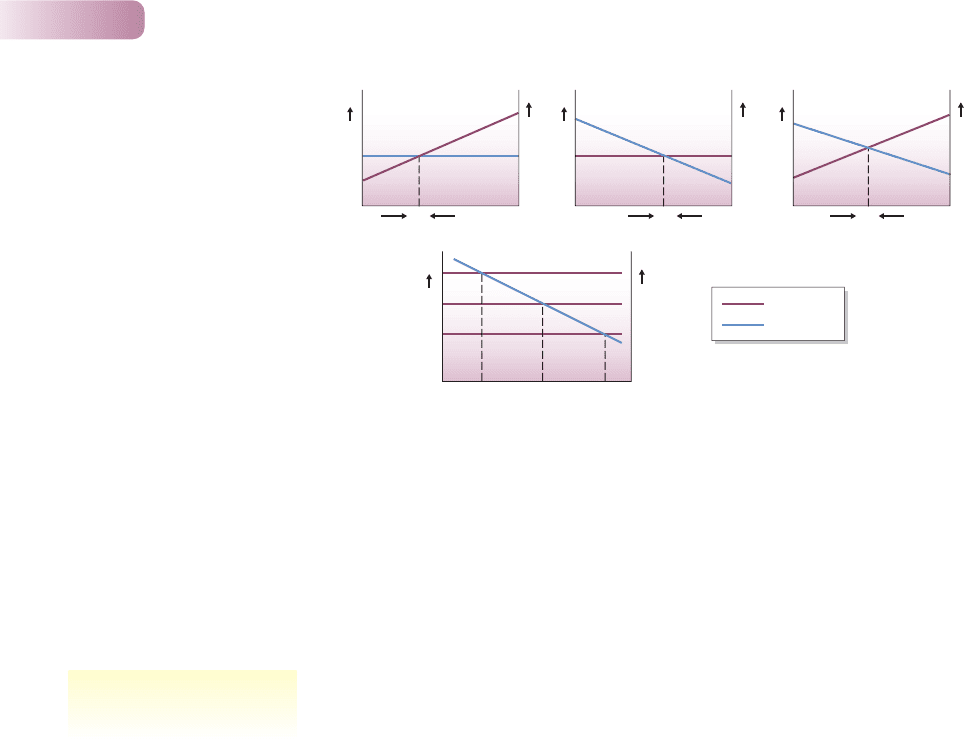
way in which abundance is regulated. Regulation is the tendency of a population
to decrease in size when it is above a particular level, but to increase in size when
below that level. In other words, regulation of a population can, by definition,
occur only as a result of one or more density-dependent processes (see Chapters 3
and 5) that act on rates of birth and/or death and/or movement (Figure 9.4a).
Various potentially density-dependent processes have been discussed in earlier
chapters on competition, predation and parasitism. We must look at regulation,
therefore, to understand how it is that a population tends to remain within
defined upper and lower limits.
On the other hand, the precise abundance of individuals will be determined by
the combined effects of all the factors and all the processes that affect a popula-
tion, whether they are dependent or independent of density (Figure 9.4b). We
must look at the determination of abundance, therefore, to understand how it is
that a particular population exhibits a particular abundance at a particular time,
and not some other abundance.
In the past, certainly, some have believed that density-dependent, biotic
interactions play the main role not only in regulating but also in determining
population size, holding populations in a state of balance in their environments.
Others have felt that most natural populations could be viewed as passing
through a repeated sequence of setbacks and recovery. This view tends to
reject any subdivision of the environment into density-dependent and density
independent ‘factors’, preferring instead to see populations as sitting at the
Part III Individuals, Populations, Communities and Ecosystems
286
(a)
(b)
(i) (ii) (iii)
Population size
b
Population size
Birth rate
Death rate
Death rate
Birth rate
Death rate
Birth rate
Death rate
Birth rate
Birth rate
Death rate
N*
N
1
* N
2
* N
3
*
N* N*
d
1
d
2
d
3
Figure 9.4
(a) Population regulation with: (i) density-independent birth and density-dependent death; (ii) density-
dependent birth and density-independent death; and (iii) density-dependent birth and death. Population
size increases when birth rate exceeds death rate and decreases when death rate exceeds birth rate.
N* is therefore a stable equilibrium population size. The actual value of the equilibrium population size
is seen to depend on both the magnitude of the density-independent rate and the magnitude and slope
of any density-dependent processes. (b) Population regulation with density-dependent birth, b, and
density-independent death, d. Death rates are determined by physical conditions which differ in three
sites (death rates d
1
, d
2
and d
3
). Equilibrium population size varies as a result (N
1
*, N
2
*, N
3
*).
the distinction between
determination and regulation
9781405156585_4_009.qxd 11/5/07 14:56 Page 286

center of an ecological web, where various factors and processes interact in their
effects on the population.
There is really no conflict between the two views. The first is preoccupied with
what regulates population size and the second with what determines popula-
tion size – and both are perfectly valid interests. No population can be absolutely
free of regulation – long-term unrestrained population growth is unknown,
and unrestrained declines to extinction are rare. Furthermore, any suggestion
that density-dependent processes are rare or generally of only minor importance
would be wrong. A very large number of studies have been made of various kinds
of animals, especially of insects. Density dependence has by no means always
been detected but it is commonly seen when studies are continued for many
generations. For instance, density dependence was detected in 80% or more of
studies of insects that lasted for more than 10 years (Hassell et al., 1989; Woiwod
& Hanski, 1992).
On the other hand, for many populations weather is typically the major deter-
minant of abundance and other factors are of relatively minor importance. For
instance, in one famous, classic study of a pest, apple thrips, weather accounted for
78% of the variation in the number of thrips (Davidson & Andrewartha, 1948);
for predicting thrips’ abundance, information on the weather is of paramount
importance. So, what regulates the size of a population need not determine its size
for most of the time. It would be wrong to give regulation or density dependence
some kind of pre-eminence. It may be occurring only infrequently or intermittently,
and it is likely that no natural population is ever truly at equilibrium: even when
regulation is occurring, it may be drawing abundance toward a level that is itself
changing in response to changing levels of resources. Thus, there are a range of
possibilities: some populations in nature are almost always recovering from the
last disaster (Figure 9.5a), others are usually limited by an abundant resource
(Figure 9.5b) or by a scarce resource (Figure 9.5c), and others are usually in
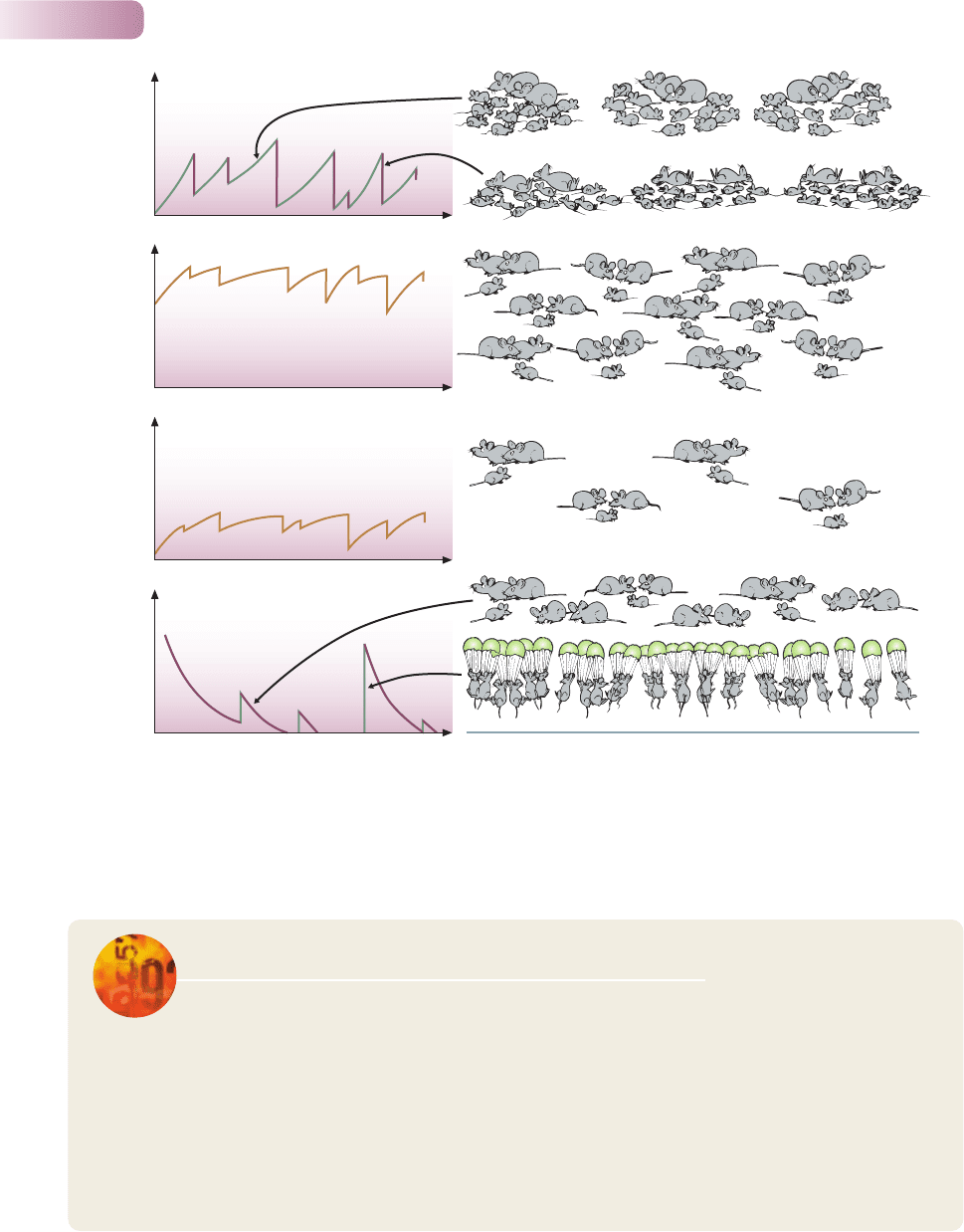
decline after sudden episodes of colonization (Figure 9.5d).
9.2.3 Key factor analysis
We can distinguish clearly between what regulates and what determines the
abundance of a population, and see how regulation and determination relate
to one another, by examining an approach known as key factor analysis. It has
been applied to many insects and some other animals and plants and is based
on calculating what are known as k-values for each phase of the life cycle. In fact,
key factor analysis is poorly named, since it identifies key phases (rather than key
factors) in the life of a study organism (those most important in determining
abundance). Details are described in Box 9.1, but the approach can be under-
stood simply by appreciating that the k-values measure the amount of mortality:
the higher the k-value, the greater the mortality (k stands for ‘killing power’).
For a key factor analysis to be carried out, data are compiled in the form of
a life table (see Chapter 5), such as that done for a Canadian population of the
Colorado potato beetle (Leptinotarsa decemlineata) in Box 9.1. The sampling
program in that case provided estimates of the population at seven stages: eggs,
early larvae, late larvae, pupae, summer adults, hibernating adults and spring
adults. One further category was included, females × 2, to take account of any
unequal sex ratios among the summer adults.
Chapter 9 From populations to communities
287
both are perfectly valid interests
Colorado potato beetles
9781405156585_4_009.qxd 11/5/07 14:56 Page 287
Part III Individuals, Populations, Communities and Ecosystems
288
(a)
(d)
(c)
(b)
Population size
Time
Figure 9.5
Idealized diagrams of population dynamics: (a) dynamics dominated by phases of population growth after disasters; (b) dynamics dominated
by limitations on environmental carrying capacity, where the carrying capacity is high; (c) same as (b) but where the carrying capacity is low;
(d) dynamics within a habitable site dominated by population decay after more or less sudden episodes of colonization or recruitment.
9.1 QUANTITATIVE ASPECTS
9.1 Quantitative aspects
Table 9.1 sets out a typical set of life table data,
collected by Harcourt (1971) for the Colorado potato
beetle, Leptinotarsa decemlineata, in Canada. The
first column lists the various phases of the life cycle.
Spring adults emerge from hibernation around the
middle of June, when potato plants are breaking
through the ground. Within 3 or 4 days egg laying
begins, and it continues for about 1 month. The eggs
Determining k-values for key factor analysis
9781405156585_4_009.qxd 11/5/07 14:56 Page 288