Townsend C.R., Begon M., Harper J.L. Essentials of Ecology
Подождите немного. Документ загружается.


Chapter 8 Evolutionary ecology
259
For example, the 10 species of Pacific salmon,
Oncorhynchus spp., can be effectively distinguished
from one another by RFLP profiling of targeted nuclear
genes (Withler et al., 2004). Some results of applying
such analyses to cases of suspected illegal posses-
sion of salmon are shown in Table 8.1. Case 2, for
instance, involved a disaffected chef reporting a
restaurant owner to the authorities. A fish was iden-
tified as a coho salmon, O. kisutch, which, because it
showed no signs of having been frozen, could not
have come from the previous years’ legal harvest. The
owner was duly fined.
Moreover, analyses based largely on micro-
satellites, with their finer scale of resolution, are able,
even within a species, to tie a sample to a particular
river – if not with certainty then at least with a very
high probability. Some results of these analyses are
shown in Table 8.2. In case 2 here, for instance, illeg-
ally sourced Fraser River sockeye salmon, O. nerka,
were identified in an analysis of 50 cans of salmon
and the defendant, fined $15,000, was found to be
in possession of 100,000 cans with a ‘street value’ of
$300,000–400,000.
What do you think of the level of the fines imposed?
How does the seriousness of crimes like this compare
to those of other crimes: street robbery or the posses-
sion of illegal drugs for personal use? Should those
convicted be punished in proportion to the economic
harm they may be doing to these particular fisheries,
or should their fines be seen as a signal sent out to
all those who ignore the need to restrain activity in
exploited but vulnerable populations and to conserve
them for future generations?
AFTER WITHER ET AL., 2004
CASE (YEAR) TISSUES RESULT LEGAL OUTCOME FINE ($)
1 (1995) Blood/scales/slime from containers Coho Conviction 1500
2 (1998) Muscle Chum Conviction 1800
Chinook
Coho
3 (1998) Muscle Coho Conviction ?
4 (1999) Muscle Atlantic No charges –
Chinook
Coho
5 (2000) Muscle Coho Guilty plea 7500
6 (2000) Muscle Sockeye Conviction 1000
Table 8.1
Species identification of salmonid samples obtained by fisheries officers in Canada because the material was believed to have been
obtained illegally.
AFTER WITHER ET AL., 2004
CASE (YEAR) SPECIES RESULTS OUTCOME FINE ($)
1 (1998) Sockeye 96.5% Fraser; 96.5% IF&T Guilty plea 2,000
2 (1999) Sockeye 100% Fraser; 100% IF&T Conviction 15,000
3 (1999) Chinook 91.4% Fraser No conviction, under appeal
4 (2000) Sockeye 100% Fraser; 100% IF&T Guilty plea 8,000
5 (2001) Sockeye 97.8% Fraser; 97.8% IF&T Guilty plea 3,000
Table 8.2
Stock identification of salmonid samples obtained by fisheries officers in Canada because the material was believed to have been obtained
illegally. IF&T refers to the Interior Fraser and Thompson tributaries.
9781405156585_4_008.qxd 11/5/07 14:54 Page 259

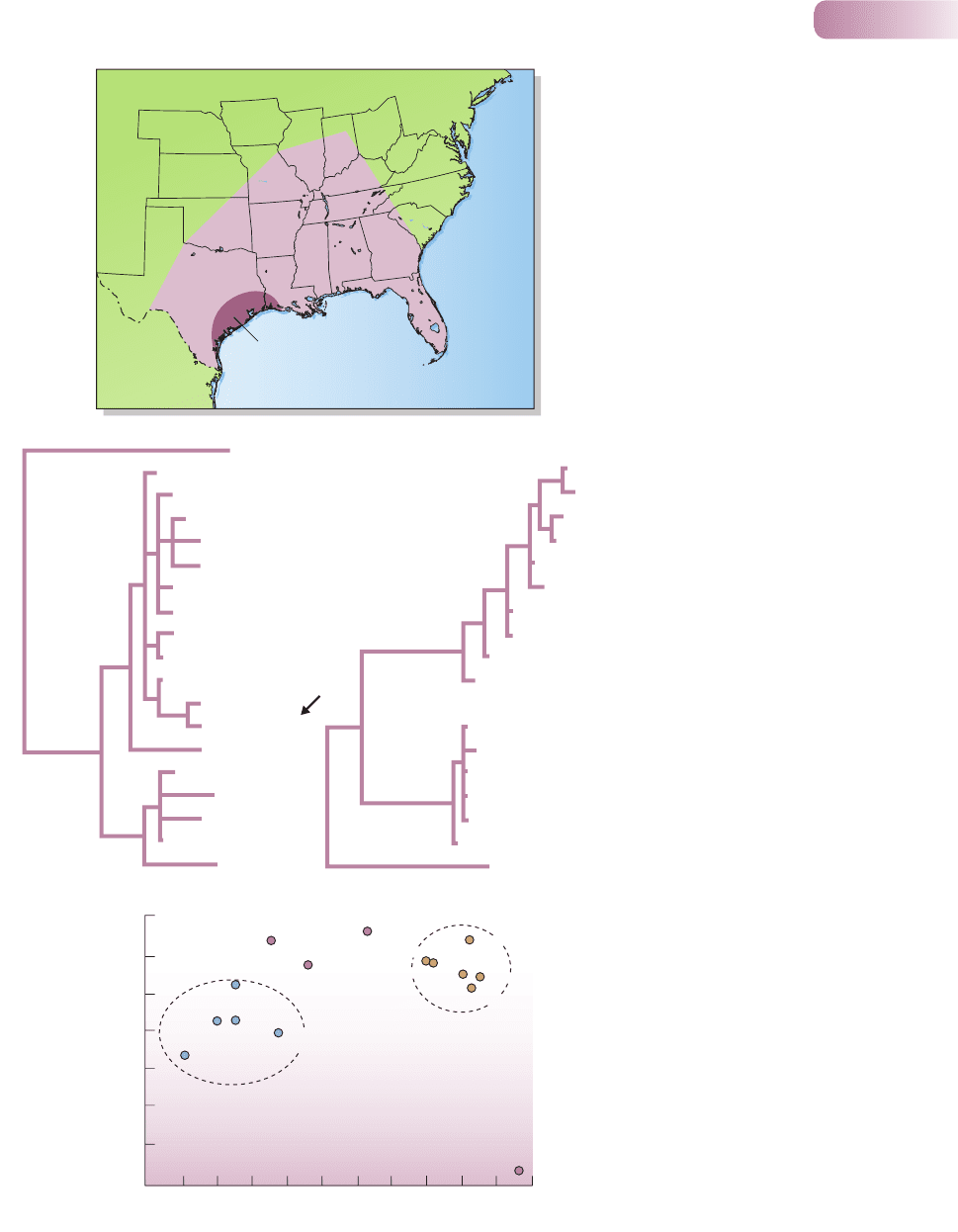
8.2.3 Differentiation between species: the red wolf
Issues in conservation surface again when we shift our focus from differentiation
within to differentiation between species. The red wolf, Canis rufus, once had
a widespread distribution in the southeastern United States (Figure 8.4a), but
when, by the mid-1970s, that distribution had shrunk to a single population in
eastern Texas, the US Fish and Wildlife Service instituted an emergency program
to save it from extinction. Fourteen individuals were rescued from its final refuge
and bred in captivity with a view to subsequent reintroduction in the wild. In the
United States as a whole, the red wolf coexists with two other, closely related
species, the gray wolf, C. lupus, and the coyote, C. latrans. Traditional analyses,
based on morphological features, placed the red wolf as a genuine, separate species,
intermediate in many ways between the gray wolf and the coyote (Nowak, 1979).
However, as we shall see below, molecular markers suggest strongly that the red
wolf is a hybrid arising from interbreeding between gray wolves and coyotes.
A number of questions therefore suggest themselves (Wayne, 1996), including:
‘Should the conservation status of the red wolf, and the amount of money spent
on its conservation, be downgraded if it is acknowledged that it is ‘only’ a hybrid
and not a full species?’ And will attempts to save the red wolf by reintroduction be
doomed, in any case, because of ‘introgression’ – the movement of genes from gray
wolves or coyotes into the red wolf gene pool as a result of interbreeding?
The first molecular markers used to assess the degree of genetic isolation of
red wolves from gray wolves and coyotes, albeit for a relatively small sample,
were from mtDNA – both restriction fragment genotypes (RFLPs – see Box 8.1)
and sequence variation within the cytochrome b gene. From the restriction site
analysis carried out on contemporary captures (Figure 8.4b), it is clear, first, that
the gray wolf and coyote samples were quite separate from one another; but
also that samples from captive red wolves all fitted squarely within the cluster of
coyote genotypes. And when sequence analysis was applied to museum pelts
of red wolves from a variety of locations, and to a number of contemporary gray
wolves and coyotes (Figure 8.4c), these too showed separate clusters for gray
wolves and coyotes, and this time that red wolves had either gray wolf or coyote
genotypes. Thus, the status of the red wolf as a separate species was called
seriously into question, and its origin as a gray wolf–coyote hybrid was further
supported by the observation of common, contemporary introgression of coyote
genes into gray wolf populations throughout a region on the USA–Canadian
border, where recent contact (the last 100 years) has been made as coyotes have
moved north (Lehmann et al., 1991).
Investigations of microsatellites in the nuclear DNA have further clarified
the red wolf story (Roy et al., 1994). First, studies on the USA–Canadian border
confirmed the high frequency of contemporary coyote introgression into gray
wolf gene pools (Figure 8.4d). Second, an analysis of 40 captive red wolves
revealed that every one of the 53 microsatellite alleles they carried was also found
in coyotes. Museum specimens of red wolves, too, failed to turn up unique red
wolf alleles, and indeed, the historical and contemporary red wolf samples were
themselves very similar. Finally, overall, red wolf samples, like contemporary
gray wolf samples in the zone of hybridization, appear intermediate between
coyotes and non-hybridizing gray wolves (Figure 8.4d). All of this argues in
favor of the red wolf having its origins in gray wolf–coyote hybridization, with
Part III Individuals, Populations, Communities and Ecosystems
260
species or hybrid?
mtDNA
nuclear microsatellites
9781405156585_4_008.qxd 11/5/07 14:54 Page 260
Chapter 8 Evolutionary ecology
261
(a)
(b) (c)
NE
KS
IA
TX
AR
MO
KY
TN
AL
GA
FL
OH
WV
VA
NC
SC
MS
IL
IN
OK
LA
1970
Gray-1
Gray-1
Gray-4
Gray-MEX
Coyote-1
Coyote-2
Coyote-5
Coyote-11
Coyote-3
Coyote-7
Coyote-22
Coyote-14
Coyote-24
Coyote-1
Coyote-21
Coyote-24
Coyote-30
Coyote-13
Coyote-20
Coyote-14
Coyote-25
Coyote-32/Red
Coyote-8
Coyote-3
Coyote-7
Coyote-20
Coyote-22
Coyote-26
Red-ARK2
Red-MO
Red-CAP
Red-ARK1
Red-LA
Red-OK
Red-TX
Golden Jackal
–2.0
–2.5
1.0
0.5
1.0–0.6–0.8–1.0–1.2 –0.4 –0.2 0 0.2 0.4 0.6 0.8
0
–0.5
–1.0
–1.5
(d)
Dimension 1
Dimension 2
Gray wolves
non-hybridizing
Coyotes
Vancouver
North West
Territories
N. Quebec
Alberta Kenai
Kenai
Alberta
Maine
Minnesota
Washington
California
Red wolf
Minnesota
wolves
S. Quebec
wolves
Golden
jackal
Figure 8.4
(a) The geographic range (light
maroon) of the red wolf, Canis rufus,
in the United States around 1700,
and within that the smaller bounded
area showing its range in
southeastern Texas around 1970.
(b) A ‘phylogenetic tree’ of coyote
and red wolf mtDNA restriction-site
genotypes (RFLPs). In a phylogenetic
tree, the most similar (closely
related) types are placed closest
together, then linked to the type that
is most similar to them, and so on,
the lengths of the horizontal lines
representing the degree of difference.
The tree is ‘rooted’ (to give it context)
by inclusion of a gray wolf (Gray-1).
The numbers refer to different
individuals. The arrow points to
the single genotype shared by the
eight captive red wolves that were
sampled, which is clearly simply
part of the coyote ‘cluster’. (c) A
phylogenetic tree constructed on
similar principles but based on
sequences of the cytochrome b gene
in the mtDNA. Museum red wolf
samples are from Arkansas (ARK),
Missouri (MO), Louisiana (LA),
Oklahoma (OK) and Texas (TX); CAP
refers to a captive red wolf; and MEX
refers to a gray wolf from Mexico.
The tree is rooted by the inclusion
of sequence data from the golden
jackal, C. aureus. The red wolf
genotypes are clearly parts of either
the coyote or the gray wolf clusters.
(d) The relationships between
various coyote, gray wolf and red
wolf populations at 10 nuclear DNA
microsatellite loci, as demonstrated
by an analysis that condenses the
data from these 10 loci into two
dimensions. The details of this
analysis are unimportant here, as
long as it is appreciated that the
most similar populations are closest
together in the figure. There are two
clusters: coyotes and gray wolves
from populations in which there is
no hybridization with coyotes. Red
wolves, and the populations of gray
wolves from Minnesota and south
Quebec where there is hybridization
with coyotes, are located between
these two clusters. Context, again,
is provided by the location of the
golden jackal.
(a–c) AFTER WAYNE & JENKS, 1991; (d) AFTER ROY ET AL., 1994
9781405156585_4_008.qxd 11/5/07 14:54 Page 261

subsequent further hybridization with coyotes, as gray wolves became rare in the
southeastern USA.
In answer to our original questions, then, (i) the red wolf seems, ultimately,
to be a hybrid rather than a separate species with a more ancient origin, and (ii)
any program of reintroduction clearly is in danger of failing as a result of intro-
gression from coyotes, requiring sufficient densities of red wolves to minimize
this possibility, and perhaps even barriers to the ‘species’ meeting (Fredrickson
& Hedrick, 2006). However, whether biological status and practical difficulties
combine to undermine even the desirability of reintroducing red wolves is not
simply a scientific question. Public perception and opinion (in this case regarding
the conservation importance of the red wolf) must also be taken into account.
Similar remarks apply to most conservation issues, especially when public funds
are involved. A molecular ecology perspective has been immensely informative –
but information may sometimes muddy rather than clarify the waters.
8.3 Coevolutionary arms races
We turn now from evolution at the molecular level to evolution at the level of
species interactions, starting with those in which species are ‘in opposition’ to
one another. Following some general background, we turn first to interactions
between insects and the plants they eat (Section 8.3.2) and then to those between
parasites and their hosts (Section 8.3.3).
8.3.1 Coevolution
The dynamics of consumer resource pairs (see Chapter 7) are linked to the
dynamics of whole webs of interacting species (see Chapter 9) by how specialized
or generalized particular consumers are. Generalists draw the species of a com-
munity together into large interactive networks. Specialists divide communities
into detached or semidetached compartments. Coevolution plays a vital part in
determining how specialized or generalized particular consumers are.
It is not surprising, as we saw in Chapter 3, that many organisms have evolved
defenses that reduce the chance of an encounter with a consumer and/or increase
the chance of surviving such an encounter. But the interaction does not necessarily
stop there. A better defended food resource (the ‘prey’) itself exerts a selection
pressure on consumers to overcome that defense. A consumer that does so is
likely to have invested in counteracting that defense as opposed to others, and will
steal a march on its competitors, and so is likely to become relatively specialized
on that prey type – which is then under particular pressure to defend itself against
that particular consumer, and so on. A continuing interaction can therefore be
envisaged in which the evolution of both the consumer and the prey depend
crucially on the evolution of the other: what Ehrlich and Raven (1964) called a
coevolutionary ‘arms race’, which, in its most extreme form, has a coadapted pair
of species locked together in perpetual struggle.
Indeed, what is unacceptable to most animals may be the chosen, even unique,
diet of others. It is, after all, an inevitable consequence of having evolved resistance
to a prey’s defenses that a consumer will have gained access to a resource unavailable
to most (or all) other species. For example, the tropical legume Dioclea metacarpa
Part III Individuals, Populations, Communities and Ecosystems
262
one man’s poison is another
man’s meat
9781405156585_4_008.qxd 11/5/07 14:54 Page 262
is toxic to almost all insect species because it contains a non-protein amino acid,
L-canavanine, which those insects incorporate (lethally) into their proteins in place
of arginine. But a species of bruchid beetle, Caryedes brasiliensis, has evolved a
modified enzyme that distinguishes between
L-canavanine and arginine, and the
larvae of these beetles feed solely on D. metacarpa (Rosenthal et al., 1976).
8.3.2 Insect–plant arms races
We discussed in Section 3.4.2 how attacks by herbivores select for plant-defensive
chemicals. We also saw that these can be divided into ‘qualitative’ chemicals that
are poisonous, can kill in small doses and tend to be induced by herbivore attacks,
and ‘quantitative’ chemicals that are digestion-reducing, rely on an accumula-
tion of ill effects and tend to be produced constitutively (i.e. all the time). These
chemicals will select for adaptations in herbivores that can overcome them. It
seems probable, however, that toxic chemicals, by virtue of their specificity, are
likely to be the foundation of an arms race, requiring an equally specific response
from a herbivore; whereas chemicals that make plants generally indigestible are
much more difficult to overcome through any ‘targeted’ adaptation (Cornell &
Hawkins, 2003). Put simply: plants relying on toxins are more prone to becoming
involved in arms races with their herbivores (like the beetle and legume described
above) than those relying on more ‘quantitative’ chemicals.
We can seek evidence for the toxin arms race hypothesis by asking whether
specialist herbivores generally, locked in their coevolutionary arms races, per-
form better when faced with their plants’ toxic chemicals than generalists; whereas
generalists, having invested in overcoming a wide range of chemicals, perform
better than specialists when faced with chemicals that have not provoked
coevolutionary responses. Such evidence is provided by an analysis of a wide
range of data sets for insect herbivores fed on artificial diets with added chemicals
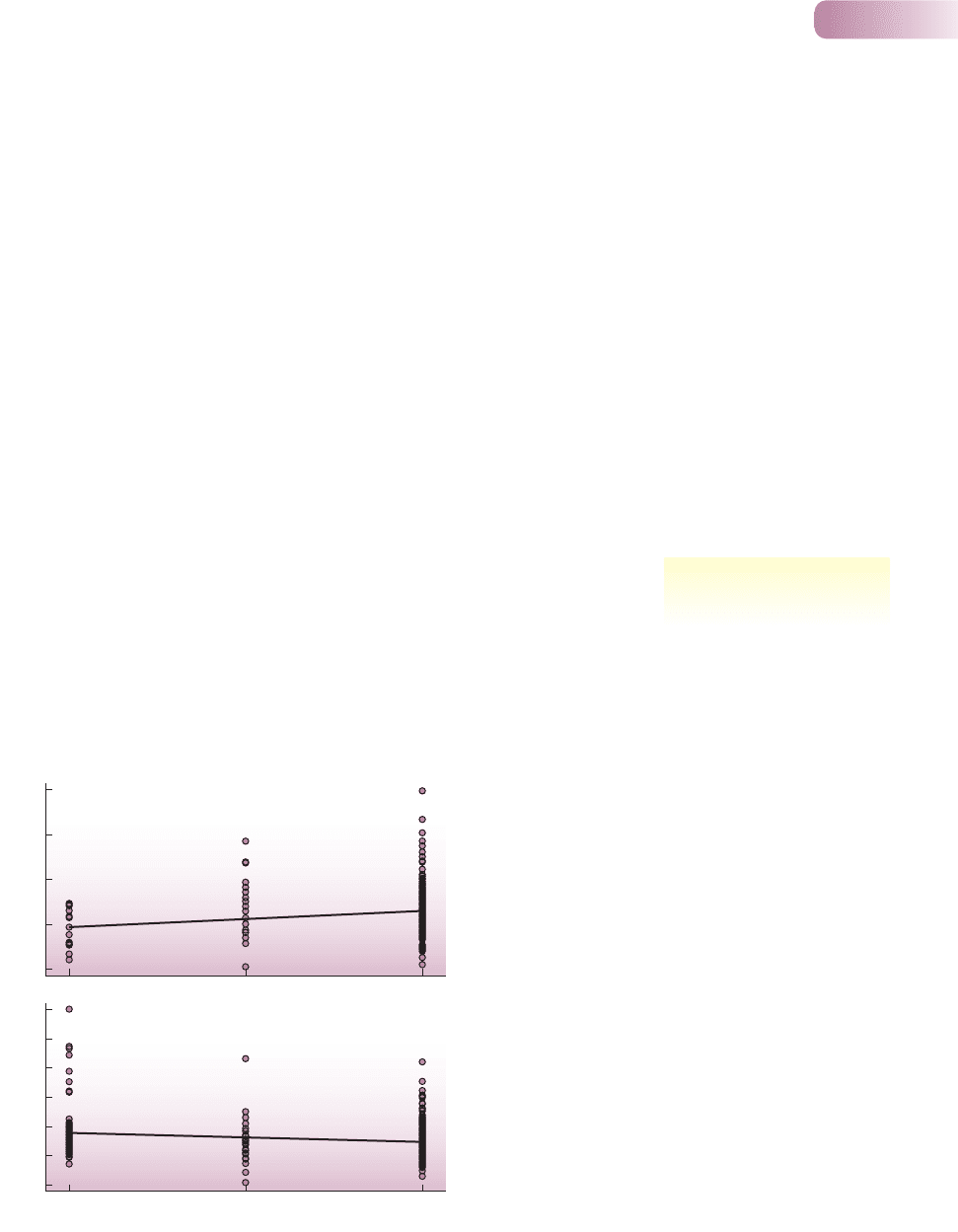
(892 insect–chemical combinations; Figure 8.5).
Chapter 8 Evolutionary ecology
263
specialists are more prone to
arms races
31
1.3
5.3
2
312
(a)
–3
5
7
9
Specialism group
(b)
3
1
–1
ToxicityToxicity
–2.7
3.3
–0.7
Figure 8.5
Combining data from a wide range of published studies, insect
herbivores were split into three groups: 1, specialists (feeding from
one or two plant families); 2, ‘oligophages’ (3–9 families); and 3,
generalists (more than nine families). Chemicals were split into
two groups: (a) those that are found in the normal hosts of the
specialists and oligophages, and (b) those that are not. ‘Toxicity’ is
measured from the mortality rates of insects on a standardized
scale, since many studies have been combined. (a) It is apparent
that more specialized insects suffered lower mortality on chemicals
that have provoked a coevolutionary response from specialist
herbivores. (b) It is apparent that more generalist insects
suffered lower mortality on chemicals that have not provoked a
coevolutionary response from specialist herbivores. P < 0.005 in
both cases.
AFTER CORNELL & HAWKINS, 2003
9781405156585_4_008.qxd 11/5/07 14:54 Page 263
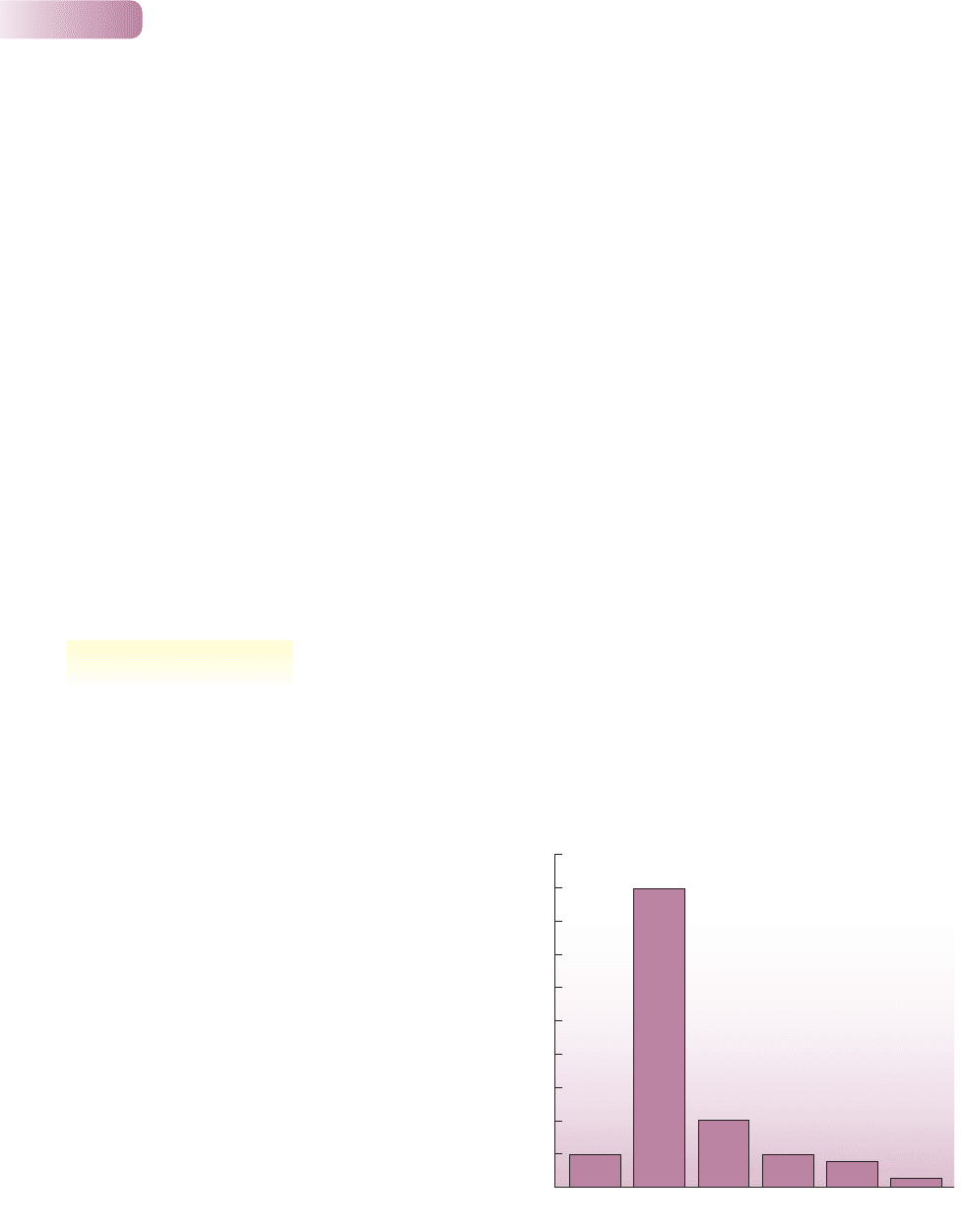
8.3.3 Coevolution of parasites and their hosts
The intimate association between parasites and their hosts makes them especially
prone to coevolutionary arms races. Indeed, the specialization may go further
than that between species. Within species, it is common to find a high degree of
genetic variation in the virulence of parasites and/or in the resistance or immunity
of hosts. Every few years, for example, as we are perhaps more aware than ever,
a new strain of the influenza virus evolves of sufficient virulence and novelty
to generate a widespread epidemic and mortality in human populations that had
been relatively resistant to previously circulating strains. No strain has been more
devastating – at the time of writing – than the worldwide epidemic (pandemic) of
Spanish flu that followed World War I in 1918/19 and killed 20 million people –
many more than died in the war itself. Human diseases can also provide examples
of variation in host resistance. When the Native Americans of the Canadian
Plains were forcibly settled onto reservations in the 1880s, their death rate due
to tuberculosis (TB) initially exploded but then gradually declined (Figure 8.6).
Environmental factors (inadequate diet, overcrowding, spiritual demoralization)
undoubtedly played some part in this, but variation in resistance is also likely
to have been significant. The mortality rate among the Native Americans was
often 20 times that of the surrounding European colonist population, living
in similar conditions but having been exposed previously to TB. Some native
families had a particularly low mortality rate in the 1880s epidemic, and many of
the survivors in the 1930s were descendants of those families (Ferguson, 1933;
Dobson & Carper, 1996).
It may seem straightforward that parasites in a population select for the
evolution of more resistant hosts, which in turn select for more infective parasites:
a classic arms race. In fact, the process is not necessarily so straightforward,
though there are certainly examples where host and parasite drive one another’s
evolution. A most dramatic example involves the rabbit and the myxoma virus,
which causes myxomatosis. The virus originated in the South American jungle
rabbit Sylvilagus brasiliensis, where it causes a mild disease that only rarely kills
Part III Individuals, Populations, Communities and Ecosystems
264
100
90
80
70
60
50
40
30
20
10
0
Deaths per 1000 individuals
1881 1886 1901 1907 1926 1930
Year
Figure 8.6
The mortality rate due to tuberculosis in three generations of
Canadian Plains Native Americans after their forced settlement
onto reservations.
AFTER FERGUSON, 1933; DOBSON & CARPER, 1996
myxomatosis
9781405156585_4_008.qxd 11/5/07 14:54 Page 264
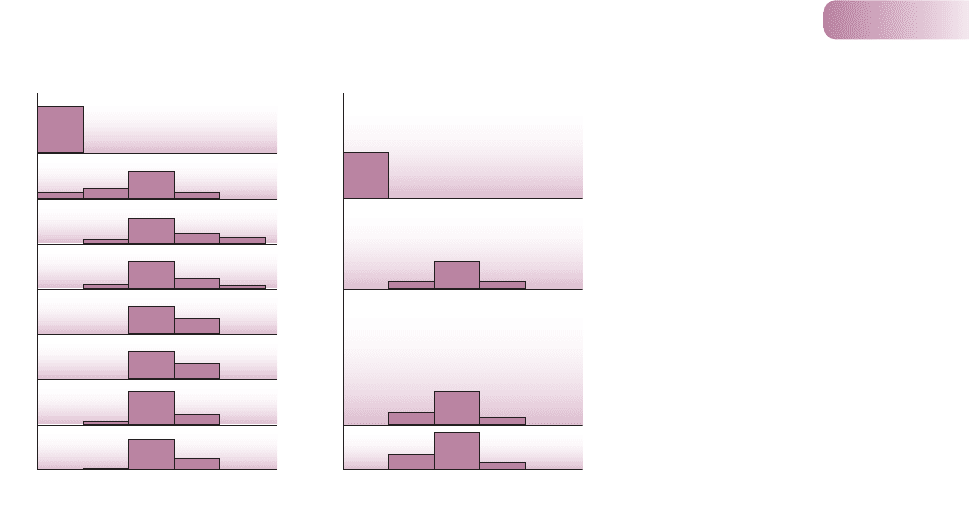
the host. The South American virus, however, is usually fatal when it infects the
European rabbit Oryctolagus cuniculus. In one of the greatest examples of bio-
logical pest control, the myxoma virus was introduced into Australia in the 1950s
to control the European rabbit, which had become a pest of grazing lands. The
disease spread rapidly in 1950/51, and rabbit populations were greatly reduced
– by more than 90% in some places. At the same time, the virus was introduced
to England and France, and there too it resulted in huge reductions in the rabbit
populations. The evolutionary changes that then occurred in Australia were
followed in detail by Fenner and his associates, who had the brilliant foresight
to establish baseline genetic strains of both rabbits and virus (Fenner, 1983). They
used these to measure subsequent changes in the virulence of the virus and the
resistance of the host as they evolved in the field.
When the disease was first introduced to Australia it killed more than 99% of
infected rabbits. This ‘case mortality’ fell to 90% within 1 year and then declined
further. The virulence of virus isolates was graded according to host survival time
and the case mortality of control rabbits. The original, highly virulent virus was
grade I, which killed > 99% of infected laboratory rabbits. Already by 1952, most
of the virus isolates from the field were the less virulent grades III and IV. At the
same time, the rabbit population in the field was increasing in resistance. When
injected with a standard grade III strain, field samples of rabbits in 1950/51 had
a case mortality of nearly 90%, which had declined to less than 30% only 8 years
later (Figure 8.7).
This evolution of resistance is easy to understand: resistant rabbits are obviously
favored by natural selection in the presence of the myxoma virus. The case of the
virus, however, is subtler. The contrast between the virulence of the virus in the
European rabbit and its lack of virulence in the American host with which it had
coevolved, combined with the attenuation of its virulence in Australia and Europe
after its introduction, fit a commonly held view that parasites evolve toward
Chapter 8 Evolutionary ecology
265
1950–51
VIVII
0
I
100
III VIVIII III
0
100
0
100
0
100
1975–81
1952–55
1955–58
1959–63
1964–66
1967–69
1970–74
1976–80
1953
1962
1975
Proportions (%)
(a) Australia (b) Britain
Virulence grade
Figure 8.7
(a) The percentages in which various grades
of myxoma virus have been found in wild
populations of rabbits in Australia at different
times from 1951 to 1981. Grade I is the most
virulent. (After Fenner, 1983.) (b) Similar data
for wild populations of rabbits in Great Britain
from 1953 to 1980.
FROM FENNER, 1983; AFTER MAY & ANDERSON, 1983
9781405156585_4_008.qxd 11/5/07 14:54 Page 265

becoming benign to their hosts in order to prevent the parasite eliminating its
host and thus eliminating its habitat. This view, however, is quite wrong. The
parasites favored by natural selection are those with the greatest fitness (broadly,
the greatest reproductive rate). Sometimes this is achieved through a decline
in virulence, but sometimes it is not. In the myxoma virus, an initial decline in
virulence was indeed favored – but further declines were not.
The myxoma virus is blood-borne and is transmitted from host to host by
blood-feeding insect vectors. In the first 20 years after its introduction to Australia,
the main vectors were mosquitoes, which feed only on live hosts. The problem
for grade I and II viruses is that they kill the host so quickly that there is only a
very short time in which the mosquito can transmit them. Effective transmission
may be possible at very high host densities, but as soon as densities decline, it is
not. Hence, there was selection against grades I and II and in favor of less virulent
grades, giving rise to longer periods of host infectiousness. At the other end of
the virulence scale, however, the mosquitoes are unlikely to transmit grade V
of the virus because it produces very few infective particles. The situation was
complicated in the late 1960s when an alternative vector of the disease, the
rabbit flea Spilopsyllus cuniculi (the main vector in England), was introduced
to Australia, apparently favoring more virulent strains than the mosquitoes had
done. Overall, however, there has been selection in the rabbit–myxomatosis
system not for decreased virulence as such, but for increased transmissibility
(and hence increased fitness) – which happens in this system to be maximized at
intermediate grades of virulence.
In other cases, host–parasite coevolution is more definitely antagonistic:
increased resistance in the host and increased infectivity in the parasite. A
classic example is the interaction between agricultural plants and their pathogens
(Burdon, 1987), though in this case the resistant hosts are often introduced by
human intervention. There may even be gene-for-gene matching, with a particu-
lar virulence allele in the pathogen selecting for a resistant allele in the host,
which in turn selects for alleles other than the original allele in the pathogen,
and so on. In fact, these detailed processes have proved difficult to observe,
but this has been done with a system comprising the bacterium Pseudomonas
fluorescens and its viral parasite, the bacteriophage (or phage) SBW25φ2, where
such evolution is relatively easy to observe because generation times are so short.
Changes in both host and parasite were monitored as 12 replicate coexisting
populations of bacterium and phage were transferred from culture bottle to cul-
ture bottle. It is apparent that the bacteria became generally more resistant to
the phage at the same time as the phage became generally more infective to the
bacteria: each was being driven by the directional selection of an arms race
(Figure 8.8).
This was only apparent, however, because each bacterial strain (from one
of the 12 replicate pairs) was tested against all 12 phage strains, and each phage
strain tested against all bacterial strains, and mean resistances and infectivities
calculated. When, at the end of the experiment (Table 8.3), the resistance of
each bacterial strain was tested against each phage strain in turn, it was clear
that bacteria were almost always most resistant (and often wholly resistant) to
the phage strain with which they coevolved. Clearly, the specific problems posed
by particular phage strains had provoked equally specific evolutionary responses
on the part of the bacterial strains.
Part III Individuals, Populations, Communities and Ecosystems
266
bacteria and bacteriophage
9781405156585_4_008.qxd 11/5/07 14:54 Page 266

8.4 Mutualistic interactions
No species lives in isolation, but often the association with other species is
especially close: for many organisms, the habitat they occupy is an individual of
another species. Parasites live within the body cavities or even the cells of their
hosts, nitrogen-fixing bacteria live in nodules on the roots of leguminous plants,
Chapter 8 Evolutionary ecology
267
Mean resistance
1.0
0.8
0.6
0.4
0.2
0
50403020100
(a)
Mean infectivity
0.8
0.6
0.4
0.2
0
50403020100
(b)
Transfer number
Figure 8.8
(a) Over evolutionary time (1 ‘transfer’ ≈ 8 bacterial generations)
bacterial resistance to phage increased in each of 12 bacterial
replicates (designated by different symbols). ‘Mean’ resistance was
the mean calculated over the 12 phage isolates from the respective
time points. (b) Similarly, phage infectivity increased, where ‘mean’
infectivity was calculated over the twelve bacterial replicates.
AFTER BUCKLING & RAINEY, 2002
Table 8.3
For each of 12 bacterial replicates (B1–B12) and their 12 respective phage replicates (φ1–φ12), entries in the table are the proportion of bacteria
resistant to the phage at the end of a period of coevolution (50 transfers ≈ 400 bacterial generations). Coevolving pairs are shown along the
diagonal in bold. Note that bacterial strains are usually most resistant to the phage strain with which they coevolved.
AFTER BUCKLING & RAINEY, 2002
BACTERIAL REPLICATES
PHAGE REPLICATES B1 B2 B3 B4 B5 B6 B7 B8 B9 B10 B11 B12
φ1 0.8 0.9 1 1 1 1 1 1 0.85 0.85 0.75 0.65
φ2 0.1 1 0.3 1 0.85 0.25 1 1 0.85 0.9 0.8 0.65
φ3 0.75 0.75 1 1 1 0.9 1 1 0.85 0.9 0.9 0.65
φ4 0.15 0.9 0.8 1 0.85 0.6 0.6 1 0.85 1 0.85 0.35
φ5 0.25 0.9 1 1 1 0.9 1 0.8 0.85 1 0.8 0.65
φ6 0.2 1 0.85 0.8 0.75 0.8 0.85 0.9 0.85 0.75 0.45 0.25
φ7 0.2 0.75 0.6 1 0.4 0.45 1 0.9 0.85 1 0.75 0.35
φ8 0 0.95 0.55 0.95 0.35 0.25 0.8 1 0.85 1 0.7 0.25
φ9 0 0.7 0.55 0.45 0.7 0.35 1 1 0.85 1 0.5 0.1
φ10 0 0.7 0.9 0.7 0.55 0.9 1 1 0.7 1 0.5 0.4
φ11 0 0.5 0.9 0.75 0.7 1 1 0.95 0.75 1 1 0.35
φ12 0 0.15 0 0.1 0.65 0.35 1 1 0.7 0.8 0.85 0.4
symbiosis and mutualism
9781405156585_4_008.qxd 11/5/07 14:54 Page 267

and so on. Symbiosis (‘living together’) is the term that has been coined for such
close physical associations between species, in which a ‘symbiont’ occupies a
habitat provided by a ‘host’. In fact, though, parasites are usually excluded from
the category of symbionts, which is reserved instead for interactions where
there is at least the suggestion of mutualism. A mutualistic relationship is simply
one in which organisms of different species interact to their mutual benefit.
Mutualism, therefore, need not involve close physical association: mutualists
need not be symbionts. For example, many plants gain dispersal of their seeds
by offering a reward to birds or mammals in the form of edible fleshy fruits,
and many plants assure effective pollination by offering a resource of nectar in
their flowers to visiting insects. These are mutualistic interactions but they are
not symbioses.
It would be wrong, however, to see mutualistic interactions simply as conflict-
free relationships from which nothing but good things flow for both partners.
Rather, current evolutionary thinking views mutualisms as cases of reciprocal
exploitation where nonetheless each partner is a net beneficiary (Herre & West,
1997).
Mutualisms themselves have often been neglected in the past compared to
other types of interaction, yet mutualists compose most of the world’s biomass.
Almost all the plants that dominate grasslands, heaths and forests have roots
that have an intimate mutualistic association with fungi. Most corals depend on
the unicellular algae within their cells, many flowering plants need their insect
pollinators, and many animals carry communities of microorganisms within their
guts that they require for effective digestion.
The rest of this section is organized as a progression. We start with mutualisms
in which no intimate symbiosis is involved; rather, the association is largely
behavioral: that is, each partner behaves in a manner that confers a net benefit
on the other. By Section 8.4.4, when we discuss mutualisms between animals and
the microbiota living in their guts, we will have moved on to closer associations
(one partner living within the other), and in Sections 8.4.5 and 8.4.6 we examine
still more intimate symbioses in which one partner enters between or within
another’s cells.
8.4.1 Mutualistic protectors
‘Cleaner’ fish, of which at least 45 species have been recognized, feed on ecto-
parasites, bacteria and necrotic tissue from the body surface of ‘client’ fish.
Indeed, the cleaners often hold territories with ‘cleaning stations’ that their clients
visit – and visit more often when they carry many parasites. The cleaners gain a
food source and the clients are protected from infection. In fact, it has not always
proved easy to establish that clients benefit, but experiments off Lizard Island on
Australia’s Great Barrier Reef were able to do this for the cleaner fish, Labroides
dimidiatus, which eats parasitic gnathiid isopods from its client fish, Hemigymnus
melapterus. Clients had significantly (3.8 times) more parasites 12 days after
cleaners were excluded from caged enclosures (Figure 8.9a); but even in the short
term (up to 1 day), although removing cleaners, which only feed during daylight,
had no effect when a check was made at dawn (Figure 8.9b), this led to there
being significantly (4.5 times) more parasites following a further day’s feeding
(Figure 8.9c).
Part III Individuals, Populations, Communities and Ecosystems
268
mutualism: reciprocal exploitation
not a cosy partnership
cleaner and client fish
9781405156585_4_008.qxd 11/5/07 14:54 Page 268