Townsend C.R., Begon M., Harper J.L. Essentials of Ecology
Подождите немного. Документ загружается.

and so forth) in all communities, since this will depend on the inherent stability of
the community combined with the variability of the environment. One study tend-
ing to support this investigated 10 small streams in New Zealand that differ in the
intensity and frequency of flow-related disturbances to their beds (Figure 9.26).
Food webs in the more disturbed, ‘unstable’ streams were characterized by less
complex communities: fewer species and fewer links between species.
Chapter 9 From populations to communities
319
0
0.4
0.8
0.6
0.2
0
0.4
0.8
0.6
0.2
0
0.4
0.8
0.6
0.2
0
0.4
0.8
0.6
0.2
0.80.40 0.60.2 0.80.40 0.60.2 0.80.40 0.60.2 0.80.40 0.60.2
Least connected species removed firstRandom species removalMost connected species removed first
Cumulative secondary extinctions / S
Species removed / S
Grassland
(S = 61)
Scotch Broom
(S = 85)
Ythan 1
(S = 124)
Ythan 2
(S = 83)
El Verde
(S = 155)
Canton
(S = 102)
Stony
(S = 109)
Chesapeake
(S = 31)
St Marks
(S = 48)
St Martin
(S = 42)
Little Rock
(S = 92)
Lake Tahoe
(S = 172)
Mirror
(S = 172)
Bridge Brook
(S = 25)
Chachella
(S = 29)
Skipwith
(S = 25)
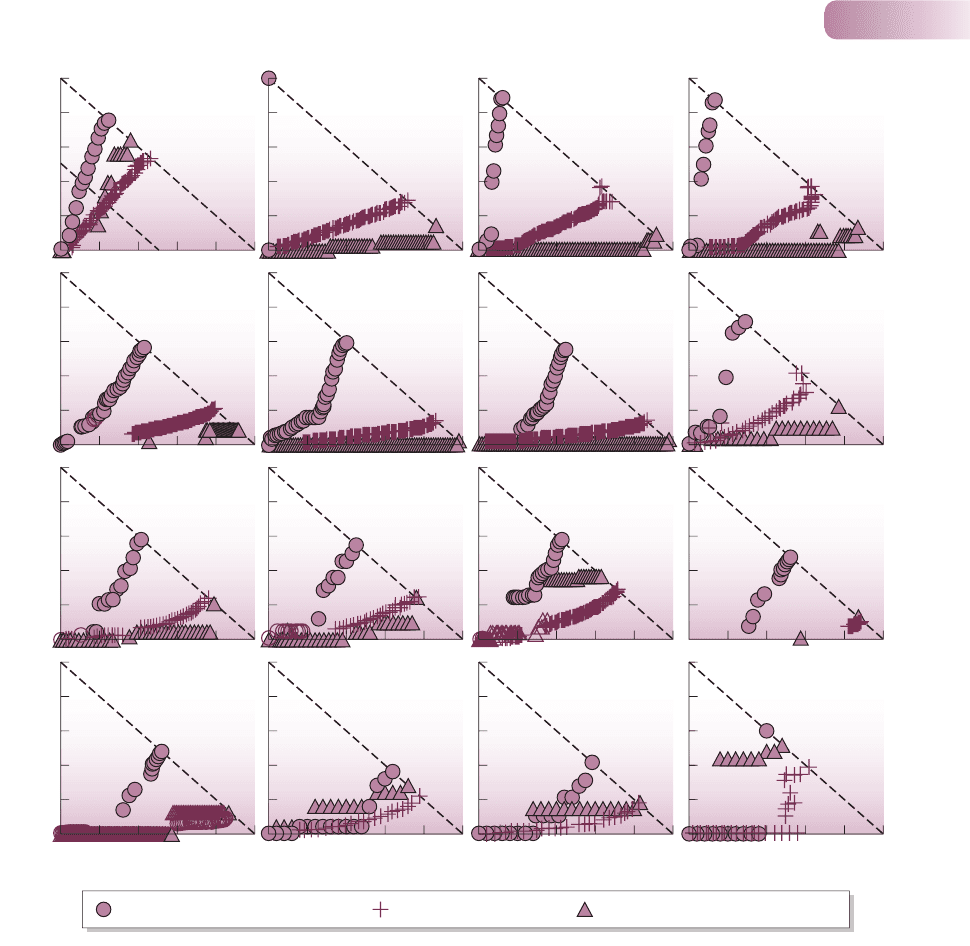
Figure 9.25
The results of a simulation study. The effect of sequential species removal on the number of consequential (secondary) species extinctions, as a
proportion of the total number of species originally in the web, S, for each of 16 previously described food webs. The three different rules for
species removal are described in the lower panel. Robustness of the webs (the tendency not to suffer secondary extinctions) was usually lowest
when the most connected species were removed first and highest when the least connected were removed first.
AFTER DUNNE ET AL., 2002
9781405156585_4_009.qxd 11/5/07 14:56 Page 319

Part III Individuals, Populations, Communities and Ecosystems
320
120
100
80
60
40
20
0
020406080100
020406080100
Intensity of disturbance
18
16
14
12
10
8
6
4
2
0
(a)
(b)
Web size (no. of species)Mean number of feeding links
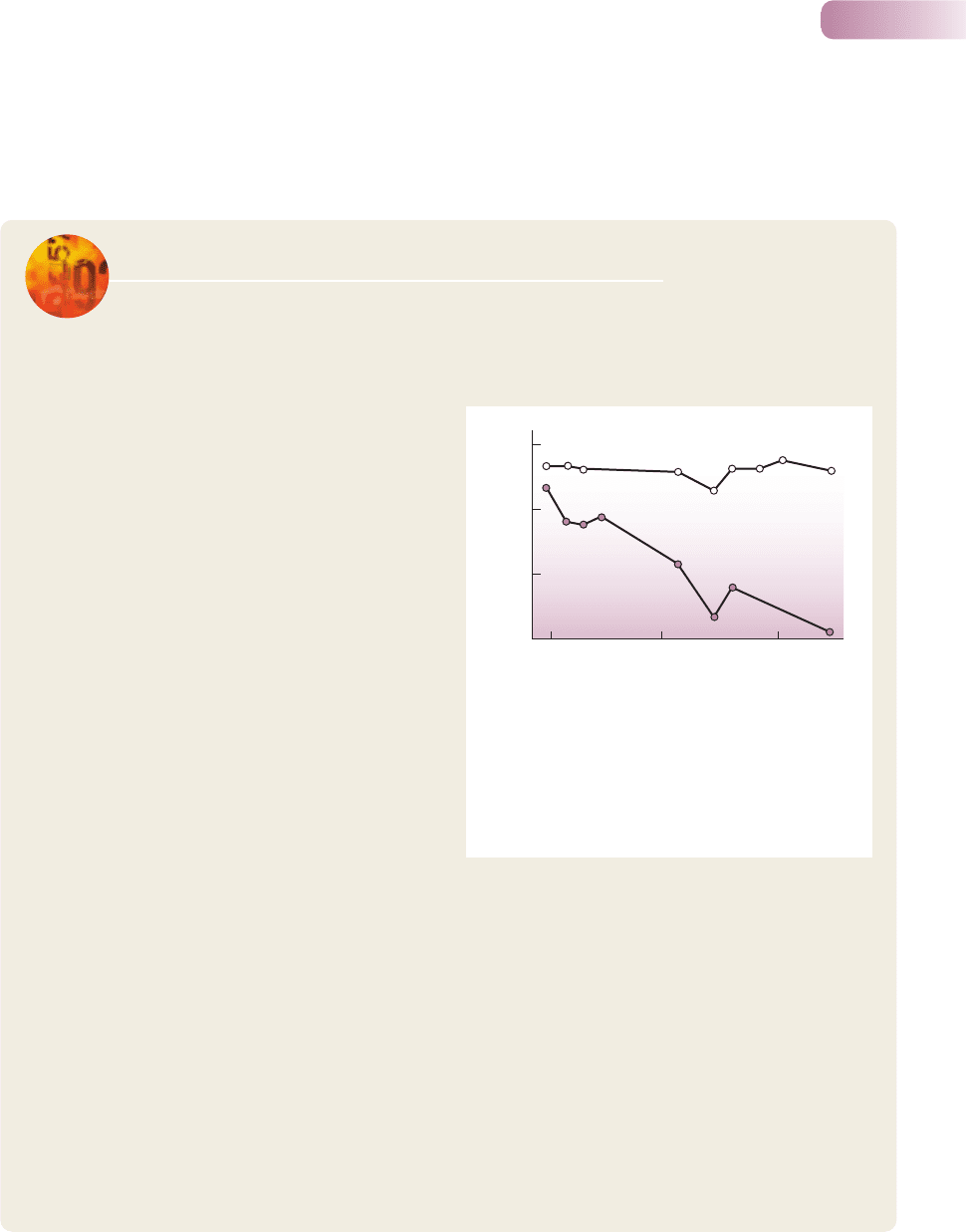
Figure 9.26
In New Zealand streams, less disturbed sites support more
‘complex’ communities, with (a) more species (greater web size)
and (b) greater connectance between species. The average number
of feeding links per animal species (number of prey species in the
diet) increases with the intensity of flow-related disturbances to
the streambed.
AFTER TOWNSEND ET AL., 1998
Multiple determinants of the dynamics of
populations
To understand the factors responsible for the popula-
tion dynamics of even a single species in a single
location, it is necessary to have a knowledge of
physicochemical conditions, available resources, the
organism’s life cycle, and the influence of competi-
tors, predators and parasites on rates of birth, death,
immigration and emigration.
There are contrasting theories to explain the abund-
ance of populations. At one extreme, researchers
emphasize the apparent stability of populations and
SUMMARY
Summary
This line of argument, moreover, carries a further, very important implication
for the likely effects of unnatural perturbations caused by humans on communities.
We might expect these to have their most profound effects on the dynamically
fragile, complex communities of stable environments, which are relatively
unaccustomed to perturbations, and least effect on the simple, robust communities
of variable environments, which have previously been subjected to repeated
(albeit natural) perturbations.
9781405156585_4_009.qxd 11/5/07 14:56 Page 320

Chapter 9 From populations to communities
321
point to the importance of forces that stabilize (density-
dependent factors). At the other extreme, those who
place more emphasis on density fluctuations may
look at external (often density-independent) factors
to explain the changes. Key factor analysis is a
technique that can be applied to life table studies to
throw light both on determination and on regulation of
abundance.
Dispersal, patches and metapopulation
dynamics
Movement can be a vital factor in determining and/or
regulating abundance. A radical change in the way
ecologists think about populations has involved
focusing attention less on processes occurring within
populations and more on patchiness, the colonization
and extinction of subpopulations within an overall meta-
population, and dispersal between subpopulations.
Temporal patterns in community composition
Disturbances that open up gaps (patches) are com-
mon in all kinds of community. Founder-controlled
communities are those in which all species are
approximately equivalent in their ability to invade gaps
and are equal competitors that can hold the gaps
against all comers during their lifetime. Dominance-
controlled communities are those in which some
species are competitively superior to others so that
an initial colonizer of a patch cannot necessarily main-
tain its presence there.
The phenomenon of dominance control is respon-
sible for many examples of community succession.
Primary successions occur in habitats where no
seeds or spores remain from previous occupants
of the site: all colonization must be from outside the
patch. Secondary successions occur when existing
communities are disturbed but some at least of their
seed, etc. remain. It can be very difficult to identify
when a succession reaches a stable climax com-
munity, since this may take centuries to achieve and in
the meantime further disturbances are likely to occur.
The exact nature of the colonization process in an
empty patch depends on the size and location of that
patch. Many communities are mosaics of patches at
different stages in a succession.
Food webs
No predator–prey, parasite–host or grazer–plant pair
exists in isolation. Each is part of a complex food web
involving other predators, parasites, food sources
and competitors within the various trophic levels of a
community.
The effect of one species on another (its herbivor-
ous prey) may be direct and straightforward. But
indirect effects may also be felt by any of the myriad
species linked more remotely in the food web. One of
the most common is a ‘trophic cascade’, in which,
say, a predator reduces the abundance of a herbivore,
thus increasing the abundance of plants.
Top-down control of a food web occurs in situ-
ations in which the structure (abundance, species
number) of lower trophic levels depends on the effects
of consumers from higher trophic levels. Bottom-up
control, on the other hand, occurs in a community
structure dependent on factors, such as nutrient
concentration and prey availability, that influence
a trophic level from below. The relative importance
of these forces varies according to the trophic level
under investigation and the number of trophic levels
present.
Some species are more tightly woven into the food
web than others. A species whose removal would
produce a significant effect (extinction or a large
change in density) in at least one other species may
be thought of as a strong interactor. Removal of some
strong interactors leads to significant changes that
spread throughout the food web; we refer to these as
keystone species.
The relationship between food web complexity and
stability is uncertain (and care is needed in deciding
what is meant by stability). Mathematical and empir-
ical studies agree in suggesting that, if anything,
population stability decreases with complexity, whereas
the stability of aggregate properties of whole com-
munities increases with complexity, especially species
richness.
9781405156585_4_009.qxd 11/5/07 14:56 Page 321

Part III Individuals, Populations, Communities and Ecosystems
322
REVIEW QUESTIONS
Review questions
Asterisks indicate challenge questions
1* Construct a flow diagram (boxes and arrows)
with a named population at its center to
illustrate the wide range of abiotic and biotic
factors that influence its pattern of abundance.
2 Population census data can be used to
establish correlations between abundance and
external factors such as weather. Why can such
correlations not be used to prove a causal
relationship that explains the dynamics of the
population?
3 Distinguish between the determination and
regulation of population abundance.
4* Imagine a number of species with patchy
distributions: a plant, an insect and a mammal
– or consider examples of such species with
which you are familiar. How would you identify
‘habitable patches’ of these species that are
not currently occupied by them?
5 What is meant by a ‘metapopulation’ and how
does it differ from a simple ‘population’?
6 Define founder control and dominance control
as they apply to community organization.
In a mosaic of habitat patches, how would
you expect communities to differ if they were
dominated by founder or dominance control?
7 What factors are responsible for changes
in species composition during an old-field
succession?
8* Draw up a food web of, say, six or seven
species with which you are familiar and
which spans at least three trophic levels.
Take each species in turn and suggest the
kind of community organization that would be
necessary for this to be a keystone species.
9 What are meant by bottom-up and top-down
control? How is the importance of each likely
to vary with the number of trophic levels in a
community?
10 Discuss what is understood about the
relationship between the complexity and
stability of food webs.
9781405156585_4_009.qxd 11/5/07 14:56 Page 322

323
Chapter 10
Patterns in species
richness
CHAPTER CONTENTS
10.1 Introduction
10.2 A simple model of species richness
10.3 Spatially varying factors that influence species richness
10.4 Temporally varying factors that influence species richness
10.5 Gradients of species richness
10.6 Patterns in taxon richness in the fossil record
10.7 Appraisal of patterns in species richness
Chapter contents
KEY CONCEPTS
In this chapter you will:
l
understand the meanings of species richness, diversity indices and
rank–abundance diagrams
l
appreciate that species richness is limited by available resources, the
average portion of the resources used by each species (niche breadth)
and the degree of overlap in resource use
l
recognize that species richness may be highest at intermediate levels of
productivity, predation intensity or disturbance but tends to increase
with spatial heterogeneity
l
appreciate the importance of habitat area and remoteness in
determining richness, especially with reference to the equilibrium
theory of island biogeography
l
understand richness gradients with latitude, altitude and depth, and
during community succession, and the difficulties of explaining them
l
appreciate how theories of species richness can also be applied to the
fossil record
Key concepts
9781405156585_4_010.qxd 11/5/07 14:58 Page 323

10.1 Introduction
Why the number of species varies from place to place, and from time to time,
are questions that present themselves not only to ecologists but to anybody
who observes and ponders the natural world. They are interesting questions in
their own right – but they are also questions of practical importance. It is clear
that if we wish to conserve or restore the planet’s biological diversity, then we
must understand how species numbers are determined and how it comes about
that they vary. We will see that there are plausible answers to the questions we
ask, but these answers are not always clearcut. Yet this is not so much a dis-
appointment as a challenge to ecologists of the future. Much of the fascination
of ecology lies in the fact that many of the problems are blatant, whereas the
solutions can be difficult to find. We will see that a full understanding of patterns
in species richness must draw on our knowledge of all the areas of ecology dis-
cussed so far in this book.
The number of species in a community is referred to as its species richness.
Counting or listing the species present in a community may sound a straight-
forward procedure, but in practice it is often surprisingly difficult, partly because
of taxonomic inadequacies, but also because only a proportion of the organisms
in an area can usually be counted. The number of species recorded then depends
on the number of samples that have been taken, or on the volume of the habitat
that has been explored. The most common species are likely to be represented in
the first few samples, and as more samples are taken, rarer species will be added
to the list. At what point does one cease to take further samples? Ideally, the
investigator should continue to sample until the number of species reaches a
plateau. At the very least, the species richness of different communities should
be compared only if they are based on the same sample sizes (in terms of area of
habitat explored, time devoted to sampling or, best of all, number of individuals
included in the samples).
An important aspect of the structure of a community is completely ignored,
though, when its composition is described simply in terms of the number of species
present – namely, that some species are rare and others common. Intuitively, a
community of 10 species with equal numbers in each seems more diverse than
another, again consisting of 10 species, with 91% of the individuals belonging to
the most common species and just 1% in each of the other nine species. Yet, each
community has the same species richness. Diversity indices are designed to com-
bine both species richness and the evenness or equitability of the distribution
Part III Individuals, Populations, Communities and Ecosystems
324
determining species richness
diversity indices and
rank–abundance diagrams
An accurate appreciation of the world’s biological diversity is becoming
increasingly important. For our conservation efforts to be effective we must
understand why species richness varies widely across the face of the Earth. Why
do some communities contain more species than others? Are there patterns or
gradients in this biodiversity? If so, what are the reasons for these patterns?
9781405156585_4_010.qxd 11/5/07 14:58 Page 324
of individuals among those species (Box 10.1). Moreover, attempts to describe
a complex community structure by one single attribute, such as richness, or even
diversity, can still be criticized because so much valuable information is lost.
A more complete picture of the distribution of species abundance in a community
is therefore sometimes provided in a rank–abundance diagram (Box 10.1).
Chapter 10 Patterns in species richness
325
3
2
1
0
1860 1900
Year
1940
Control
Fertilized
Species diversity (H)
10.1 QUANTITATIVE ASPECTS
10.1 Quantitative aspects
The measure of the character of a community that
is most commonly used to take into account both
species richness and the relative abundances of those
species is known as the Shannon or the Shannon–
Weaver diversity index (denoted by H). This is calculated
by determining, for each species, the proportion of
individuals or biomass (P
i
for the ith species) that that
species contributes to the total in the sample. Then,
if S is the total number of species in the community
(i.e. the richness), diversity (H) is:
H =−∑P
i
ln P
i
where the summation sign ∑ indicates that the
product (P
i
ln P
i
) is calculated for each of the S species
in turn and these products are then summed. As
required, the value of the index depends on both
the species richness and the evenness (equitability)
with which individuals are distributed among the
species. Thus, for a given richness, H increases with
equitability, and for a given equitability, H increases
with richness.
An example of an analysis using diversity indices is
provided by the unusually long-term study that com-
menced in 1856 in an area of pasture at Rothamsted
in England. Experimental plots received a fertilizer
treatment once every year, and control plots did not.
Figure 10.1 shows how species diversity (H) of grass
species changed between 1856 and 1949. While the
unfertilized area remained essentially unchanged,
the fertilized area showed a progressive decline in
diversity. This ‘paradox of enrichment’ is discussed
in Section 10.3.1.
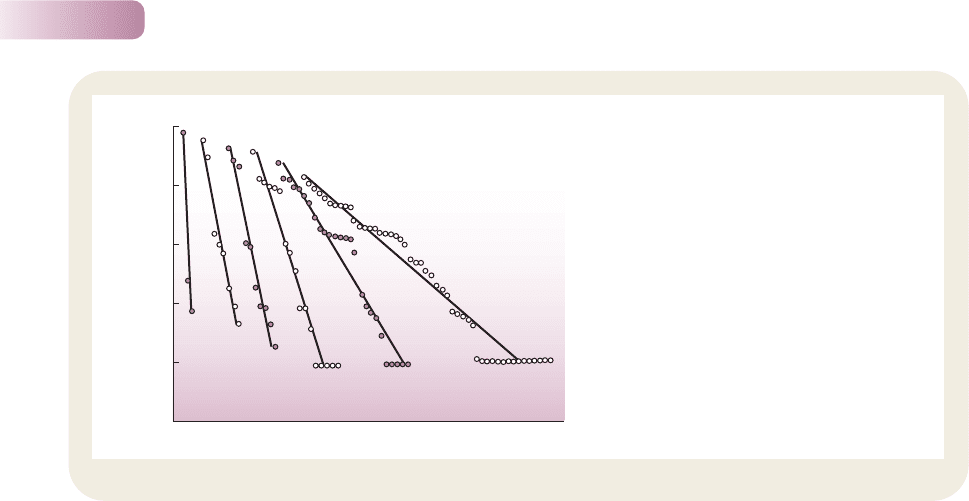
Rank–abundance diagrams, on the other hand,
make use of the full array of P
i
values by plotting P
i
against ‘rank’; i.e. the most abundant species takes
rank 1, the second most abundant rank 2, and so
on, until the array is completed by the rarest species
of all. The steeper the slope of a rank–abundance
diagram, the greater the dominance of common
species over rare species in the community (a steep
slope means a sharp drop in relative abundance,
P
i
, for a given drop in rank). Thus, in the case of the
Rothamsted experiment, Figure 10.2 shows how the
dominance of commoner species steadily increased
(steeper slope) while species richness decreased
over time.
Diversity indices and rank–abundance diagrams
s
Figure 10.1
Species diversity (H) declined progressively in a plot of
pasture that regularly received fertilizer in an experiment
commencing in 1856 at Rothamsted in England. In
contrast, species diversity remained constant in a
control plot that received no fertilizer.
AFTER TILMAN, 1982
9781405156585_4_010.qxd 11/5/07 14:58 Page 325
Nonetheless, for many purposes, the simplest measure, species richness,
suffices. In the following sections, therefore, we examine the relationships between
species richness and a variety of factors that may, in theory, influence richness in
ecological communities. It will become clear that it is not always easy to come up
with unambiguous predictions and clean tests of hypotheses when dealing with
something as complex as a community.
10.2 A simple model of species richness
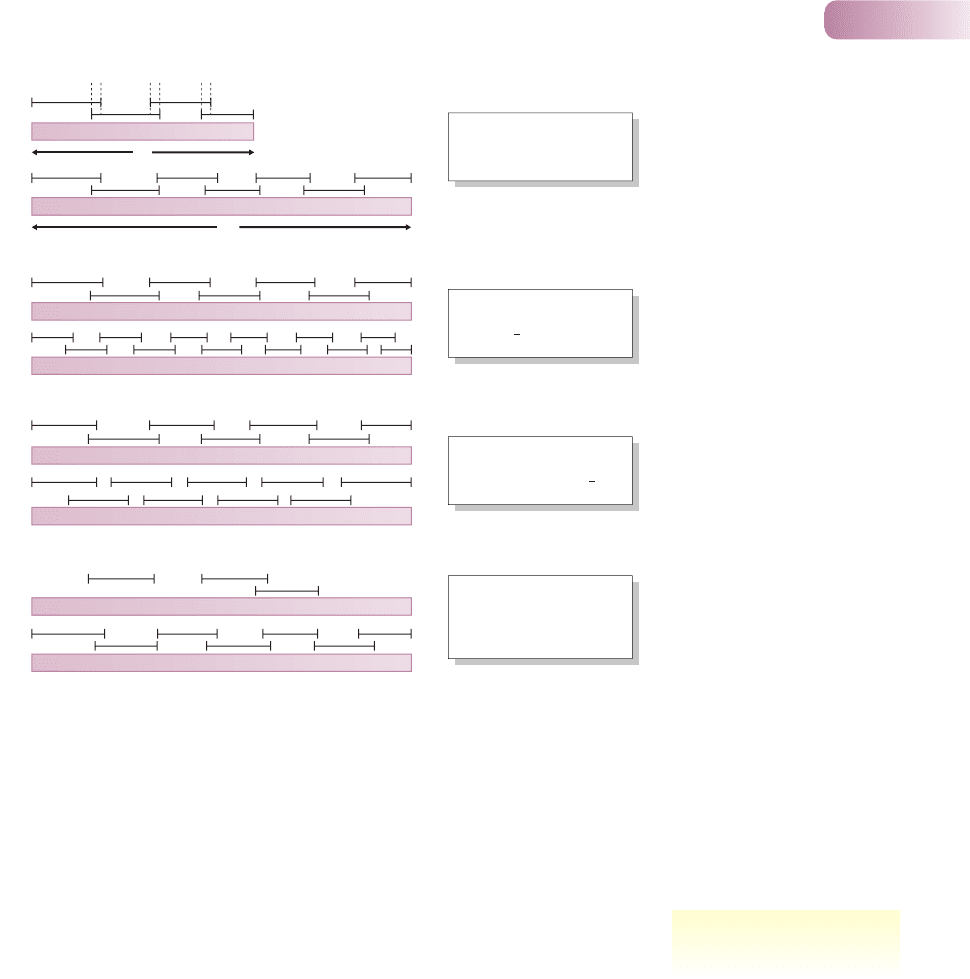
To try to understand the determinants of species richness, it will be useful
to begin with a simple model (Figure 10.3). Assume, for simplicity, that the
resources available to a community can be depicted as a one-dimensional con-
tinuum, R units long. Each species uses only a portion of this resource continuum,
and these portions define the niche breadths (n) of the various species: the
average niche breadth within the community is n¯. Some of these niches overlap,
and the overlap between adjacent species can be measured by value o. The
average niche overlap within the community is then o¯. With this simple back-
ground, it is possible to consider why some communities should contain more
species than others.
First, for given values of n¯ and o¯, a community will contain more species
the larger the value of R, i.e. the greater the range of resources (Figure 10.3a).
Second, for a given range of resources, more species will be accommodated
if n¯ is smaller, i.e. if the species are more specialized in their use of resources
(Figure 10.3b). Alternatively, if species overlap to a greater extent in their use of
resources (greater o¯), then more may coexist along the same resource continuum
(Figure 10.3c). Finally, a community will contain more species the more fully
saturated it is; conversely, it will contain fewer species when more of the resource
continuum is unexploited (Figure 10.3d).
Part III Individuals, Populations, Communities and Ecosystems
326
s
1.0
1949
1919
1903
1872 1862
1856
Relative abundance
Species rank
10
–1
10
–2
10
–3
10
–4
Figure 10.2
Change in the rank–abundance pattern of plant
species in the Rothamstead fertilized plot from 1856
to 1949. Note how the slope of the regression line
becomes progressively steeper with time since
commencement of fertilizer addition. A steeper plot
indicates that the commoner species comprise a
greater proportion of the total community – in other
words, this pasture community gradually became
dominated by just a few species.
AFTER TOKESHI, 1993
9781405156585_4_010.qxd 11/5/07 14:58 Page 326
We can now consider the relationship between this model and two important
kinds of species interactions described in previous chapters: interspecific com-
petition and predation. If a community is dominated by interspecific competition
(see Chapter 6), the resources are likely to be fully exploited. Species richness
will then depend on the range of available resources, the extent to which species
are specialists and the permitted extent of niche overlap (Figure 10.3a–c). We
will examine a range of influences on each of these three.
Predation, on the other hand, is capable of exerting contrasting effects (see
Chapter 7). First, we know that predators can exclude certain prey species; in
the absence of these species the community may then be less than fully saturated,
in the sense that some available resources may be unexploited (Figure 10.3d). In
this way, predation may reduce species richness. Second though, predation may
tend to keep species below their carrying capacities for much of the time, reducing
the intensity and importance of direct interspecific competition for resources.
This may then permit much more niche overlap and a greater richness of species
than in a community dominated by competition (Figure 10.3c).
The next two sections examine a variety of factors that influence species richness.
To organize these, Section 10.3 focuses on factors that often vary spatially (from
place to place): productivity, predation intensity, spatial heterogeneity and envir-
onmental ‘harshness’. Section 10.4 then focuses on factors reflecting temporal
variation: climatic variation, disturbance and evolutionary age.
Chapter 10 Patterns in species richness
327
R
R
n
o
(a)
(b)
(c)
(d)
More species because
greater range of resources
(larger R
)
More species because
each is more specialized
(smaller n
)
More species because
each overlaps more with
its neighbors (larger o)
More species because
resource axis is more fully
exploited (community
more fully saturated)
Figure 10.3
A simple model of species richness.
Each species utilizes a portion n
of the available resources (R),
overlapping with adjacent species
by an amount o. More species may
occur in one community than in
another because: (a) a greater range
of resources is present (larger R),
(b) each species is more specialized
(smaller average n), (c) each species
overlaps more with its neighbors
(larger average o), or (d) the
resource dimension is more fully
exploited.
AFTER MACARTHUR, 1972
competition and predation may
influence species richness
9781405156585_4_010.qxd 11/5/07 14:58 Page 327

10.3 Spatially varying factors that influence
species richness
10.3.1 Productivity and resource richness
For plants, the productivity of the environment can depend on whichever nutri-
ent or condition is most limiting to growth (dealt with in detail in Chapter 11).
Broadly speaking, the productivity of the environment for animals follows the
same trends as for plants, mainly as a result of the changes in resource levels at
the base of the food chain.
If higher productivity is correlated with a wider range of available resources, then
this is likely to lead to an increase in species richness (Figure 10.3a). However,
a more productive environment may have a higher rate of supply of resources
but not a greater variety of resources. This might lead to more individuals per
species rather than more species. Alternatively again, it is possible, even if the
overall variety of resources is unaffected, that rare resources in an unproductive
environment may become abundant enough in a productive environment for
extra species to be added, because more specialized species can be accommodated
(Figure 10.3b).
In general, though, we might expect species richness to increase with
productivity – a contention that is supported by an analysis of the species
richness of trees in North America in relation to a crude measure of available
environmental energy, potential evapotranspiration (PET). This is the amount
of water that under prevailing conditions would evaporate or be transpired from
a saturated surface (Figure 10.4a). However, while energy (heat and light) is
Part III Individuals, Populations, Communities and Ecosystems
328
increased productivity might be
expected to lead to increased
richness...
. . . and often does
160
120
80
40
0
0 600 1200 1800
(a)
(b)
600
100
200
300
400
500
10
20
30
40
50
60
70
200
400
1400
600
800
1000
1200
Species richness
Tree species richness
Potential evapotranspiration (mm yr
–1
)
Rainfall (mm yr
–1
)
Potential evapotranspiration (mm yr
–1
)
Figure 10.4
(a) Species richness of trees in North America (north of the Mexican border) in relation to potential
evapotranspiration. For this analysis the continent was divided into 336 quadrats following lines of latitude
and longitude. (b) Species richness of southern African trees (each dot represents a 25,000 km
2
map
quadrat) in relation to both rainfall and potential evapotranspiration. The three-dimensional surface
describes the regression relationship of species richness with rainfall and potential evapotranspiration.
The surface is divided into zones of increasing depth of color representing increasing species richness.
(a) AFTER CURRIE & PAQUIN, 1987; CURRIE, 1991; (b) DATA FROM O’BRIEN, 1993; AFTER WHITTAKER ET AL., 2003
9781405156585_4_010.qxd 11/5/07 14:58 Page 328