Волькенштейн М.В. Общая биофизика
Подождите немного. Документ загружается.


§
6.4.
ХЕМИОСМОТИЧЕСКОЕ
СОПРЯЖЕНИЕ
301
т.
е.
уже известная формула для равновесия Доннана — Нернста
(см.
стр. 123).
С
перемещением протонов через мембрану связано опреде-
ленное
значение
ее
энергетической емкости,
т. е.
буферного дей-
ствия.
Емкости фаз
ей/
равны
fi
<-
=
d (pH
e
)
'
B
i =
где dH
+
означает число протонов, добавляющихся
в
соответ-
ствующую
фазу. Буферная емкость мембраны
В
определяется
числом протонов ДН
+
, переходящих
из
фазы
е в
фазу
i на
еди-
ницу
понижения рН
е
по отношению
к
рН,-, или
д-
rf
(
AH+
)
(6
29)
"~ rf(ApH)
' ^-
Zy
>
причем
l/B=l/B
e
+l/B
t
.
Если
В
=
const,
то
ДН
+
=
-ВАрН.
(6.30)
Аналогичным образом, электрическая емкость сопрягающей
мембраны равна
г -
d
(Ae+)
(6
3П
где Де
+
есть число положительных зарядов, переносимых из фазы
е
в
фазу
i.
При
С =
const
Ле+
=
СД^.
(6.32)
Работа, совершаемая при генерировании разности
рН
обрати-
мым переносом протонов, равна
-ЛрН
WA
PH
'=
\
ZApHd(AH),
(6.33)
-ЛрН»
где А рН° — равновесное значение Д рН
и
Д рН'
=
Д рН — Д рН°.
При
постоянной буферной емкости
В
Wд
р
н-
=
'/2 ZB {(ДрН)
2
-
(ДрН
0
)
2
}.
(6.34)
Аналогичным образом получаем выражение для работы, совер-
шаемой при генерировании
Д^
обратимым переносом протонов
=
7
2
С
{(Дг|>)
2
-
(Дф°)
2
}.
(6.35)
Если
установлено равновесие
для
протонов,
то
протонодвижу-
щая
сила Д/=
0 (ср.
(6.26)), Д^°
—
ZApH
0
и
полная энергия

302 ГЛ. 6. СОПРЯЖЕННЫЕ ПРОЦЕССЫ В МИТОХОНДРИЯХ
W
Af
хемиосмотической системы, связанная с электрическим и
химическим компонентами этой силы, равна
W*f = Глрн' + WV = '/
2
С (Дг|))
2
+ '/
2
ZB (ДрН)
2
-
-7
2
2(ZC + iS)(ApH
0
)
2
.
(6.36)
Интегрируя от 0 до
—•
АрН', находим выражение WAJ через
ЛрН'
и Дф'= Дф — Дф°:
W
Af
= '/
2
С (Дг|/)
2
+ 7г 2S (АрН')
2
.
(6.37)
Соответственно, уравнение
(6.26)
можно переписать в виде
(6.38)
В митохондриях (из печени крысы) площадь сопрягающей мемб-
раны оценивается в 40 м
2
на 1 г белка [47]. Электрическая ем-
кость при частотах порядка мегациклов составляет около
1
мкФ/см
2
[48]. Отсюда находим С « 0,4 Ф на 1 г белка или
4 мкэкв-В/г белка. Значение В та 12 мкэкв-В/единица рН на
1
г белка. Принимая величину А/ = 270 мВ, получаем W
A
j =
=
0,15 мкэкв -В на 1 г белка при ДрН' = 0 и W
A
i = 7,2 мкэкв- В
на
1 г белка при Дг|/ = 0. Свободная энергия гидролиза
АТФ составляет около 10 ккал/моль или около 0,44 экв-В. Сле-
довательно, W&f соответствует 0,35 мкмоля АТФ на 1 г белка
при
АрН' = 0 и 17 мкмоль АТФ на 1 г белка при Дч|/ = 0. Бо-
лее точные оценки
дают
соответственно 0,24 и 12 мкмоль АТФ
на
1 г белка. Если синтез АТФ сопряжен с переносом
двух
за-
рядов электрона, что
следует
из экспериментально найденного
отношения Ф„/2е^ = 1 для каждого места фосфорилирования, то
необходимое значение А\|/ действительно оказывается не менее
200 мВ, т. е. при толщине мембраны порядка 100 А напряжен-
ность электрического поля в ней не менее 2-Ю
5
В/см. Значение
Дг)/ = 200 мВ эквивалентно вкладу АрН' = 3,5 в общую прото-
нодвижущую силу А/.
Разность потенциалов Дх|) в митохондриальной мембране мо-
жет возникать не только за счет транспорта протонов, но и за
счет транспорта
других
ионов, происходящего одновременно.
Так,
для ионов К
+
в равновесии справедливо соотношение
Нернста
Ai!- = ZApK,
(6.39)
или
(
+
)
D
(
6Ж)
)
где D — эффективный буферный дифференциал для К
+
, опреде-
ляемый как

§
6.4. ХЕМИОСМОТИЧЕСКОЕ СОПРЯЖЕНИЕ 303
Соотношение между Дг|) и полным смещением зарядов, вызван-
ным перемещением всех ионов через мембрану, есть
£d(Ae
+
)
=
Cd(Ai|>),
(6.41)
и
связь между Аг|з и полным перемещением зарядов (за исклю-
чением перемещения, вызванного ионами К
+
) дается выраже-
нием
d(Ae
+
) = Cd(Aifl-d(AK
+
) = Cd(Ai|>) + |-d(A\|,). (6.42)
кроме
К
+
Ионы
К
+
считаются здесь находящимися в равновесии. Величина
С
-\-
DZ~
l
есть полная эффективная емкость. Ее значение может
сильно возрастать, если мембрана проницаема для данного иона.
Если мембрана непроницаема для ионов Н
+
и i порознь, но
возможен их совместный транспорт в одном направлении, т. е.
симпорт
(symport) по Митчеллу, то условие равновесия запи-
шется в виде (v — стехиометрические коэффициенты)
v,Api+v
H
+ApH = 0. (6.43)
Если реализуется антипорт (antiport), т. е. движение ионов i
в направлении, противоположном транспорту Н
+
, то
v.Api
— v
H
+ApH = 0. (6.44)
Симпорт с Н
+
установлен, в частности, для некоторых анионов:
Др (дикарбоксилат
2
") + 2 ДрН = 0,
Др (трикарбоксилат
3
~) + 3 ДрН = 0.
Возможны и более сложные ситуации, например,
Др (фосфат*-) + 2 ДрН + АрК = 0.
Антипорт должен осуществляться для Н
+
и катионов.
Соотношения между ДрН и Лгр можно представить в виде
(см.
стр. 301)
2-
dt " dt '
(6.46)
Суммирование проводится по различным типам реакций, в
ходе
которых происходит транслокация протонов и
других
заряжен-
ных частиц. При постоянных В и С получаем выражения, ана-
логичные (6.30) и (6.32):
" ' ~
b
= CAi|-. (6.47)

304 ГЛ. 6.
СОПРЯЖЕННЫЕ
ПРОЦЕССЫ
В МИТОХОНДРИЯХ
Второе уравнение можно переписать в виде
(6.48)
где Д1
+
— члены суммы Де
+
, отличные от ДН
+
. Из выражений
(6.26),
(6.47)
и
(6.48)
следует
,
Д
Р
Н
.
...
C
-L-
-
(649)
L
ц ~ с + в/z •
уолу)
При
равновесном распределении протонов Д/= 0 и
У
(д/
+
)°
ZApH°
= c + B/Z •
(6
'
50)
Выражение, содержащее лишь отклонения от равновесия, имеет
вид
ZApH'
С— У(Д/
+
)'/Д/
дГ = CTBJZ •
<
6
-
51
)
Таким образом, относительные вклады —ZApH и Лг|) в Д/ зави-
сят лишь от С, В и от числа смещаемых непротонных зарядов.
Вообще говоря, сумма £ (А/
+
)' отрицательна вследствие движе-
ния
ионов через мембрану по электрическому градиенту, т. е.
в направлении, противоположном движению протонов. Эта сум-
ма минимизируется при симпорте протонов и анионов или при
антипорте протонов и катионов. Напротив, увеличение переноса
ионов
в присутствии, скажем, валиномицина (см. стр. 151)
должно увеличивать отрицательное значение 2 Д/
+
и, таким об-
разом, увеличить вклад —ZApH' по сравнению с Д\|/.
Не
ограничиваясь приведенными соотношениями, Митчелл
рассматривает кинетику генерации и затухания ДрН' и A\J/ в не-
которых простейших
случаях.
Цель этого исследования состоит,
в частности, в интерпретации изменений рН, возникающих после
достижения равновесия под влиянием импульса окисления —
восстановления или гидролиза АТФ. Такой импульс может быть
создан добавлением соответствующих веществ.
Полная
протонодвижущая сила Д/ состоит из специфического
потенциала Д\|/, определяемого электрическими свойствами про-
тонов, и разности —ZApH', определяемой их кислотными свой-
ствами. Сила А/ стремится восстановить электрическое и кис-
лотно-основное равновесие. Митчелл предполагает существова-
ние
линейной связи
между
силой и потоком, т. е. рассматривает
близкую к равновесию ситуацию. Полную ионную проводимость,
включающую протонную, можно описать уравнением
^е
+
)
dt
|/
- Z ДрН'),
(6.52)

§
6.4. ХЕМИОСМОТИЧЕСКОЕ СОПРЯЖЕНИЕ 305
где п — число зарядов иона, эффективная протонная проводи-
мость представлена как доля \/п от полной проводимости, г. е.
g =
(l/n)g,
и члены, отвечающие ZgApH для
других
ионов, счи-
таются пренебрежимо малыми. Опыт дает на 1 г митохондри-
ального белка g = 1,7 мкэкв/(В-с). Для протонной проводимо-
сти имеем
d (АН
+
) .
f
1а
к
„,
—j
t
—
= — ё*1- (6.53)
Из
уравнений (6.26), (6.29) и (6.53)
следует,
что
^Р = g (Д*' - Z АрН'), (6.54)
и
из уравнений (6.31) и (6.52) находим
С
d (
У'
]
=g(Z АрН' - п Аг|/). (6.55)
Допустим, что ДрН' и Дгр' генерируются импульсом окисления —
восстановления, причем скорость переноса Н
+
постоянна и равна
Полные
скорости генерации ДрН' и Ai|/ даются выражениями
В
d
У
}
= g(A^-Z АрН') -к, (6.56)
С
d
(
У
7)
=g(Z
АрН'
- п
Аг|)')
+ к.
(6.57)
Решения
этих уравнений при начальных условиях АрН' = Д\|/ =
=
0 при / = 0 имеют вид
2w
(6.58)
где q = Z/B + n/C,
w=^s
2
+ r, s = Z/B — n/C, r =
4Z/CB.
При
стремлении —ZApH' к стационарному значению Ai|/ возра-
стает до максимума, а затем падает до нуля. Если мембрана про-
ницаема
практически лишь для ионов Н+ или ОН"" и g = g или

306
ГЛ.
6.
СОПРЯЖЕННЫЕ ПРОЦЕССЫ
В
МИТОХОНДРИЯХ
п = 1, то решения (6.56) и (6.57) не
могут
быть получены в виде
(6.58) и (6.59). В этом специальном случае уравнения (6.56) и
(6.57) не независимы и их можно переписать в виде
-4-
(
6
-60)
rf(ApH')
It
~~
С d
(Аг|/)
В dt
Интегрирование (6.60) и
(6.61)
дает
Zk
Дф'=-у
С (-Z ДрН').
(6.61)
(6.62)
(6.63)
В этом случае генерация Д\|/ и —ZApH' происходит синхронно.
Время затухания обеих величин до половины исходного значе-
ния
определяется в основном величиной С. Для митохондрий
оно
составляет около 1,5 с. Вообще говоря, временной ход
—Z
АрИ' зависит от значения п, при больших п это время со-
ставляет около 80 с.
Дальнейшие подробности, относящиеся к теории Митчелла,
изложены в работах [42, 44]. На языке линейной неравновесной
термодинамики (см. § 6.3) соответствую-
щие соотношения можно записать на ос-
нове упрощенной схемы, приведенной на
рис.
6.14 [34]. В области / имеем (ср.
стр. 294)
ЛТф
АДФ
у"
—z/
~
H
°
/H
^
e
°'
I
и
в области //
Рис.
6.14. Схема потоков
в
химиосмотической мо-
дели.
(6.64)
(6.65)
В области / окисление вызывает транс-
порт протонов в направлении, противо-
положном АДн, в области // спонтанный поток создает фосфо-
рилирование. Общий поток протонов
1 l' J- I
й
•'
н = J н
-+-
J и.
или
'н
=
(6.66)

§
6.5.
ЭКСПЕРИМЕНТАЛЬНЫЕ
ИССЛЕДОВАНИЯ 307
Основное предположение теории Митчелла состоит в том, что
коэффициент
прямого сопряжения окисления и фосфорилирова-
ния
равен
нулю
(/.ф
О
= 0). Иными словами, если Дрн поддер-
живается постоянным, то фосфорилирование должно быть неза-
висимым от окисления, и наоборот. В частности, при АДн = 0
разобщение полное и /ф = 0, если зФ%— 0 при любом значении
Jo, а также J
o
= 0, если s4-l = 0 при любом значении /
ф
.
Теория
Митчелла — феноменологическая теория, не рассмат-
ривающая механизм процесса и не учитывающая электронно-
конформационные
свойства митохондриальной системы. Тем не
менее,
эта теория позволяет прийти к
ряду
конкретных выводов,
подлежащих прямой экспериментальной проверке.
Вместе
с тем
подлежит проверке и предполагаемая в теории Митчелла бли-
зость процесса к равновесию.
§ 6.5. ЭКСПЕРИМЕНТАЛЬНЫЕ ИССЛЕДОВАНИЯ ХЕМИОСМОТИЧЕСКОГО
СОПРЯЖЕНИЯ
Подробное описание экспериментальных исследований хе-
миосмотического сопряжения дано в монографии Скулачева [46].
Здесь мы приводим лишь краткое изложение основных резуль-
татов.
Прежде всего необходимо было проверить основное положе-
ние
теории Митчелла, согласно которому дыхательная цепь ге-
нерирует трансмембранный потенциал ионов Н
+
. Прямые микро-
электродные измерения не подтвердили предполагаемое в теории
Митчелла высокое сопротивление мембраны
(10
7
—10
8
Ом-см
2
)
и
дали значение мембранного потенциала 10—15 мВ со знаком,
противоположным предполагавшемуся [49]. Однако эти резуль-
таты нельзя считать вполне достоверными (см. [46]). Более на-
дежную
информацию
дает
исследование транспорта специфиче-
ских ионов через мембрану, вызываемого трансмембранным по-
тенциалом.
Валиномицин
вызывает поглощение ионов К
+
энергизован-
ными
митохондриями, т. е. митохондриями, способными к реа-
лизации
сопряжения окисления и фосфорилирования [50]. При
нарушении сопряжения митохондрии теряют накопленные ионы
К
+
.
Эти
результаты
согласуются
с представлениями Митчелла,
но
не доказывают их однозначно [46]. Для обнаружения элек-
трического поля, направленного поперек мембраны энергизован-
ных митохондрий, было необходимо найти ионы, способные про-
никать
через эти мембраны, перенос которых мог бы осуществ-
ляться ферментами — транслоказами без участия электрического
поля.
Либерман и его сотрудники изучили транспорт ряда спе-
циально
синтезированных ионов (синтетические ионы) через ис-
кусственные фосфолипидные мембраны (см. стр. 140) [51, 52].
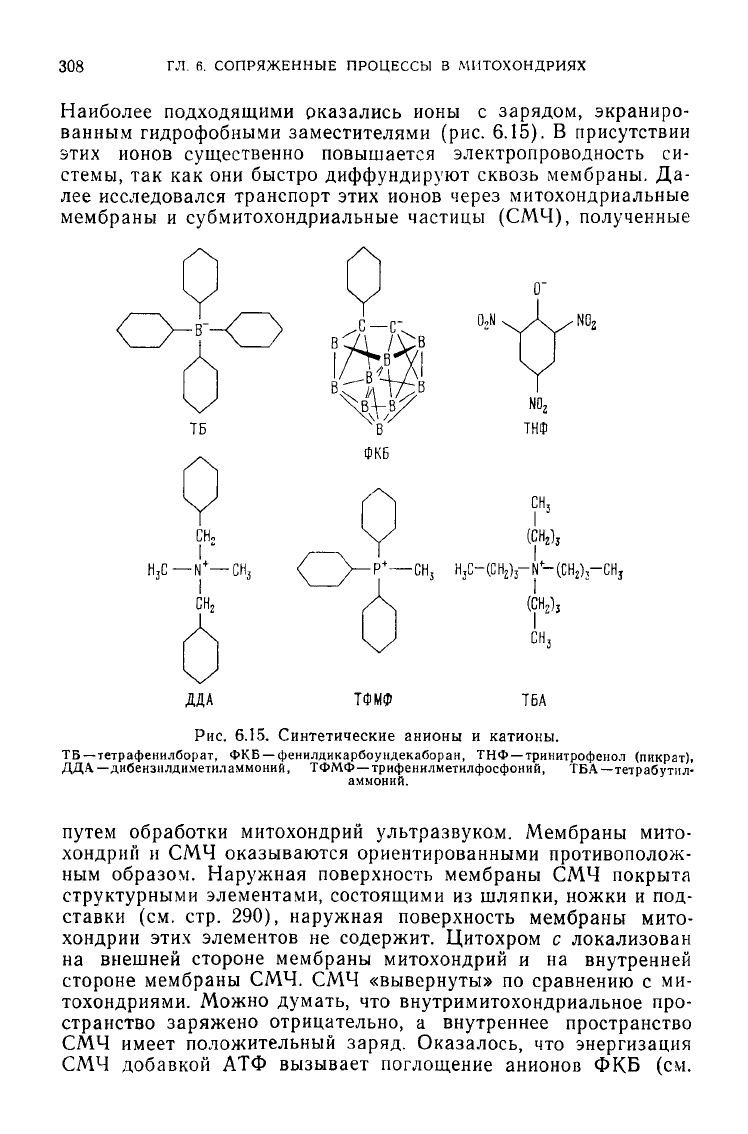
308
ГЛ. 6.
СОПРЯЖЕННЫЕ ПРОЦЕССЫ
В
МИТОХОНДРИЯХ
Наиболее подходящими оказались ионы
с
зарядом, экраниро-
ванным
гидрофобными заместителями
(рис.
6.15).
В
присутствии
этих ионов существенно повышается электропроводность
си-
стемы,
так как они
быстро диффундируют сквозь мембраны.
Да-
лее исследовался транспорт этих ионов через митохондриальные
мембраны
и
субмитохондриальные частицы
(СМЧ),
полученные
0,N
ТНФ
СН
3
Р
+
—СН
3
H
3
C-(CH
2
)
3
-N-(CH
2
)J-CHJ
(СН
2
)
3
сн
3
ТФМФ
TEA
Рис.
6.15. Синтетические анионы и катионы.
ТБ—тетрафеннлборат,
ФКБ —
феннлдикарбоундекаборан,
ТНФ—тринитрофенол (пикрат),
ДДА—дибензилдиметил
аммоний, ТФМФ —трифенилметилфосфоний,
ТБА—тетрабутил-
аммоний.
путем обработки митохондрий ультразвуком. Мембраны мито-
хондрий
и СМЧ
оказываются ориентированными противополож-
ным
образом. Наружная поверхность мембраны
СМЧ
покрыта
структурными элементами, состоящими
из
шляпки, ножки
и под-
ставки
(см. стр. 290),
наружная поверхность мембраны мито-
хондрии этих элементов
не
содержит. Цитохром
с
локализован
на
внешней стороне мембраны митохондрий
и на
внутренней
стороне мембраны СМЧ.
СМЧ
«вывернуты»
по
сравнению
с ми-
тохондриями. Можно
думать,
что
внутримитохондриальное
про-
странство заряжено отрицательно,
а
внутреннее пространство
СМЧ
имеет положительный заряд. Оказалось,
что
энергизация
СМЧ
добавкой
АТФ
вызывает поглощение анионов
ФКБ (см.

§
6.5. ЭКСПЕРИМЕНТАЛЬНЫЕ ИССЛЕДОВАНИЯ 309
рис.
6.15),
а деанергизация ингибитором дыхания (антимици-
ном) или разобщителем окислительного фосфорилирования ОКФ
(rz-трифторметоксикарбонилцианидфенилгидразон) вызывает вы-
ход анионов. В опытах с митохондриями наблюдается иная кар-
тина [46, 53—56].
Удалось
показать, что транспорт электронов
в мембранах СМЧ сопровождается поглощением анионов ФКБ
[55, 57]. Транспорт ФКБ нарушается ингибиторами электронного
транспорта, а также разобщителями окислительного фосфори-
лирования.
Аналогичные
результаты
получены и с другими синтетиче-
скими
анионами [55, 58, 59].
В целом указанные опыты с синтетическими ионами, а так-
же ряд
других,
доказывают, что в сопрягающих мембранах реа-
лизуется энергозависимый транспорт ионов, причем анионы и
катионы
движутся
в противоположных направлениях. Механизм
транспорта не зависит от
структуры
ионов, существенна лишь
их способность проникать через фосфолипидные мембраны. Из-
менение ориентации мембраны относительно окружающей среды
приводит к переориентации ионных потоков, направленных про-
тив градиентов концентрации ионов [46].
Эти электрофоретические свойства мембран нельзя объяс-
нить высказанными ранее представлениями об ответственности
специфического фермента — энергозависимой транслоказы — за
транспорт ионов (см., например, [60]). Напротив, эти свойства
согласуются
с теорией Митчелла, исходящей из того, что хими-
ческая энергия реакции окисления и фосфорилирования может
превращаться в электрическую энергию мембранного потен-
циала.
Согласно теории Митчелла (см. § 6.4), перенос протонов и
электронов сквозь мембрану не приводит к большим изменениям
рН.
Транспорт электронов, т. е. окислительный процесс, прекра-
щается под действием электрического поля, создаваемого избыт-
ком
отрицательных зарядов на
другой
стороне мембраны. Пере-
нос
электронов активируется вновь при уменьшении поля, т. е.
при
уменьшении мембранного потенциала. Этого можно достичь
перемещением катионов через мембрану. В
результате
должен
возникнуть градиент рН, так как перенос каждого одновалент-
ного катиона должен сопровождаться освобождением одного
протона во внешней среде. В
работах
[3,
53—55]
действительно
показано,
что добавление проникающих синтетических катионов
ДДА,
ТФМФ,
ТБА (см. рис. 6.15) к митохондриям вызывает уве-
личение концентрации Н
+
в среде. Изменения рН исчезают при
добавке разобщителей. Сам Митчелл показал, что при работе
дыхательной цепи среда подкисляется [42, 43].
Таким образом, установлено, что энергизация сопрягающей
мембраны создает трансмембранный потенциал Д\|з и ДрН.

310 ГЛ. 6.
СОПРЯЖЕННЫЕ
ПРОЦЕССЫ
В
МИТОХОНДРИЯХ
Из
теории Митчелла
следует,
что энергия дыхания, транс-
формированная
из химической формы в электрическую и осмо-
тическую, может вновь перейти в химическую форму при син-
тезе
АТФ или обратном переносе электронов. Митчелл посту-
лирует,
тем самым, обратимость биоэнергетического процесса.
Это положение также подтверждено (см. [44]). Реализуется так
называемое ионное фосфорилирование — синтез АТФ из АДФ
и
Ф„ за
счет
электрической или осмотической энергии, выде-
ляемой при движении ионов через мембрану по концентрацион-
ному
градиенту.
Хорошими тестами на трансмембранный потенциал являют-
ся
изменения спектров поглощения цитохромов, вызываемые их
окислением
и восстановлением [61, 62]. При исследованиях по-
ведения цитохромов в СМЧ может оказаться весьма полезным
магнитооптическое вращение, аномальная дисперсия которого
очень чувствительна к изменению состояния группы гема, со-
держащейся в цитохромах (см. [2], гл. 7).
Удается
наблюдать
такую
аномальную дисперсию для цитохрома с во взвеси СМЧ.
Снижение
А*ф должно приводить к нарушению сопряжения
окисления
и фосфорилирования. К этому сводится действие раз-
общителей сопряжения. Таковыми являются динитрофенол
ДНФ,
ФКФ и ряд дру-
Н
+
/ Мембрана
f
он" гих
веществ.
ДНФ, по-
видимому,
действует
как
переносчик протонов,
со-
•
Н
+
гласно
схеме
Митчелла,
показанной
на рис. 6.16.
Очевидно,
что
если
эта
схема верна,
то
любые
Рис.
6.16.
Транспорт протонов разобщите-
СЛабые
КИ
СЛОТЫ
и
осно-
лем
ДНФ.
вания должны оказывать
разобщающее действие.
Установлено,
что
разобщители действительно повышают
протонную проводимость мембран митохондрий
[63].
Сход-
ные результаты были получены
и на
искусственных фосфо-
липидных мембранах
[53,
64—66]. Выявлено далеко идущее
сходство этих мембран
с
внутренними мембранами мито-
хондрий. Найден новый класс сильных разобщителей
—
произ-
водных барена
[52]
(дальнейшие подробности
см. в
[46]).
По
схеме
Митчелла
[42, 43]
протоны
и
электроны шесть
раз
пересекают мембрану
на
пути
от
НАД-Н
к О
2
.
Стадии переноса
Н
+
и е~
чередуются.
Для
детального рассмотрения
и
аргумен-
тации этой схемы нужно иметь представление
о
локализации
компонентов реакции. Установлено,
что
цитохром
с
локализо-
ван
у
внешней стороны мембраны митохондрий,
а
ферменты
трансгидрогеназа, НАД-Н-дегидрогеназа
и
сукцинатдегидроге-