Berg J.M., Tymoczko J.L., Stryer L. Biochemistry
Подождите немного. Документ загружается.

I. The Molecular Design of Life 12. Lipids and Cell Membranes 12.4. Phospholipids and Glycolipids Readily Form Bimolecular Sheets in Aqueous Media
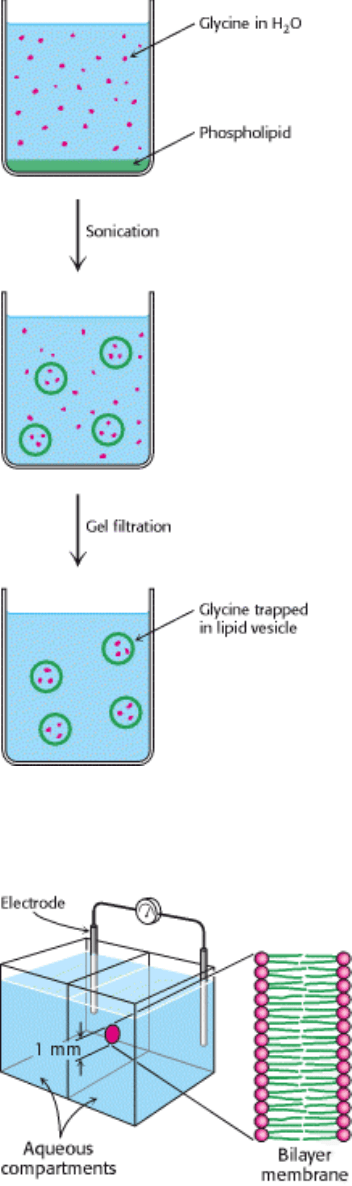
Figure 12.13. Preparation of Glycine-Containing Liposome. Liposomes containing glycine are formed by sonication
of phospholipids in the presence of glycine. Free glycine is removed by gel filtration.
I. The Molecular Design of Life 12. Lipids and Cell Membranes 12.4. Phospholipids and Glycolipids Readily Form Bimolecular Sheets in Aqueous Media
Figure 12.14. Experimental Arrangement for the Study of Planar Bilayer Membrane. A bilayer membrane is
formed across a 1-mm hole in a septum that separates two aqueous compartments. This arrangement permits
measurements of the permeability and electrical conductance of lipid bilayers.

I. The Molecular Design of Life 12. Lipids and Cell Membranes 12.4. Phospholipids and Glycolipids Readily Form Bimolecular Sheets in Aqueous Media
Figure 12.15. Permeability Coefficients (P) of Ions and Molecules in a Lipid Bilayer. The ability of molecules to
cross a lipid bilayer spans a wide range of values.
I. The Molecular Design of Life 12. Lipids and Cell Membranes
12.5. Proteins Carry Out Most Membrane Processes
We now turn to membrane proteins, which are responsible for most of the dynamic processes carried out by membranes.
Membrane lipids form a permeability barrier and thereby establish compartments, whereas specific proteins mediate
nearly all other membrane functions. In particular, proteins transport chemicals and information across a membrane.
Membrane lipids create the appropriate environment for the action of such proteins.
Membranes differ in their protein content. Myelin, a membrane that serves as an insulator around certain nerve fibers,
has a low content of protein (18%). Relatively pure lipids are well suited for insulation. In contrast, the plasma
membranes or exterior membranes of most other cells are much more active. They contain many pumps, channels,
receptors, and enzymes. The protein content of these plasma membranes is typically 50%. Energy-transduction
membranes, such as the internal membranes of mitochondria and chloroplasts, have the highest content of protein,
typically 75%.
The protein components of a membrane can be readily visualized by SDS-polyacrylamide gel electrophoresis. As
discussed earlier (Section 4.1.4), the electrophoretic mobility of many proteins in SDS-containing gels depends on the
mass rather than on the net charge of the protein. The gel-electrophoresis patterns of three membranes the plasma
membrane of erythrocytes, the photoreceptor membrane of retinal rod cells, and the sarcoplasmic reticulum membrane of
muscle are shown in Figure 12.16. It is evident that each of these three membranes contains many proteins but has a
distinct protein composition. In general, membranes performing different functions contain different repertoires of
proteins.
12.5.1. Proteins Associate with the Lipid Bilayer in a Variety of Ways
The ease with which a protein can be dissociated from a membrane indicates how intimately it is associated with the
membrane. Some membrane proteins can be solubilized by relatively mild means, such as extraction by a solution of
high ionic strength (e.g., 1 M NaCl). Other membrane proteins are bound much more tenaciously; they can be
solubilized only by using a detergent or an organic solvent. Membrane proteins can be classified as being either
peripheral or integral on the basis of this difference in dissociability (Figure 12.17). Integral membrane proteins interact
extensively with the hydrocarbon chains of membrane lipids, and so only agents that compete for these nonpolar
interactions can release them. In fact, most integral membrane proteins span the lipid bilayer. In contrast, peripheral
membrane proteins are bound to membranes primarily by electrostatic and hydrogen-bond interactions with the head
groups of lipids. These polar interactions can be disrupted by adding salts or by changing the pH. Many peripheral
membrane proteins are bound to the surfaces of integral proteins, on either the cytosolic or the extracellular side of the
membrane. Others are anchored to the lipid bilayer by a covalently attached hydrophobic chain, such as a fatty acid.
12.5.2. Proteins Interact with Membranes in a Variety of Ways

Although membrane proteins are more difficult to purify and crystallize than are water-soluble proteins, researchers
using x-ray crystallographic or electron microscopic methods have determined the three-dimensional structures of more
than 20 such proteins at sufficiently high resolution to discern the molecular details. As noted in Chapter 3, the structures
of membrane proteins differ from those of soluble proteins with regard to the distribution of hydrophobic and
hydrophilic groups. We will consider the structures of three membrane proteins in some detail.
Proteins Can Span the Membrane with Alpha Helices.
The first membrane protein that we consider is the archaeal protein bacteriorhodopsin, shown in Figure 12.18. This
protein acts in energy transduction by using light energy to transport protons from inside the cell to outside. The proton
gradient generated in this way is used to form ATP. Bacteriorhodopsin is built almost entirely of α helices; seven closely
packed α helices, arranged almost perpendicularly to the plane of the membrane, span its 45-Å width. Examination of
the primary structure of bacteriorhodopsin reveals that most of the amino acids in these membrane-spanning α helices
are nonpolar and only a very few are charged (Figure 12.19). This distribution of nonpolar amino acids is sensible
because these residues are either in contact with the hydrocarbon core of the membrane or with one another. Membrane-
spanning α helices are the most common structural motif in membrane proteins. As will be discussed in Section 12.5.4,
such regions can often be detected from amino acid sequence information alone.
A Channel Protein Can Be Formed from Beta Strands.
Porin, a protein from the outer membranes of bacteria such as E. coli and Rhodo-bacter capsulatus, represents a class of
membrane proteins with a completely different type of structure. Structures of this type are built from β strands and
contain essentially no α helices (Figure 12.20).
The arrangement of β strands is quite simple: each strand is hydrogen bonded to its neighbor in an antiparallel
arrangement, forming a single β sheet. The β sheet curls up to form a hollow cylinder that functions as the active unit. As
its name suggests, porin forms pores, or channels, in the membranes. A pore runs through the center of each cylinder-like
protein. The outside surface of porin is appropriately nonpolar, given that it interacts with the hydrocarbon core of the
membrane. In contrast, the inside of the channel is quite hydrophilic and is filled with water. This arrangement of
nonpolar and polar surfaces is accomplished by the alternation of hydrophobic and hydrophilic amino acids along each β
strand (Figure 12.21).
Embedding Part of a Protein in a Membrane Can Link It to the Membrane Surface.
The structure of the membrane-bound enzyme prostaglandin H
2
synthase-1 reveals a rather different role for α helices in
protein-membrane associations. This enzyme catalyzes the conversion of arachidonic acid into prostaglandin H
2
in two
steps: a cyclooxygenase reaction and a peroxidase reaction (Figure 12.22). Prostaglandin H
2
promotes inflammation and
modulates gastric acid secretion. The enzyme that produces prostaglandin H
2
is a homodimer with a rather complicated
structure consisting primarily of α helices. Unlike bacteriorhodopsin, this protein is not largely embedded in the
membrane but, instead, lies along the outer surface of the membrane firmly bound by a set of α helices with hydrophobic
surfaces that extend from the bottom of the protein into the membrane (Figure 12.23). This linkage is sufficiently strong
that only the action of detergents can release the protein from the membrane. Thus, this enzyme is classified as an
integral membrane protein, although it is not a membrane-spanning protein.
The localization of prostaglandin H
2
synthase-l in the membrane is crucial to its function. The substrate for this
enzyme, arachidonic acid, is a hydrophobic molecule generated by the hydrolysis of membrane lipids.
Arachidonic acid reaches the active site of the enzyme from the membrane without entering an aqueous environment by
traveling through a hydrophobic channel in the protein (Figure 12.24). Indeed, nearly all of us have experienced the
importance of this channel: drugs such as aspirin and ibuprofen block the channel and prevent prostaglandin synthesis by
inhibiting the cyclooxygenase activity of the synthase. In the case of aspirin, the drug acts through the transfer of an
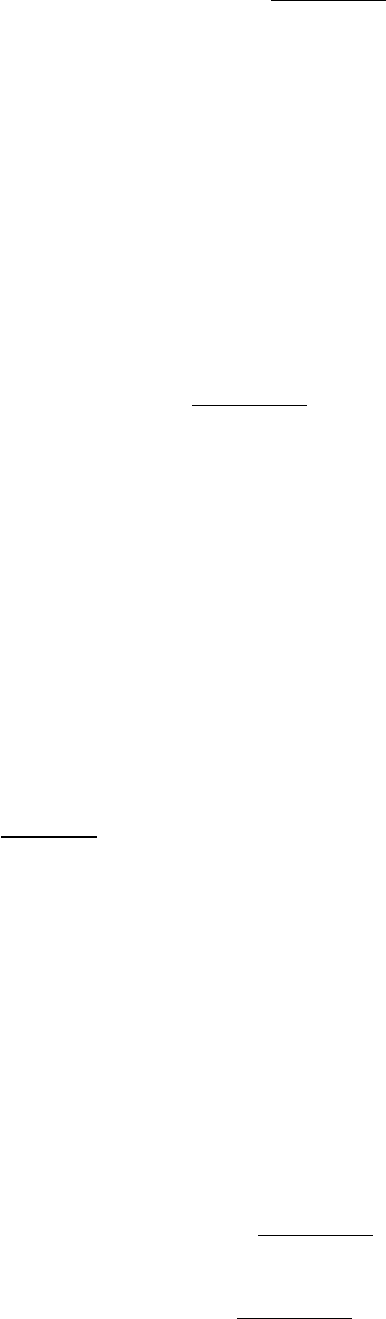
acetyl group from the aspirin to a serine residue (Ser 530) that lies along the path to the active site (Figure 12.25).
Two important features emerge from our examination of these three examples of membrane protein structure. First, the
parts of the protein that interact with the hydrophobic parts of the membrane are coated with nonpolar amino acid side
chains, whereas those parts that interact with the aqueous environment are much more hydrophilic. Second, the
structures positioned within the membrane are quite regular and, in particular, all backbone hydrogen-bond donors and
acceptors participate in hydrogen bonds. Breaking a hydrogen bond within a membrane is quite unfavorable, because
little or no water is present to compete for the polar groups.
12.5.3. Some Proteins Associate with Membranes Through Covalently Attached
Hydrophobic Groups
The membrane proteins considered thus far associate with the membrane through surfaces generated by hydrophobic
amino acid side chains. However, even otherwise soluble proteins can associate with membranes if the association is
mediated by hydrophobic groups attached to the proteins. Three such groups are shown in Figure 12.26: (1) a palmitoyl
group attached to a specific cysteine residue by a thioester bond, (2) a farnesyl group attached to a cysteine residue at the
carboxyl terminus, and (3) a glycolipid structure termed a glycosyl phosphatidyl inositol (GPI) anchor attached to the
carboxyl terminus. These modifications are attached by enzyme systems that recognize specific signal sequences near
the site of attachment.
12.5.4. Transmembrane Helices Can Be Accurately Predicted from Amino Acid
Sequences
Many membrane proteins, like bacteriorhodopsin, employ α helices to span the hydrophobic part of a membrane. As
noted earlier, typically most of the residues in these α helices are nonpolar and almost none of them are charged. Can we
use this information to identify putative membrane-spanning regions from sequence data alone? One approach to
identifying transmembrane helices is to ask whether a postulated helical segment is likely to be most stable in a
hydrocarbon milieu or in water. Specifically, we want to estimate the free-energy change when a helical segment is
transferred from the interior of a membrane to water. Free-energy changes for the transfer of individual amino acid
residues from a hydrophobic to an aqueous environment are given in Table 12.2. For example, the transfer of a poly-l-
arginine helix, a homopolymer of a positively charged amino acid, from the interior of a membrane to water would be
highly favorable [-12.3 kcal mol
-1
(-51.5 kJ mol
-1
) per arginine residue in the helix], whereas the transfer of a poly-l-
phenylalanine helix, a homopolymer of a hydrophobic amino acid, would be unfavorable [+3.7 kcal mol
-1
(+15.5 kJ mol
-
1
) per phenylalanine residue in the helix].
The hydrocarbon core of a membrane is typically 30 Å wide, a length that can be traversed by an α helix consisting of 20
residues. We can take the amino acid sequence of a protein and estimate the free-energy change that takes place when a
hypothetical α helix formed of residues 1 through 20 is transferred from the membrane interior to water. The same
calculation can be made for residues 2 through 21, 3 through 22, and so forth, until we reach the end of the sequence.
The span of 20 residues chosen for this calculation is called a window. The free-energy change for each window is
plotted against the first amino acid at the window to create a hydropathy plot. Empirically, a peak of +20 kcal mol
-1
(+84
kJ mol
-1
) or more in a hydropathy plot based on a window of 20 residues indicates that a polypeptide segment could be a
membrane-spanning α helix. For example, glycophorin, a protein found in the membranes of red blood cells, is predicted
by this criterion to have one membrane-spanning helix, in agreement with experimental findings (Figure 12.27). It
should be noted, however, that a peak in the hydropathy plot does not prove that a segment is a transmembrane helix.
Even soluble proteins may have highly nonpolar regions. Conversely, some membrane proteins contain transmembrane
structural features (such as membrane-spanning β strands) that escape detection by these plots (Figure 12.28).
I. The Molecular Design of Life 12. Lipids and Cell Membranes 12.5. Proteins Carry Out Most Membrane Processes
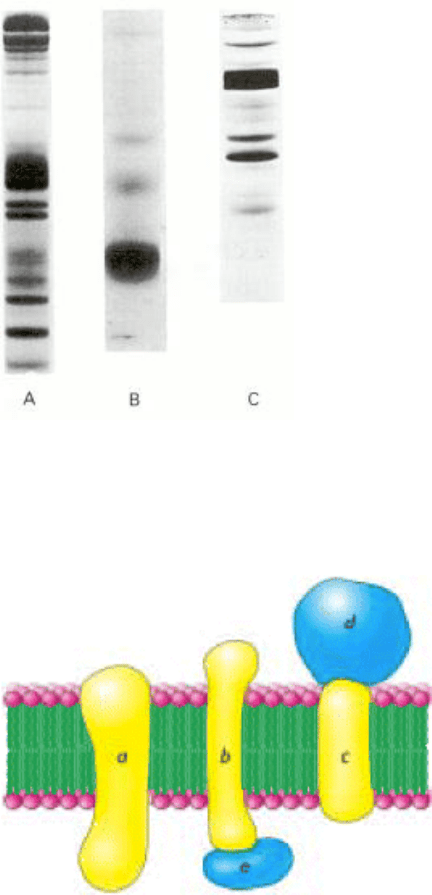
Figure 12.16. Sds-Acrylamide Gel Patterns of Membrane Proteins. (A) The plasma membrane of erythrocytes. (B)
The photoreceptor membranes of retinal rod cells. (C) The sarcoplasmic reticulum membrane of muscle cells. [Courtesy
of Dr. Theodore Steck (Part A) and Dr. David MacLennan (Part C).]
I. The Molecular Design of Life 12. Lipids and Cell Membranes 12.5. Proteins Carry Out Most Membrane Processes
Figure 12.17. Integral and Peripheral Membrane Proteins. Integral membrane proteins (a, b, and c) interact
extensively with the hydrocarbon region of the bilayer. Nearly all known integral membrane proteins traverse the lipid
bilayer. Peripheral membrane proteins (d and e) bind to the surfaces of integral proteins. Some peripheral membrane
proteins interact with the polar head groups of the lipids (not shown).
I. The Molecular Design of Life 12. Lipids and Cell Membranes 12.5. Proteins Carry Out Most Membrane Processes
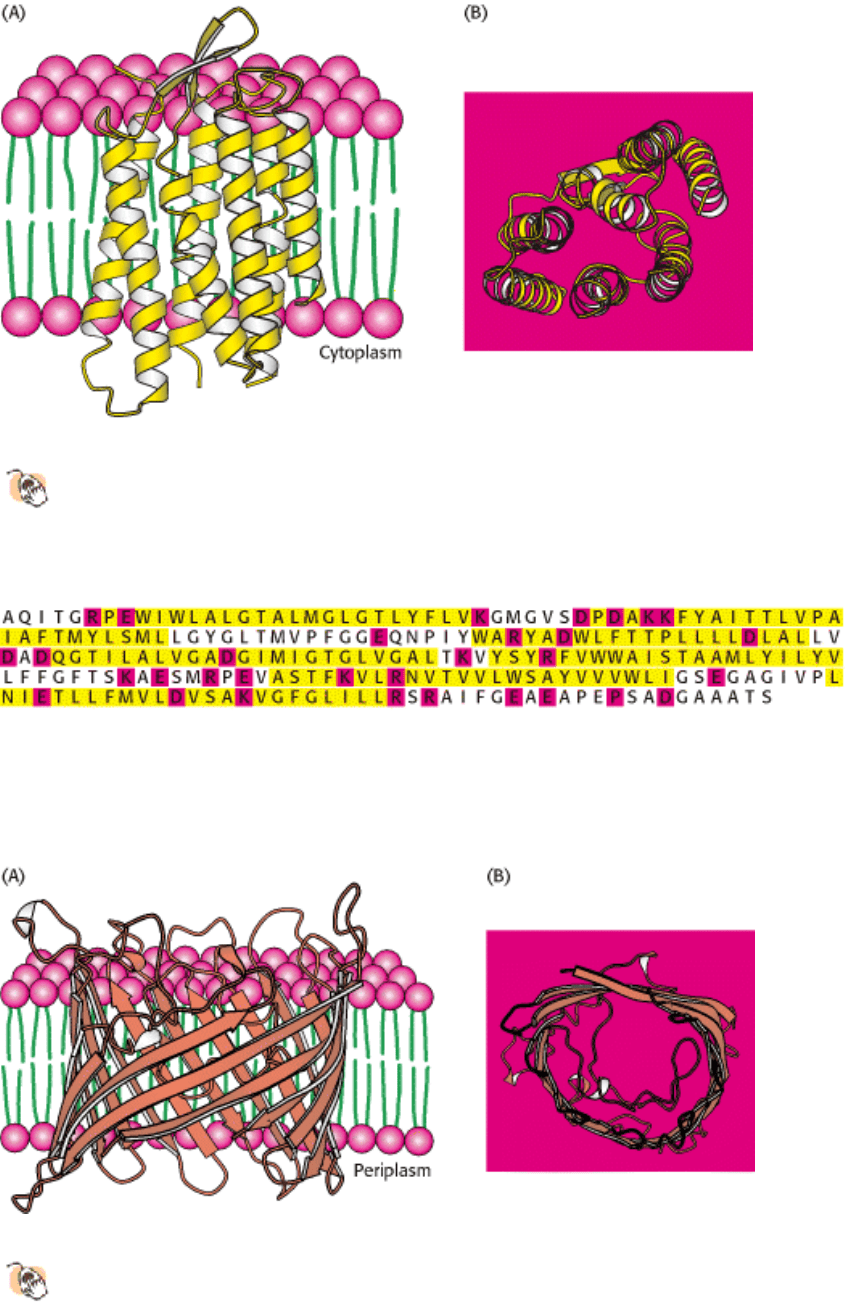
Figure 12.18. Structure of Bacteriorhodopsin.
Bacteriorhodopsin consists largely of membrane-spanning α helices.
(A) View through the membrane bilayer. The interior of the membrane is green and the head groups are red. (B) A
view from the cytoplasmic side of the membrane.
I. The Molecular Design of Life 12. Lipids and Cell Membranes 12.5. Proteins Carry Out Most Membrane Processes
Figure 12.19. Amino Acid Sequence of Bacteriorhodopsin. The seven helical regions are highlighted in yellow and
the charged residues in red.
I. The Molecular Design of Life 12. Lipids and Cell Membranes 12.5. Proteins Carry Out Most Membrane Processes
Figure 12.20. Structure of Bacterial Porin (from Rhodopseudomonas blastica).
Porin is a membrane protein built
entirely of beta strands. (A) Side view. (B) View from the periplasmic space. Only one monomer of the trimeric
protein is shown.
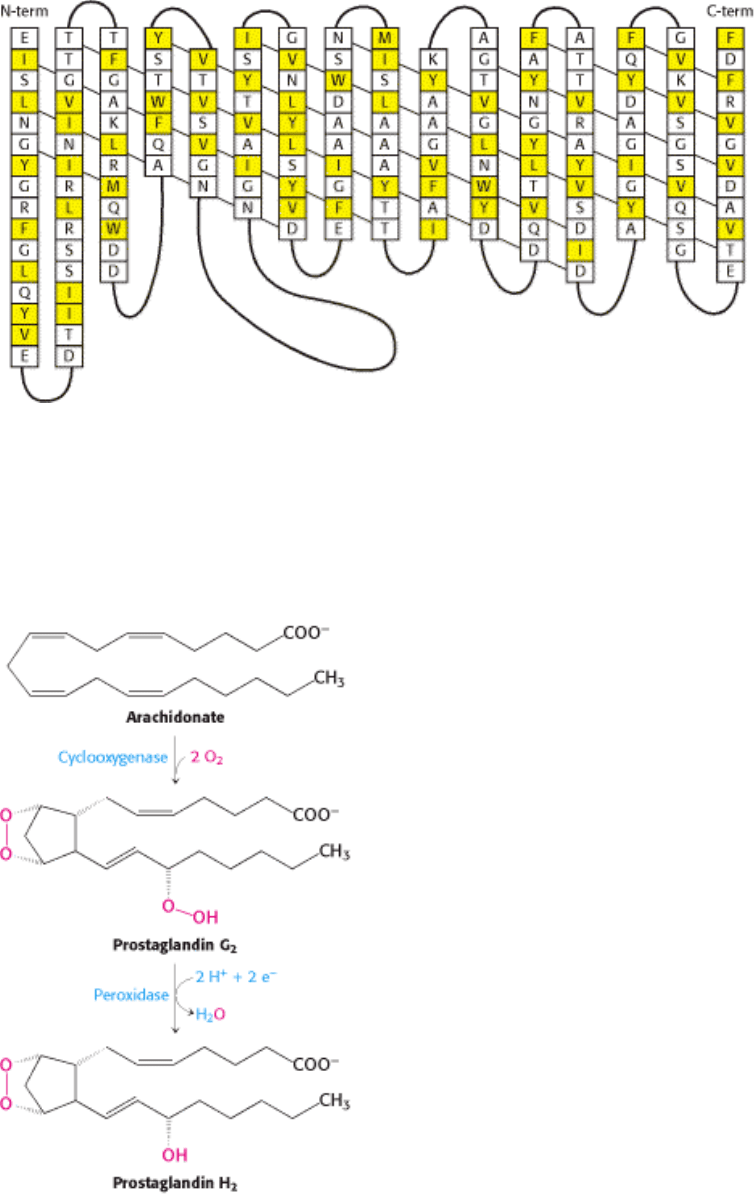
I. The Molecular Design of Life 12. Lipids and Cell Membranes 12.5. Proteins Carry Out Most Membrane Processes
Figure 12.21. Amino Acid Sequence of a Porin. Some membrane proteins such as porins are built from β strands that
tend to have hydrophobic and hydrophilic amino acids in adjacent positions. The secondary structure of
Rhodopseudomonas blastica is shown, with the diagonal lines indicating the direction of hydrogen bonding along the β
sheet. Hydrophobic residues (F, I, L, M, V, W, and Y) are shown in yellow. These residues tend to lie on the outside of
the structure, in contact with the hydrophobic core of the membrane.
I. The Molecular Design of Life 12. Lipids and Cell Membranes 12.5. Proteins Carry Out Most Membrane Processes
Figure 12.22. Formation of Prostaglandin H
2
. Prostaglandin H
2
synthase-1 catalyzes the formation of prostaglandin
H
2
from arachidonic acid in two steps.
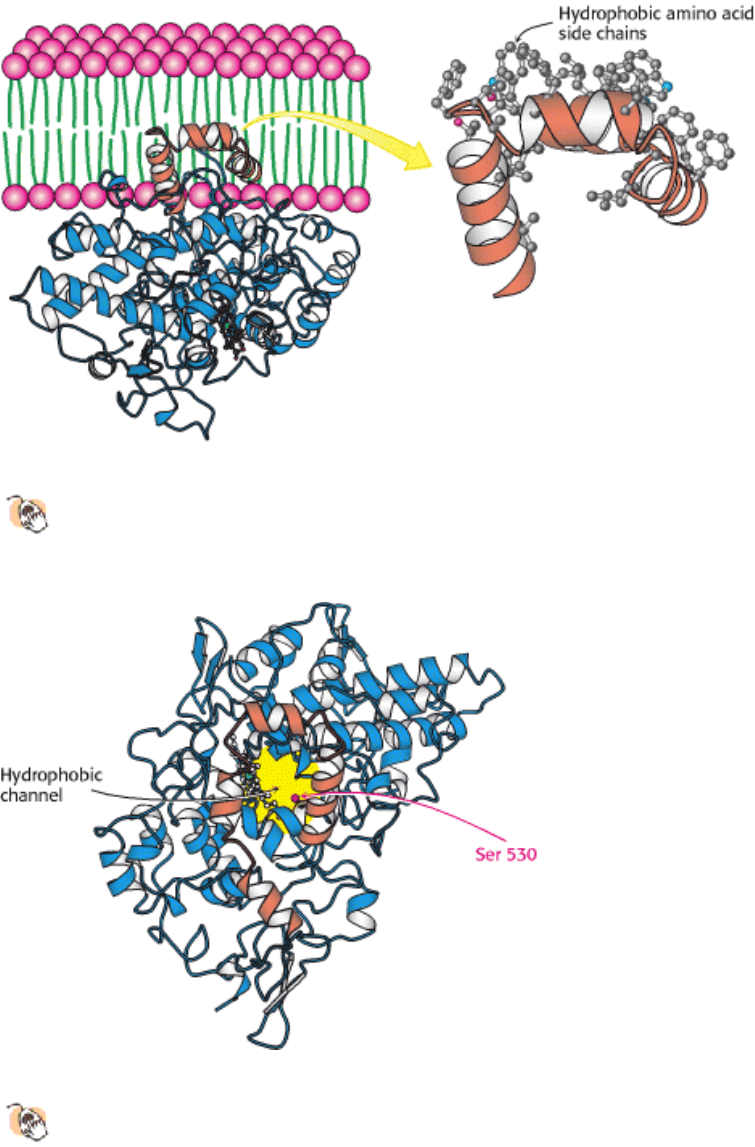
I. The Molecular Design of Life 12. Lipids and Cell Membranes 12.5. Proteins Carry Out Most Membrane Processes
Figure 12.23. Attachment of Prostaglandin H
2
Synthase-1 to the Membrane. Prostaglandin H
2
synthase-1 is held in
the membrane by a set of α helices coated with hydrophobic side chains. One monomer of the dimeric enzyme is
shown.
I. The Molecular Design of Life 12. Lipids and Cell Membranes 12.5. Proteins Carry Out Most Membrane Processes
Figure 12.24. HYdrophobic Channel of Prostaglandin H
2
Synthase. A view of prostaglandin H
2
synthase from the
membrane, showing the hydrophobic channel that leads to the active site. The membrane-anchoring helices are
shown in orange.

I. The Molecular Design of Life 12. Lipids and Cell Membranes 12.5. Proteins Carry Out Most Membrane Processes
Figure 12.25. Aspirin's Effects On Prostaglandin H
2
Synthase-1. Aspirin acts by transferring an acetyl group to a
serine residue in prostaglandin H
2
synthase-1.
I. The Molecular Design of Life 12. Lipids and Cell Membranes 12.5. Proteins Carry Out Most Membrane Processes
Figure 12.26. Membrane Anchors. Membrane anchors are hydrophobic groups that are covalently attached to proteins
(in blue) and tether the proteins to the membrane. The red and blue hexagons correspond to mannose and GlcNAc,
respectively. R groups represent points of additional modification.

I. The Molecular Design of Life 12. Lipids and Cell Membranes 12.5. Proteins Carry Out Most Membrane Processes
Table 12.2. Polarity scale for identifying transmembrane helices
Amino acid residue
Transfer free energy kcal mol
-1
(kJ mol
-1
)
Phe 3.7 (15.5)
Met 3.4 (14.3)
Ile 3.1 (13.0)
Leu 2.8 (11.8)
Val 2.6 (10.9)
Cys 2.0 (8.4)
Trp 1.9 (8.0)
Ala 1.6 (6.7)
Thr 1.2 (5.0)
Gly 1.0 (4.2)
Ser 0.6 (2.5)
Pro -0.2 (-0.8)
Tyr -0.7 (-2.9)
His -3.0 (-12.6)
Gln -4.1 (-17.2)
Asn -4.8 (-20.2)
Glu -8.2 (-34.4)
Lys -8.8 (-37.0)
Asp -9.2 (-38.6)
Arg -12.3 (-51.7)
Source: After D. M. Engelman, T. A. Steitz, and A. Goldman. Annu. Rev. Biophys. Biophys. Chem. 15(1986):330.
Note: The free energies are for the transfer of an amino acid residue in an α helix from the membrane interior (assumed to have a
dielectric constant of 2) to water.
I. The Molecular Design of Life 12. Lipids and Cell Membranes 12.5. Proteins Carry Out Most Membrane Processes