Елинов Н.П. Основы биотехнологии
Подождите немного. Документ загружается.


Некоторые из нуклеотидных фрагментов входят не только в
состав интронов, или фланкирующих (от англ. flank — бок,
сторона, прикрывать с фланга) последовательностей мРНК, но и
могут выполнять функции сигнальных последовательностей,
узнаваемых белками (промоторы транскрипции, точки начала ре-
пликации ДНК, сайты скручивания хромосом и др.). Все эти
сигнальные последовательности, повторяющиеся в геноме в виде
идентичных или сходных тандемных (от англ. tandem — гуськом,
цугом) копий, имеют небольшую длину. При разделении ДНК в
градиенте плотности хлористого цезия сигнальные последователь-
ности (порядка 5%) удается отделять от основной массы ДНК в
виде сателлитной ДНК (дополнительный(е) пик(и) при центрифу-
гировании). Эти ДНК могут быть тяжелее или легче основной
фракции ДНК и могут быть метилированными. Множество тан-
демных повторов в сателлитной ДНК придает ей определенную
аномальность при центрифугировании (сходство поведения с ос-
новной ДНК в градиенте плотности хлористого цезия). В таких
случаях говорят о криптической (от лат. cryptus — скрытый)
сателлитной ДНК.
Установлено, что высокоповторяющиеся последовательности в
сателлитных ДНК обычно не транскрибируются. Следует отметить,
что сателлитные ДНК локализуются в области центромеры хромо-
сомы (выполняет структурную функцию). Предполагается, что
сателлитные ДНК произошли от мозаики последовательностей,
состоящих из 9 пн в трех повторах:
В начале 80-х годов в геноме человека были
A TGA обнаружены последовательности ДНК, обладаю-
GAAA А щ
И
е свойствами структурного полиморфизма —
Т ACT
это так
называемые гипервариабельные области
(ГВО),
обычно содержащие короткие, ГЦ-обога-
щенные и тандемно повторенные единицы. ГВО рекомендуются в
качестве маркеров-зондов при картировании генов. Варианты
кор-последовательности ГВО гена человеческого миоглобина были
названы минисателлитными.
В 1985 г. был предложен метод "генетической дактилоскопии
— ДНК" (от греч. dactilos — палец, scopein — смотреть) в целях
оценки эволюции человека по отцовской и материнской линиям
(А. Дж. Джеффрис, У. Уилсон, С. Л. Тейн). В последовательностях
ДНК отражаются происшедшие в прошлом мутации или, по Ф.
Крику, "замороженные события". Это, в частности, поможет кар-
тировать варианты последовательностей в локусе минисателлит-
ной ДНК. Эволюцию по женской линии удобно картировать по
163

митохондриальной ДНК, так как в сперматозоидах практически
нет митохондрий, но ими "начинены" яйцеклетки. Вот почему ДНК
клеточного ядра является сочетанием материнской и отцовской
ДНК, тогда как ДНК митохондрий передается только яйцеклеткой.
Отсюда становится понятным, почему в конце 70-х годов текущего
столетия возникла новая научная дисциплина молекулярная ант-
ропология.
В
ДНК различных организмов содержатся еще так называемые
палиндромы (от греч. palindrome — перевертыш) — последова-
тельности, повторяющиеся в обратном порядке:
5'-ЦТТЦГАТГГААГ-3'
3'-
ГААГЦТАЦЦТТЦ
- 5'
В суперспирализованном состоянии длинные палиндромы (10
и более пар оснований) образуют крестообразные структуры,
служащие сигналами для узнавания определенных участков ДНК
ферментами-метилазами, рестриктазами и регуляторными белка-
ми,
регулирующими действия генов.
В хромосомных ДНК прокариотических и эукариотических
клеток имеются также контролирующие или так называемые
"прыгающие" подвижные гены — транспозоны (Тп), впервые
открытые Б. Мак-Клинток в 1940 г. у кукурузы. Они находятся на
значительном расстоянии от других генов, на которые оказывают
влияние. Благодаря мутациям, названным "транспозонными взры-
вами", возможно массовое и в известной мере направленное
перемещение генетических элементов. Транспозоны способны
реплицироваться и внедряться (инсерция) в виде одной из копий
в новое место генома (ДНК
ядра).
У
бактерий преобладающая часть
транспозонов кодирует фермент транспозазу, катализирующую
реакцию встраивания транспозона в ДНК. В последнее время их
отождествляют с интронами, рассмотренными выше.
При сравнении последовательности нуклеотидов в хозяйской
ДНК до и после встраивания транспозона оказывается, что не-
сколько нуклеотидов этой ДНК после встраивания удваиваются —
дуплицируются. Эти дупликации ДНК окаймляют транспозон.
164

Причем, для каждого транспозона характерно определенное коли-
чество дуплицированных нуклеотидов. Принято считать, что при
встраивании транспозонов хозяйская ДНК перед этим фермента-
тивно расщепляется с образованием липких концов, к выступам
которых присоединяется транспозон, а оставшиеся бреши затем
заполняются нуклеотидными последовательностями, в результате
чего и образуются небольшие дупликации.
Следует подчеркнуть, что транспозонов много и они различны
(лишь перечень Tn-ов у дрозофилы составит целую книгу). Следо-
вательно необходима точная классификция их, и, возможно, в
недалеком будущем удастся ее создать.
К настоящему времени принято считать, что механизм транс-
позиции заключается в удвоении подвижных элементов и после-
дующем встраивании одной из копий транспозона в новое место
генома, а другая копия остается в прежнем месте. Вот почему
термин "транспозиция" неточен, поскольку транспозон не покида-
ет своего первоначального места, или сайта. Более правильно
рассматривать транспозицию процессом, в результате которого
возрастает число копий транспозона. Во благо сохранения струк-
туры генома (консерватизма его) транспозиции происходят очень
редко. Так, в среднем, частота их сравнима с частотой спонтанных
мутаций, то есть 10"
5
—10"
?
на поколение, а частота реверсии путем
делеций, или выпадений, отмечается еще реже (10~
6
—10"
10
).
Обращает на себя внимание тот факт, что Tn copia из мушки
дрозофилы ведет себя и как транспозон, и как ретровирус (к их
числу, кстати сказать, относится вирус иммунодефицита человека
— причина СПИД). В та-
Ц-
i
ких вирусах интеграция
ДНК осуществляется
способом, сходным с i
Е£ЧТ:
. :зийш&с
транспозицией. Поэтому
небеспочвенной стала
гипотеза о том, что виру-
сы — это транспозоны,
Приобретшие ДОПОЛНИ- ^
С
"
55
/ Встраивание транспозона в ДНК-мишень
г г
" (схема): 1 — транспозон, 2 — центральная часть,
тельные функции ИЛИ, з — концевые повторы, 4 — дупликация ДНК-ми-
напротив, транспозоны шени.
— это выродившиеся вирусы. Проблема остается открытой. Тем
не менее, обнаружены самые короткие транспозоны, кодирующие
165

Рис.
56. Ori-сайты в плазмиде pSClOl E.coli
(цифры по внешнему кругу 1—6 обозначают
места действия различных ферментов-ре-
стриктаз).
белки, которые участвуют
только в транспозиции. Сле-
довательно ДНК таких
транспозонов можно отне-
сти к разряду "эгоистиче-
ской", работающей лишь на
себя, то есть она функцио-
нирует во имя собственного
размножения.
Схематичное встраива-
ние транспозона (после ре-
пликации) в ДНК-мишень
изображено на рис. 55.
Репликация, или самоудвоение присуще ДНК и РНК, то есть в
таких случаях происходит перенос генетической информации
соответственно от ДНК к ДНК или, например у ряда вирусов, от
РНК к РНК. Репликация осуществляется полуконсервативным
способом, когда двухспиральная ДНК деспирализуется и каждая
нить индуцирует синтез комплементарной себе нити при участии
ДНК- или РНК-полимеразы.
Геномы бактерий и фагов реплицируются как единое целое, то
есть как организованные единицы репликации, или репликоны.
Каждый репликон содержит место (точку) инициации Ori (от англ.
origin — начало) — ориентированное направление репликации,
например OriC у Escherichia coli (рис. 56).
В
некоторых репликонах
содержится около 240—600 пн. В отдельных репликонах сущест-
вует два Ori, например, в так называемых челночных векторах,
способных реплицироваться в клетках прокариот и эукариот.
Инициация репликации осуществляется с помощью специфиче-
ских белков.
Репликация ДНК необходима функционирующей клетке для
восстановления (репарации), обмена генами между участками
хромосом (рекомбинация) и перемещения генов (транспозиции).
В течение однократного деления прокариотической или эука-
риотической клетки, независимо от числа хромосом в ней, весь
геном ее реплицируется также один
раз,
и только после завершения
репликации может произойти последующее деление. Удвоенный
геном подразделяется (сегрегирует) поровну в каждую дочернюю
клетку. Единицей сегрегации является хромосома, а единицей
репликации — репликон. Кроме точки Ori в репликоне имеется
166
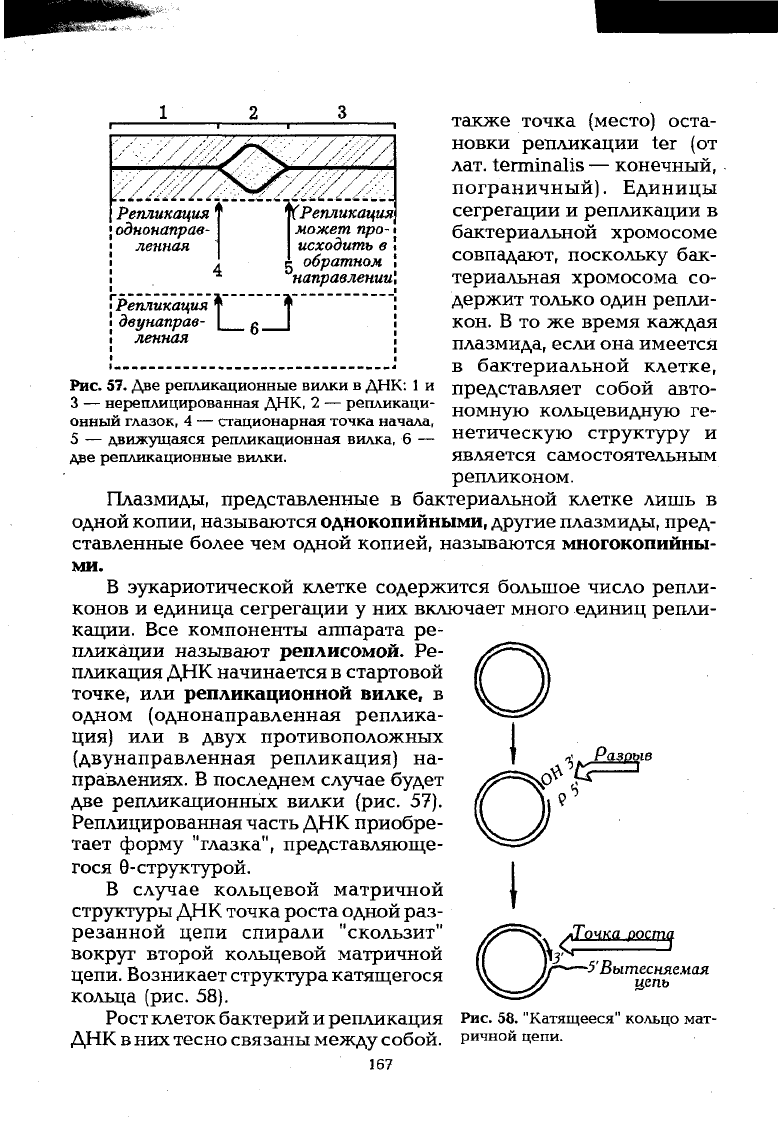
о
Репликация
однонаправ-
ленная
'(Репликация
может про-
исходить в
I обратном
направлении
Репликация т ^
двунаправ- \ g |
ленная
Рис.
57.
Две репликационные вилки в ДНК:
1
и
3 — нереплицированная ДНК, 2 — репликаци-
онный глазок, 4 — стационарная точка начала,
5 — движущаяся репликационная вилка, 6 —
две репликационные вилки.
также точка (место) оста-
новки репликации ter (от
лат. terminalis — конечный,
пограничный). Единицы
сегрегации и репликации в
бактериальной хромосоме
совпадают, поскольку бак-
териальная хромосома со-
держит только один репли-
кой. В то же время каждая
плазмида, если она имеется
в бактериальной клетке,
представляет собой авто-
номную кольцевидную ге-
нетическую структуру и
является самостоятельным
репликоном.
Плазмиды, представленные в бактериальной клетке лишь в
одной копии, называются однокопийными, другие плазмиды, пред-
ставленные более чем одной копией, называются многокопийны-
ми.
В эукариотической клетке содержится большое число репли-
конов и единица сегрегации у них включает много единиц репли-
кации. Все компоненты аппарата ре-
пликации называют реплисомой. Ре-
пликация ДНК начинается в стартовой
точке, или репликационной вилке, в
одном (однонаправленная реплика-
ция) или в двух противоположных
(двунаправленная репликация) на-
правлениях. В последнем случае будет
две репликационных вилки (рис. 57).
Реплицированная часть ДНК приобре-
тает форму "глазка", представляюще-
гося 0-структурой.
В случае кольцевой матричной
структуры ДНК точка роста одной раз-
резанной цепи спирали "скользит"
вокруг второй кольцевой матричной
цепи. Возникает структура катящегося
кольца (рис. 58).
Рост клеток бактерий и репликация
ДНК в них тесно связаны между собой.
167
Разрыв
\fi
Точка
роста
•5'Вытесняемая
цепь
Рис.
58. "Катящееся" кольцо мат-
ричной цепи.

Цикл клеточного деления на примере Е. coli можно представить
двумя временными интервалами, обозначаемыми латинскими бук-
вами С и D. Первая из них обозначает фиксированное время,
необходимое для репликации всей бактериальной хромосомы (на-
пример, 15 мин.), что соответствует скорости движения отдельной
Точка начала -. 35/0 гКонцевая точка
30
25
\
((
\
/
т_»>£гл°«и«м
т.»™
репликационнои вилки
с
присоединени-
*" "~\ , ем около 50000 пар оснований в
1
мин.;
D соответствует промежутку времени
(порядка 20 мин.) между завершением
репликации ДНК и делением клетки —
это как бы накопительный период. Сле-
довательно, весь так называемый корот-
кий клеточный цикл деления Е coli ук-
ладывается в среднем в 35 минут (рис.
59).
Длинный цикл клеточного деления
наблюдается тогда, когда велико время
до начала инициации. Для диплоидных
клеток эукариот характерно митотиче-
ское деление ядра с последующим обра-
зованием новых (дочерних) клеток. Этот
митотический цикл подразделют на 4 фазы: Gj, S, G2 и М. Фаза
Gi обозначает время роста клетки до синтеза ДНК (от англ. gap —
пробел); в фазу S происходит синтез ДНК (от англ. synthesis); G2
— вторая фаза роста в период.после синтеза ДНК; М — фаза
митоза (от англ. mytosis). На рис 60 представлена схема цикла
эукариотических клеток с обозначением продолжительности фаз
в культивируемых линиях дрожжевых клеток Schizosaccharomyces
pombe и клеток млекопитающих. На весь цикл деления клеток
эукариот усредненно приходится следующая доля времени: на
20 15
Рис.
59. Цикл деления Е. coli —
образование хромосомы со
многими вилками в процессе
репликации (а — инициация, б
— деление, в — терминация);
цифры обозначают минуты кле-
точного цикла.
1 г
Рис.
60. Схема цикла клеточного деления эукариотических клеток: 1 —
Schizosaccharomyces pombe, 2 — клетки млекопитающих.
фазу Gi — 30—40%, на фазу S
на фазу М — 5—10%.
30—50%,
на фазу G2 — 10—20%,
168

m-.
"-
За время движения репликационных вилок
в
клетках эукариот
присоединяется от 1000 до 3000 пн
в
1
минуту
—
у
млекопитающих
и
до 1000
пн/мин
— у
растений (здесь
не
исключается роль
пониженных температур для размножения клеток растений).
Синтез комплементарных цепей
во
время репликации двухни-
тевой ДНК
у
прокариот
и
эукариот катализируется ферментами,
называемыми ДНК-полимеразами. Их известно
3
(pol I,
pol
II
и pol
III
у
прокариот;
a, b и у - у
эукариот). Характеристика ДНК-по-
лимераз приведена
в
таблице
18.
Таблица 18. Характеристика прокариотических и эукариотических ДНК-
полимераз
ДНК-полиме-
раза
из клеток
Е.
1 coli:
I
II
III
из клеток
млекопита-
ющих:
а
Р
У
Размер,
кДа
109
120
250
110-120
45
60
Состав
одна цепь
—
гетеромуль-
тимер
несколько
субъединиц
1 субъединица
?
Ферментативная активность
а) элонгация
в
направлении
5'
—>
3' от 3' - ОН -
затравки
б)
3'
—»
5
1
-
экзонуклеаза
в)
5'
—»
3' -
экзонуклеаза
см.
для
I
б)
3'
—»
5' -
экзонуклеаза
а
- в)
см.
для I
репликативный синтез
ядерной
ДНК
(ядерная ДНК-
репликаза)
-
80%
репарация сегментов
поврежденной
ДНК
репликативный синтез
митохондриальной
ДНК -
2-15%
На раскрученной ДНК происходит непрерывный синтез веду-
щей цепи
в
направлении
5'
—>
3',
тогда
как на
комплементарной
цепи имеет место прерывистая репликация,
то
есть
в
направлении
169

5'
-» 3' на ней синтезируется серия фрагментов ДНК (фрагменты
Оказаки), которые затем соединяются, формируя отстающую цепь
(рис.
61). фрагменты Оказаки содержат примерно 1000—2000
'ччнч^тчч^т^
..5'
Т Ц А Г А Г
А Г У Ц У
Т Т А Ц Г
Г Ц
ОН
Рис.
61.
Фрагменты Оказаки, присоединенные к РНК — инициатору: а — матричная
ДНК, б — комплементарная ДНК, в — РНК—праймер, НТ—входящий нуклеозид-
трифосфат.
нуклеиновых оснований, и синтез фрагментов под каталитическим
действием ферментов РНК-полимераз, включая праймазу (ММ-60
кДа),
инициируется РНК—затравками, каждая из которых по длине
соответствует приблизительно 10 основаниям. РНК—затравка уд-
линяется под действием ДНК-полимеразы III, катализирующей
синтез цепи ДНК. РНК удаляется под действием ферментов экзо-
нуклеаз.
В так называемую затравочную реакцию вовлекается комплекс
белков —- праймосома. Например, праймосомный белок SSB свя-
зывается с одноцепочечной ДНК, стабилизируя ее; белок Dna В
образует предпраймерную РНК и участвует в передвижении прай-
мосомы; белок Dna С действует в комплексе с Dna В; белок Lig
сшивает разрывы между фрагментами Оказаки; белок п'-АТФ-аза,
белки п и п" образуют предзатравочную РНК, и др.
Предполагают, что праймосома собирается в каком-то одном
участке с последующим перемещением по одноцепочечной ДНК
к сайтам,тде инициируется синтез затравки. Направление движе-
ния праймосомы противоположно направлению синтеза ДНК от-
стающей цепи, но синхронно движению репликационной вилки.
По расчетным данным репликация ДНК у прокариот (вместе с
170

раскручиванием молекулы по 10 нуклеотидных пар одномоментно)
должна происходить со скоростью 400 000 оборотов в секунду, что
явно превышает реальную скорость репликации. Поэтому было
предположено, что должны существовать фиксаторы, включающи-
еся в молекулы ДНК у всех организмов. Такие фиксаторы были
обнаружены. К ним относятся ферменты — топоизомеразы. От-
дельные из них катализируют реакции объединения молекул ДНК
в зацепленные кольца — катенаны, топоизомераза II, или гираза,
катализирует суперспирализацию ДНК, топоизомераза I способна
разрезать одну из цепей суперспирализованной ДНК, при этом
цепи раскручиваются и число супервитков уменьшается, после
чего этот же фермент устраняет разрыв в нити ДНК.
Двигателем в распределении реплицированных молекул ДНК
по дочерним клеткам (по крайней мере - у прокариот) выступает
клеточная мембрана, к которой прикрепляется ДНК.
Таким образом, весь катализируемый ферментами процесс
репликации ДНК можно подразделить на 3 этапа: инициацию,
элонгацию (рост цепи) и терминацию. В процессе инициации
происходит разделение нитей ДНК с образованием репликацион-
ной вилки, формирование праймосомы и синтез затравочной РНК.
Этап роста цепи, или элонгации, реализуется в синтезе ДНК с
помощью ДНК-полимераз. Терминация, или окончание синтеза
ДНК, происходит благодаря выключению реакции с помощью
специфического "стоп-сигнала" от специального кодона (термина-
тора) в матричной цепи.
Такие же 3 этапа выделяют в процессах транскрипции и
трансляции ДНК.
Транскрипция — это процесс переписывания закодированной
в ДНК информации и перенос ее к месту синтеза белка (на
рибосомы). Этап инициации при транскрипции заключается во
взаимодействии РНК-полимеразы с ДНК-матрицей; элонгация - в
ферментативном синтезе мРНК на матрице ДНК; терминация —
в остановке синтеза мРНК благодаря "стоп-сигналу" от гена —
терминатора.
Трансляция заключается в переводе закодированной в мРНК
информации в полипептидную цепь. Организующими центрами
процесса трансляции являются рибосомы. При трансляции на
этапе инициации происходит активация аминокислот с помощью
ферментов аминоацил-тРНК-синтетаз (АРСаз) при использовании
энергии АТФ с последующим образованием комплекса
инициации, включающего 3 фактора инициации
(IF-1,
IF-2,
IF-3 — у прокариот, eIF-2,
eIF-3,
eIF-5 и др. — у эукариот), мРНК,
171

гуанозилтрифосфат (ГТФ) и 30S(40S) — субчастицу рибосомы.
Указанный комплекс соединяется с 50S(60S) — субчастицей рибо-
сомы и формирует функциональную 70S (80S) рибосому. На этапе
элонгации в ходе трансляции 'осуществляется синтез полипептид-
ной цепи функционирующей рибосомой с участием факторов
элонгации (EF-Tu, EF-Ts, EF-G — у прокариот; EF-1 и EF-2 — у.
эукариот). Указанные факторы не входят в структуру рибосом, а
присоединяются к ним на определенных этапах.
Во время синтеза белка рибосомы движутся вдоль мРНК,
последовательно считывая триплеты и поэтапно наращивая по-
липептидную цепь. У мРНК, как правило, имеется постоянная
рамка считывания — кодон AUG. При элонгации одна мРНК
связывается с несколькими рибосомами, образуя функционирую-
щий комплекс - полирибосомы, или полисомы. Из указанных трех
этапов с наибольшей скоростью протекает элонгация. Терминация
процесса трансляции осуществляется стоп-кодоном в мРНК.
Как следует из представленных выше данных РНК занимает
место посредника между ДНК и белком. Причем эта центральная
функция присуща мРНК, тогда как тРНК и рРНК являются транс-
криптами, обладающими активностью как завершенные в постро-
ении и функции молекулы. Исключение представляют некоторые
РНК — содержащие вирусные геномы, по матрице которых син-
тезируется РНК, причем реализация генетической информации
может осуществляться по схемам: для одних вирусов, у которых
отсутствует транскрипция, РНК —> Белок (вирус полиомиелита
и др.), для других, располагающих собственной вирионной РНК —
зависимой РНК-полимеразой, или вирионной транскриптазой,
РНК
—>
РНК
—>
Белок (вирусы гриппа, кори и др.), и, наконец, для
третьих — РНК -> ДНК -» РНК
—>
Белок (ретровирусы, в том числе
— ВИЧ, или вирус СПИД).
мРНК, рРНК и тРНК у бактерий синтезируются под каталити-
ческим действием одной и той же РНК-полимеразы (ММ-480 кДа).
В клетках эукариот обнаружены три ядерные РНК-полимеразы (I,
II и III), а также РНК-полимеразы митохондрий и хлоропластов.
Установлено, что РНК-полимераза I, находящаяся в ядрышке,
отвечает за синтез про-рРНК, из которой впоследствии образуются
28S и 18S рРНК; РНК-полимераза II катализирует реакцию синтеза
про-мРНК и только с этим ферментом связано так называемое
кэпирование РНК. А. Шаткин в 1976 г. впервые описал характер-
ную группировку, присоединяющуюся к 5'-концу фактически всех
про-мРНК и мРНК посттранскрипционно. Автор назвал ее КЭПом
172