Хаусман К., Хюльсман H., Радек Р. Протистология
Подождите немного. Документ загружается.


338 Избранные главы общей протистологии
Кариоплазма содержит хроматин,
который имеет тонкофибриллярную
структуру и один или несколько элект-
ронно-плотных агрегатов — ядрышек
(рис. 296). Ядрышки являются центра-
ми синтеза рибосомных субъединиц.
Они отсутствуют в микронуклеусах
инфузорий и в ядрах некоторых гамет.
Степень конденсации хроматина в
интерфазе различна у разных протис-
тов. Компактный, плотно упакованный
хроматин, заполняющий все ядро, ха-
рактерен для ядер микрогамет боль-
шинства Apicomplexa и микронуклеусов
цилиат. У эвгленовых хроматин интер-
фазного ядра формирует плотные па-
лочковидные скопления, похожие на
митотические хромосомы. У ряда ги-
пермастигид (например, Barbulanym-
pha, Trichonympha) хромосомы видны
постоянно, они прикреплены кинетохо-
рами к внутренней стороне ядерной
оболочки. У большинства динофлагел-
лят ядра мелкие и в них постоянно при-
сутствуют конденсированные хромосо-
мы, которые принципиально отличают-
ся от хромосом всех других протистов
способом упаковки хроматина. Предпо-
лагается, что каждая хромосома диноф-
лагеллат состоит из длинной кольцевой
молекулы ДНК, уложенной в суперспи-
раль. По этому признаку ядро динофла-
геллат разительно отличается от всех
остальных эукариотических ядер, и ему
дано собственное название — динока-
рион. Уникальные черты организации
динокариона считаются вторично при-
обретенными.
Как и у многоклеточных, ядерная
ДНК протистов обычно ассоциирована
с гистонами и, таким образом, органи-
зована в хроматин. Все пять гистонов,
известные для многоклеточных, обна-
ружены в макронуклеусе Tetrahymena.
Их аминокислотные последовательнос-
ти на определенных участках отличны
от таковых многоклеточных. Интерес-
но, что транскрипционно пассивное
ядро — микронуклеус — по составу
гистонов существенно отличается от
макронуклеуса. Ядра большинства ди-
нофлагеллат лишены гистонов, но все-
таки содержат немного основных бел-
ков.
Жизненные циклы протистов мно-
гообразны: гаплофазный цикл с зиготи-
ческой редукцией (характерен для Vol-
vocida, динофлагеллат, гипермастигид и
оксимонад, Apicomplexa), дишюфазный
с гаметической редукцией (например,
Actinophrys, Noctiluca) и гетерофазный
с чередованием диплоидной и гаплоид-
ной фаз (фораминиферы). Поэтому у
них широко представлены гаплоидные
и диплоидные ядра, а также немало и
полиплоидных, которые отличаются
большими размерами.
Микронуклеусы всех изученных
цилиат диплоидны
1
, а плоидность мак-
ронуклеуса обычно высока (за исклю-
чением кариореликтид с квазидиплоид-
ным макронуклеусом — см. гл. 2). Со-
держание ДНК в макронуклеусе Tetra-
hymena thermophila в 10-20 раз больше,
чем в микронуклеусе. Макронуклеус
гипотрих содержит только 5% всего ге-
1
По-видимому, существуют редкие исключения. Есть данные, что у определенных линий
Paramecium caudatum и Paramecium aurelia микронуклеус низкополиплоиден (тетраплоиден, ок-
таплоиден) в норме. — Прим. ред.

Ядро и половое размножение 339
нома микронуклеуса. Большее количе-
ство ДНК в макронуклеусе объясняет-
ся тем, что немногие оставшиеся гены
представлены огромным количеством
копий. ДНК макронуклеуса гипотрих
представлена миллионами коротких
линейных фрагментов размером в ген.
Число копий каждого фрагмента прямо
пропорционально размерам клетки.
Роль микронуклеуса
и макронуклеуса
в жизненном цикле
цилиат
Экспериментальные исследования
привели к широко распространенному
мнению, что микронуклеус не участву-
ет в поддержании и регуляции клеточ-
ных функций, а отвечает только за
хранение и рекомбинацию генетичес-
кого материала. Это подтверждается
существованием видов без микронук-
леусов. Однако эксперименты по пере-
саживанию ядер, особенно на Parame-
cium и гипотрихах, показывают, что
микронуклеусы все-таки выполняют и
другие функции. У Paramecium с экс-
периментально удаленными микронук-
леусами в следующем поколении осо-
бей развивается ненормальный рото-
вой аппарат. Поэтому можно предпо-
ложить, что микронуклеус участвует в
формировании ротового аппарата (т.е.
в стоматогенезе). После нескольких
циклов бесполого размножения рото-
вой аппарат формируется уже без на-
рушений, то есть макронуклеус бе-
рет на себя эту функцию утраченного
микронуклеуса.
Преобразования ядра
в ходе жизненного
цикла
Клеточный цикл эукариотной клет-
ки традиционно делится на несколько
четких стадий (см. рис. 321). В интер-
фазе, которая включает стадию репли-
кации ДНК (S-фазу) и две промежуточ-
ные фазы G, и G
2
, клетка непрерывно
растет. В М-фазе происходит распреде-
ление хромосом по дочерним ядрам
(митоз) и деление клетки (цитокинез).
Репликация генетического
материала
Синтез ДНК может идти на разных
стадиях жизненного цикла. У много-
ядерных гомокариотных протистов реп-
ликация генетического материала и
митоз идут независимо в каждом ядре
и не связаны непосредственно с циклом
деления клетки. У цилиат с нескольки-
ми микронуклеусами последние также
вступают в S-фазу независимо друг от
друга и от макронуклеуса.
В макронуклеусах гипотрих синтез
ДНК и гистонов идет синхронно в од-
ной или двух особых зонах (рис. 299).
Преобразование хроматина, отчетливо
видное на микрофотографиях, начина-
ется в определенных зонах вблизи ядер-
ной оболочки. Затем эти зоны сливают-
ся в так называемую репликационную
полоску, которая движется через весь
макронуклеус, вызывая по ходу удвое-
ние ДНК и гистонов. Непосредственно
перед началом деления ядра репликаци-
онная полоска исчезает. В макронуклеу-
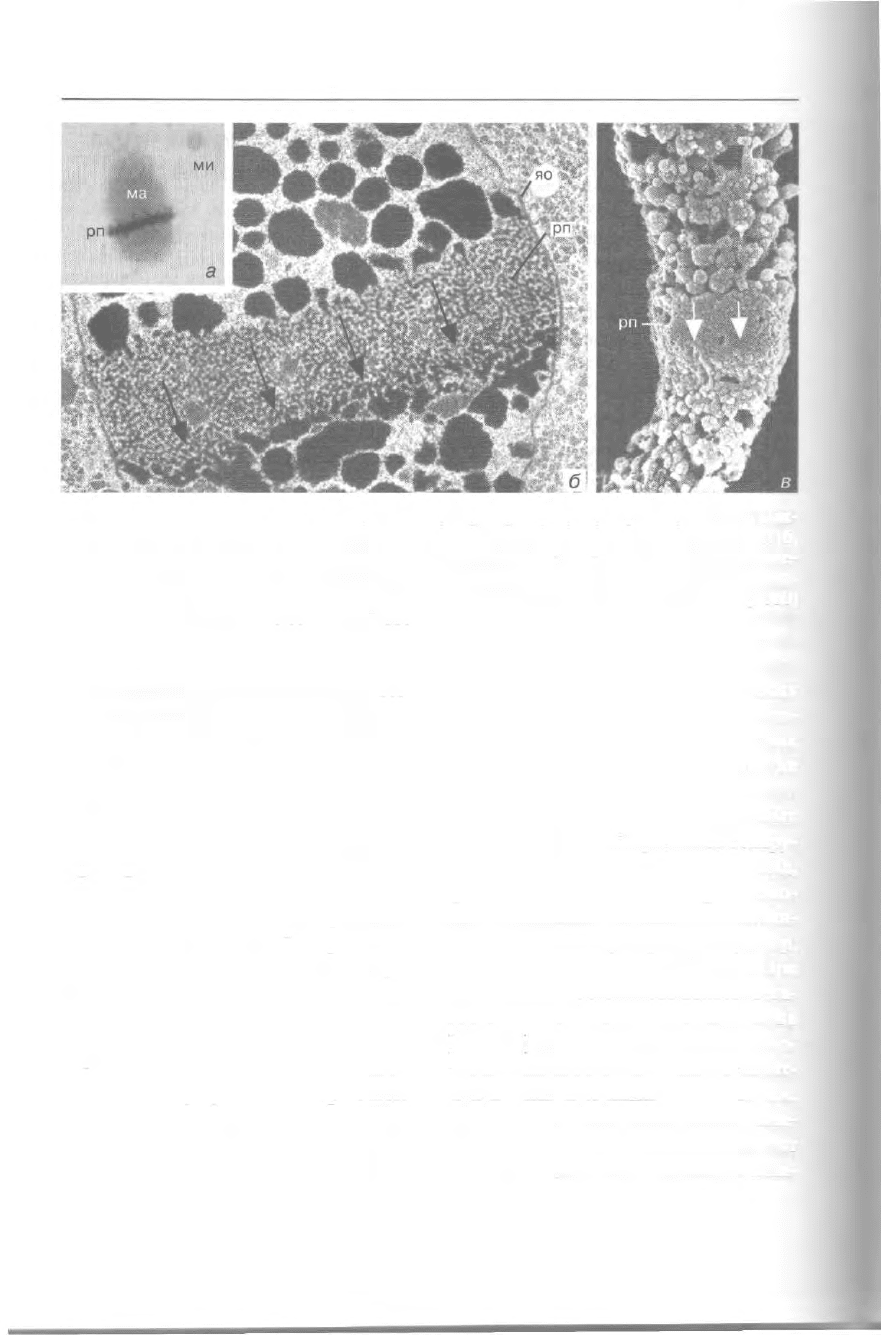
340 Избранные главы общей протистологии
Рис. 299. Репликационная полоска (рп) в макронуклеусе (ма) Stylonychia (световая мик-
роскопия) (а) и Euplotes (трансмиссионная и сканирующая электронная микроскопия) (б,
в), ми — микронуклеус, яо — ядерная оболочка. Стрелки указывают направление движе-
ния репликационной полоски (а — из: Steinbruck: Europ. J. Protistol. 26 [1991] 2; б — лю-
безно предоставлен К. Эйслер, Тюбинген; в — из: Arikawa et al.: Europ. J. Protistol. 36 [2000]
40). Увел.: a - 650x, 6- 13 500x, в — 7 000x.
сах других цилиат синтез ДНК на уров-
не ультраструктуры не выявляется.
Митоз
В противоположность высшим
эукариотам, у которых форма деления
ядра варьирует лишь незначительно, у
протистов обнаружены существенно
различающиеся типы митоза (рис. 300).
Они различаются судьбой ядерной обо-
лочки, а также положением, симметри-
ей и строением веретена деления и цен-
тров его организации.
При открытом митозе (как у много-
клеточных организмов) ядерная оболоч-
ка полностью разрушается, а при закры-
том митозе, наоборот, оболочка ядра
сохраняется на всех его стадиях. При
полузакрытом митозе ядерная оболочка
разрушается только на определенных
участках. В ортомитозе аппарат верете-
на имеет осевую симметрию, тогда как
при плевромитозе оси полуверетен не
совпадают.
Характерный для многоклеточных
животных открытый ортомитоз обнару-
жен у многих протистов: хризомонад,
криптомонад, гаптомонад, празиномо-
над, пединеллид, акантамеб, лабирин-
тул, некоторых грегарин (например,
Monocystis) и некоторых Ешпуха. При
полузакрытом ортомитозе центры орга-
низации микротрубочек веретена
(ЦОМТы) развиваются на одном полю-
се снаружи ядра и в профазе расходят-
ся к противоположным его полюсам.
После этого ядерная оболочка прорыва-
ется на полюсах и микротрубочки от
ЦОМТов входят в ядро, формируя аппа-
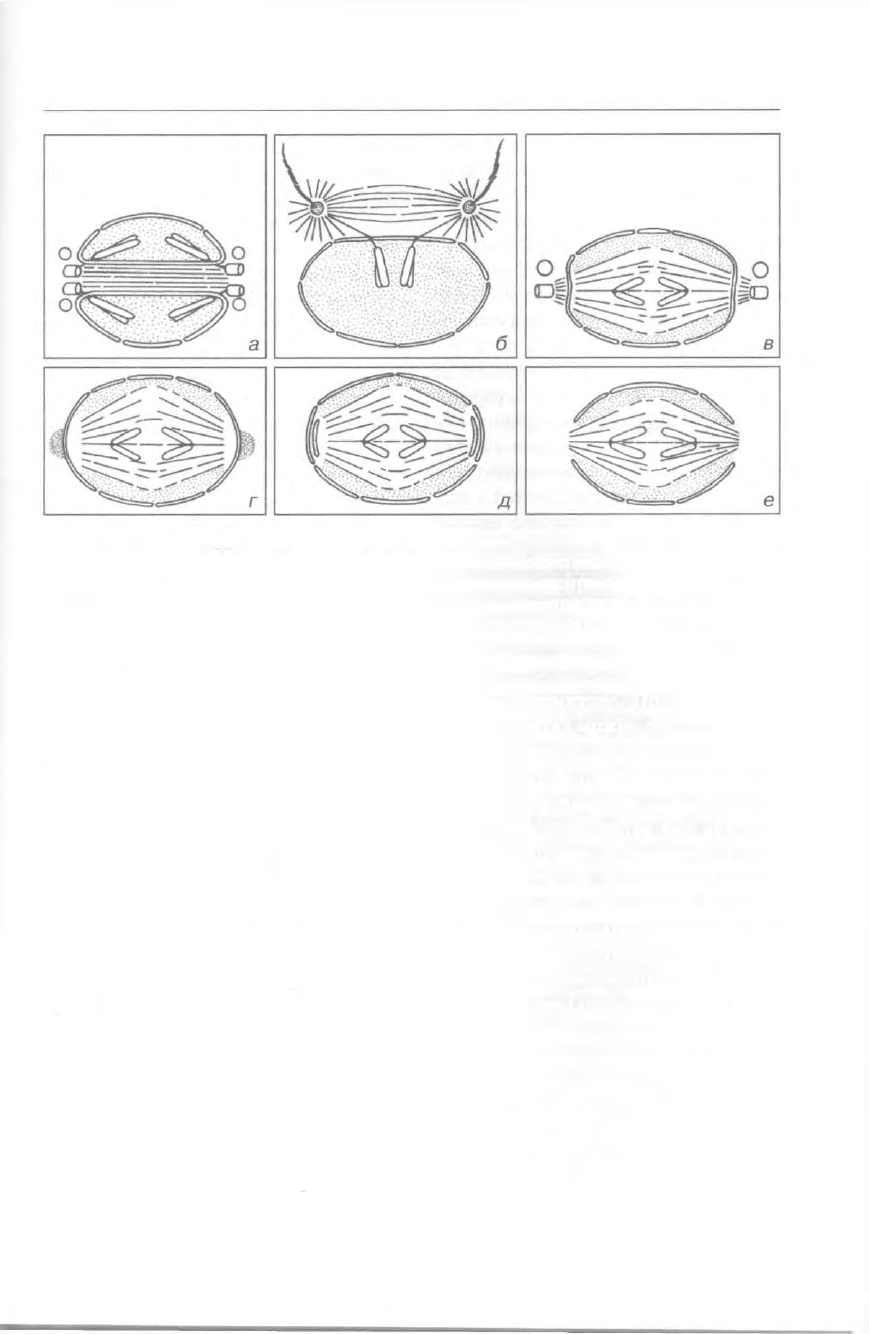
Ядро и половое размножение 341
Рис. 300. Типы митозов в различных таксонах (анафаза): а — микротрубочки проходят в
каналах ядра (динофлагеллаты); б — микротрубочки внеядерного веретена взаимодейству-
ет с хромосомами (гипермастигиды); в, г — внутриядерное веретено и внеядерные орга-
низующие центры (фораминиферы); д — аппарат веретена внутри интактной ядерной обо-
лочки (инфузории); е — ядерная оболочка разрушена на полюсах ядра (некоторые хризо-
фитовые) (по Грелю).
рат веретена. Такой тип митоза описан
для хлоромонад (особенно Volvocida) и
рафидомонад, некоторых крупных амеб
(Amoeba proteus), хоанофлагеллат
(Monosiga ovata), грегарин (например,
Deplauxis) и некоторых солнечников
(например, Actinophrys). При закрытом
ортомитозе аксиально-симметричное
веретено развивается внутри ядерной
оболочки. Этот тип митоза обнаружен
у трипаносоматид, раковинных амеб
{Euglypha, Arcella), вампиреллид (на-
пример, Arachnula, Lateromyxa, Vampy-
rella) и в микронуклеусах инфузорий. У
трипаносоматид формируются два неза-
висимых полуверетена. При этом у од-
них видов (например, Trypanosoma
cruzi) они располагаются вдоль одной
оси (ортомитоз), а у других (например,
Т. raiae) лежат под углом друг к другу,
как в плевромитозе. Поэтому митоз
трипаносоматид можно считать проме-
жуточным между закрытым ортомито-
зом и закрытым плевромитозом.
При полузакрытом плевромитозе
ЦОМТы располагаются вне ядерной
оболочки, которая перфорируется, ког-
да микротрубочки полуверетен прохо-
дят в ядро. Этот тип митоза типичен
для Apicomplexa. На стадии множе-
ственного деления (шизогонии) после
очередного митоза веретено сохраняет-
ся, а хромосомы остаются прикреплен-
ными к его микротрубочкам до тех пор,
пока не закончатся все другие митозы.
Такой механизм препятствует случайно-
му распределению хромосом во вре-
менно полиплоидном ядре. Закрытые
плевромитозы идут у фораминифер,
радиолярий, оксимонад, некоторых пра-

342 Избранные главы общей протистологии
зиномонад, кинетопластид, микроспо-
ридий, гаплоспоридий и многих грибов.
ЦОМТы микротрубочек веретена закла-
дываются в ядерной оболочке поблизо-
сти друг от друга и затем расходятся в
противоположные стороны за счет раз-
растания между ними ядерной оболоч-
ки или каким-то образом перемещают-
ся в ней.
У трихомонад, гипермастигид и ди-
нофлагеллат организующие центры ве-
ретена расположены снаружи ядер-
ной оболочки; веретено также развива-
ется вне ядра, либо сбоку от него (три-
хомонады, гипермастигиды), либо
внутри пронизывающих его каналов
(динофлагеллаты).
На рис. 300а показан динокарион с
пронизывающим его каналом. Стенка
канала сформирована обычной ядерной
оболочкой. Микротрубочки берут нача-
ло от центросом, расположенных снару-
жи от ядра. И полюсные, и кинетохор-
ные микротрубочки проходят по кана-
лу. Последние присоединяются к ядер-
ной оболочке. В тех же точках к
ядерной оболочке изнутри прикрепля-
ются хромосомы. Рис. 3006 иллюстри-
рует сходную картину. Микротрубочки
отходят от центросом, которые у пара-
базалий называются аттрактосферами,
и присоединяются снаружи к ядерной
оболочке, однако здесь нет пронизыва-
ющих ядро каналов.
Новая оболочка дочерних ядер со-
бирается, как правило, из частей или
фрагментов старой, однако в ряде слу-
чаев (например, Actinophrys, микронук-
леус некоторых инфузорий) она появля-
ется под оболочкой материнского ядра
еще до того, как последняя дегради-
рует.
Большое разнообразие типов деле-
ния ядра позволяет предположить, что
их становление произошло в процессе
эволюции протистов. Предковые черты
могут сохраняться в современных так-
сонах наряду с примитивными и про-
двинутыми признаками. При делении
прокариот хромосомы прикрепляются к
плазматической мембране и разделяют-
ся благодаря росту мембраны между
точками прикрепления. Внешне это на-
поминает митоз динофлагеллат (дино-
митоз), поэтому ряд авторов считает
диномитоз примитивным. Однако моле-
кулярно-биологические исследования
показывают, что динофлагеллаты пред-
ставляют собой эволюционно продви-
нутую группу. Диномитоз существенно
отличается от деления прокариот по
многих параметрам, в частности, учас-
тием микротрубочек. Эволюционная
последовательность разных типов ми-
тоза у протистов еще неизвестна
1
.
Деление макронуклеуса
инфузорий
Макронуклеус инфузорий (исклю-
чая кариореликтид) делится во время
цитокинеза. Обычно он мигрирует в
область разделения клеток еще до фор-
мирования борозды деления, затем вы-
тягивается продольно, располагаясь в
обеих будущих дочерних клетках, и пе-
решнуровывается в процессе цитокине-
за. Его половинки разделяются непос-
1
Пути эволюции митоза аргументированно обсуждаются в книге И.Б. Райкова «Ядро простей-
ших». Л.: Наука. 1978. — Прим. пер.

Ядро и половое размножение 343
редственно перед отделением дочерних
клеток друг от друга. В течение всего
этого процесса оболочка ядра сохраня-
ется.
В процессах миграции, удлинения и
формирования новых ядер участвуют
внутриядерные и цитоплазматические
микрофиламенты и микротрубочки.
Эксперименты на Stentor показали, что
расположение макронуклеуса и его уд-
линение контролируется кортикальны-
ми элементами. По-видимому, это спра-
ведливо и для других цилиат (напри-
мер, Bursaria, Paramecium), макронук-
леусы которых прикрепляются к
кортексу клетки.
Деление макронуклеуса инфузорий
часто называют амитозом, поскольку
отсутствуют хромосомы и типичный
аппарат веретена. С другой стороны,
маловероятно, что генетический мате-
риал между дочерними клетками рас-
пределяется случайно. Разветвленные
или сегментированные ядра обычно
конденсируются перед делением в еди-
ную компактную массу; то же происхо-
дит и с набором мелких макронуклеу-
сов, как, например, у Urostyla grandis.
Более того, генетический материал
иногда перемешивается внутри ядра.
Так, у Homalozoon было замечено вра-
щение содержимого макронуклеуса пе-
ред делением клетки. Внутриядерные
микротрубочки в макронуклеусах Рго-
tocruzia и других инфузорий, по-види-
мому, участвуют в разделении элемен-
тов, соответствующих хромосомам.
Возникающие в составе ДНК различия
между дочерними ядрами нивелируют-
ся либо избирательным синтезом, либо
избирательным удалением генетическо-
го материала.
Мейоз
Мейоз большинства эукариот состо-
ит из двух последовательных делений
ядра, которые сопровождаются клеточ-
ными делениями. Если хромосомы в
интерфазе деспирализованы, они кон-
денсируются во время пролонгирован-
ной профазы I первого мейотического
деления. Предварительно удвоенные
гомологи конъюгируют, образуя бива-
ленты, связанные синаптонемальными
комплексами (рис. 301). Реципрокный
обмен гомологичными участками ДНК
между несестринскими хроматидами
(генетическая рекомбинация) осуществ-
ляется в процессе кроссинговера. В
метафазе I биваленты выстраиваются
по экватору веретена, затем гомологич-
ные хромосомы разделяются и движут-
ся к противоположным полюсам клет-
ки (анафаза I). Материнские и отцовс-
кие гомологи случайно распределяются
между дочерними ядрами, а количество
хромосом уменьшается вдвое. Второе
мейотическое деление соответствует
митозу. В результате мейотических де-
лений из каждого диплоидного ядра
получается четыре генетически различ-
ных гаплоидных ядра.
У большинства протистов мейоз
соответствует этой общей схеме. Одна-
ко у многих из них особенности мейо-
за на ультраструктурном уровне еще
неизвестны. Синаптонемальные комп-
лексы, свидетельствующие о том, что
мейоз является двухступенчатым, на-
блюдались во многих таксонах: у аме-
боидных протистов (Arcella, Lateromyxa
gallica), микроспоридий (Amblyospora),
Myxozoa (Aurantiactinomyxon), дино-
флагеллат (Prorocentrum), инфузорий,
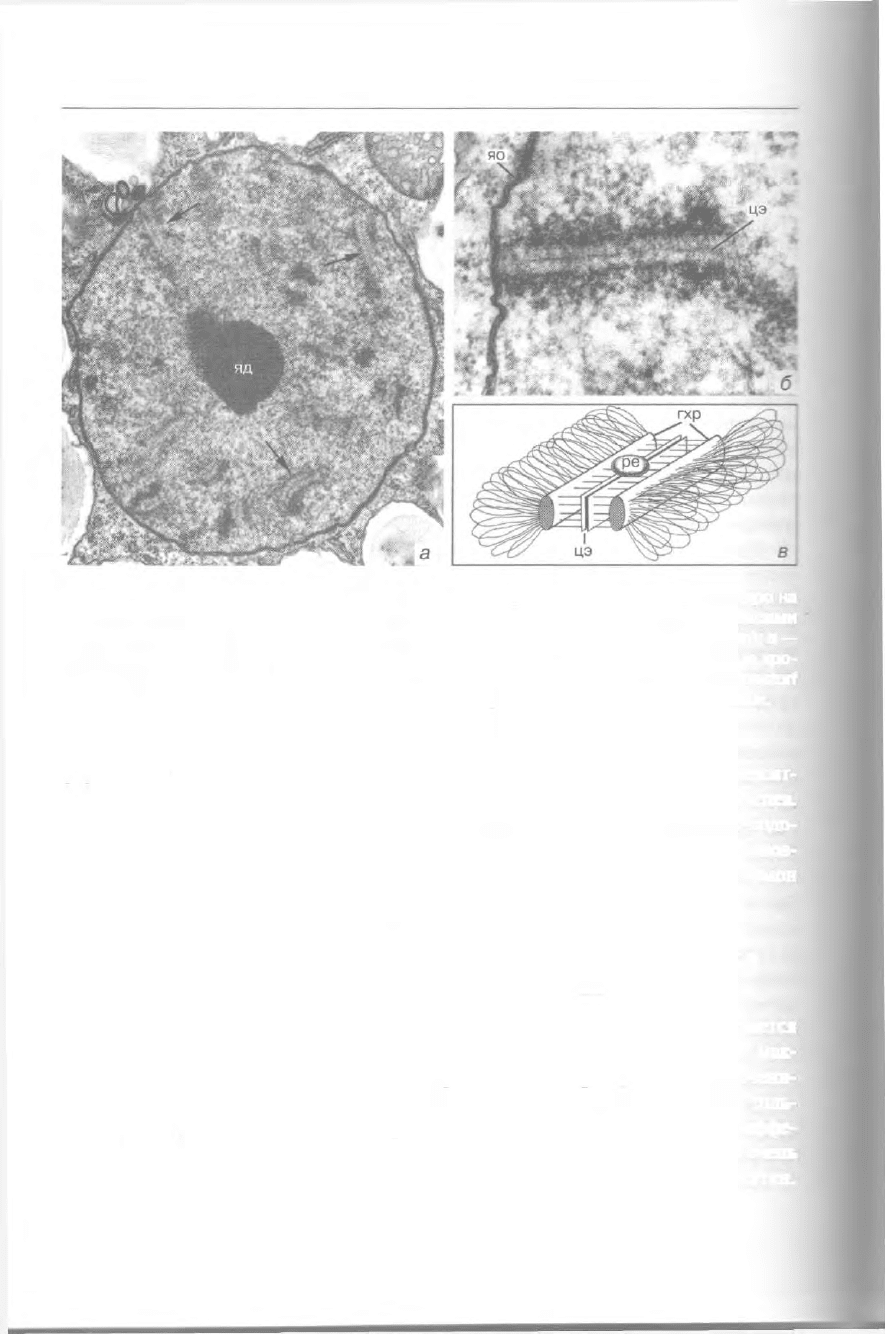
344 Избранные главы общей протистологии
Рис. 301. Синаптонемальные комплексы у Lateromyxa gallica (Vampyrellidae): а — ядро на
стадии зиготены с ядрышком (яд) и несколькими синаптонемальными комплексами
(стрелки); б — прикрепление синаптонемального комплекса к ядерной оболочке (яо); в —
схема спаривания гомологов в синаптонемальном комплексе, гхр — гомологичные хро-
мосомы, ре — рекомбинационный узелок, цэ — центральный элемент (а и б — из: Ropstorf
et al.: Europ. J. Protistol. 29 [1993] 302; s — по Крстичу). Увел.: a - 32 000x, 6-75 OOOx.
фораминифер, Chlamydomonas, Labyrin-
thula и Actinophrys. Считается, что у
споровиков (Coccidea, Haemosporida и
Piroplasmida) мейоз также двухступен-
чатый, хотя иногда он идет без кроссин-
говера.
Так называемый одноступенчатый
мейоз (с одним делением ядра) был
описан у жгутиконосцев из кишечника
таракана Cryptocercus. При этом типе
мейоза гомологичные хромосомы конъ-
югируют, по-видимому, без предвари-
тельного удвоения, между ними не на-
блюдаются хиазмы и кроссинговер. Та-
ким образом, уровень рекомбинации при
одноступенчатом мейозе значительно
ниже, чем при двухступенчатом. Мейоз
с одним делением ядра, по всей вероят-
ности, имеется только у жгутиконосцев,
обитающих в Cryptocercus. Есть подо-
зрение, что определенную роль в станов-
лении этого типа мейоза сыграл гормон
линьки таракана (эгдизон).
Развитие макронуклеуса
у инфузорий
Геном макронуклеуса развивается
из генома микронуклеуса. Новые мак-
ронуклеусы инфузорий (за исключени-
ем кариореликтид) формируются толь-
ко после конъюгации. Процесс диффе-
ренциации генома макронуклеуса очень
сложен и еще не до конца понятен.

Ядро и половое размножение 345
Содержание ДНК (пг)
а б в г д е ж з
Рис. 302. Развитие зачатка макронуклеуса у Stylonychia lemnae: а — сразу после разделе-
ния эксконъюгантов (t = 0) размер и содержание ДНК зачатка макронуклеуса соответствует
таковым микронуклеуса; б — увеличение содержания ДНК; в — формирование гигантских
хромосом; г — фрагментация гигантских хромосом и падение содержания ДНК; д — фор-
мирование ядрышек (яд); е, ж — появление репликационных полосок (рп) и рост содер-
жания ДНК; з — через 100 часов макронуклеус полностью сформирован (по Аммерману).
У большинства изученных к настояще-
му времени цилиат в процессе реорга-
низации ДНК выделяют по меньшей
мере три события: разрыв хромосом,
удаление внутренних участков
1
и амп-
лификация ДНК. У Paramecium фраг-
ментация хромосом микронуклеуса
приводит к образованию субхромосом-
ных участков размером 300-600 т.п.н. У
гипотрих размер фрагментов генома в
макронуклеусе варьирует от 0,4 до 20
т.п.н.
Эти молекулы размером в один ген
похожи на настоящие хромосомы в том
отношении, что они способны к авто-
номной репликации и транскрипции,
однако у них нет центромер для при-
крепления микротрубочек. Молекулы
ДНК размером в ген обнаружены так-
же в макронуклеусе олиготрих (напри-
мер, Halteria) и филлофарингид (напри-
мер, Chilodonella). Окончательная сте-
пень амплификации ДНК варьирует от
вида к виду, но постоянна в пределах
вида. Так, у Paramecium tetraurelia уро-
вень плоидности макронуклеуса 1000—
2000, а у Tetrahymena pyriformis —
только 45. У всех инфузорий гены ри-
' Внутренних элиминируемых последовательностей — internal eliminated segments, или internal
eliminated sequences. — Прим. пер.

346 Избранные главы общей протистологии
босомных РНК в макронуклеусе — это
отдельные короткие молекулы, пред-
ставленные большим количеством ко-
пий.
Экстенсивная репликация ДНК в
макронуклеарном зачатке приводит к
развитию гигантских хромосом, напо-
минающих политенные хромосомы на-
секомых. Это отмечено у большинства
стиготрих (например, Stylonychia) и ги-
потрих (например, Euplotes), а также у
некоторых представителей других
групп (например, Chilodonella cuculus,
Loxophyllum meleagris, Nyctotherus
cordiformis). У Stylonychia политениза-
цию претерпевают лишь немногие из
хромосом микронуклеуса, тогда как
другие деградируют. Политенные хро-
мосомы распадаются на фрагменты,
(рис. 302). Во время развития макро-
нуклеуса разрушается большая часть
ДНК, в том числе повторяющиеся пос-
ледовательности и специфичные мик-
ронуклеарные гены. В результате в за-
чатке макронуклеуса Stylonychia lemnae
остается только около 2% всех генов
микронуклеуса. Они представлены от-
дельными мелкими фрагментами ДНК
(один фрагмент — один ген), рассеян-
ными в макронуклеусе.
Такие инфузории, как Paramecium,
Tetrahymena, гетеротрихи и Litostoma-
tea, не формируют политенных хромо-
сом. Однако и у них наблюдается вре-
менное падение содержания ДНК при
развитии макронуклеуса. Например, у
Tetrahymena во многих участках гено-
ма элиминируется часть нуклеотидов.
Реорганизация ДНК у Paramecium, по-
видимому, не сводится к простому
уменьшению размеров хромосом, так
как из уникального набора хромосом
микронуклеуса появляются различные,
но близкие друг другу по нуклеотидным
последовательностям субхромосомы
макронуклеуса. Показано, что фрагмен-
тация хромосом у Paramecium (как и у
других цилиат) связана с вырезанием
участков ДНК, однако размеры удаляе-
мых участков, похоже, различаются в
разных копиях субхромосом. Так же как
и у гипотрих, степень редукции генома
при развитии макронуклеуса здесь пря-
мо пропорциональна размеру генома
микронуклеуса: чем меньше геном мик-
ронуклеуса, тем меньше степень его
редукции при созревании макронукле-
уса. Например, у Paramecium aurelia
микронуклеарный геном 270-320 млн
п.н. редуцирован до 46-80 млн п.н. в
макронуклеусе; у P. bursaria геном мик-
ронуклеуса 3500 млн п.н. в макронук-
леусе редуцирован до 17 млн п.н.
Половой процесс
в жизненном цикле
протистов
Половой процесс заключается, глав-
ным образом, в слиянии двух гаплоид-
ных гаметических ядер, образовавших-
ся в результате мейоза, и формировании
диплоидной зиготы. Он не обнаружен у
эвгленовых, хоанофлагеллат, трихомо-
над, шизопиренид и некоторых других
протистов. Неизвестно, первично отсут-
ствует у них половой процесс, или ут-
рачен в эволюции, или просто половые
стадии еще не найдены.
У многих протистов гаметические
ядра формируются в специальных
дифференцированных половых клет-
ках, или гаметах. Если половая диф-
ференцировка проявляется уже в роди-

Ядро и половое размножение 347
тельских клетках, такие клетки назы-
ваются гамонтами. Гаметы могут быть
морфологически одинаковы (изогаме-
ты) или различны (анизогаметы). Оп-
лодотворение неподвижной яйцеклет-
ки подвижной мужской гаметой назы-
вается оогамией. Следует различать
слияние гамет (гаметогамию) и слия-
ние гамонтов (гамонтогамию). Слия-
ние гаметических ядер, сформирован-
ных одной родительской клеткой, на-
зывается автогамией.
В жизненном цикле многих проти-
стов происходит чередование вегетатив-
ного и полового размножения. При го-
мофазном чередовании поколений тип
размножения меняется, а число хромо-
сом остается неизменным. Это харак-
терно, например, для Apicomplexa (см.
рис. 84). Смена поколений у многих
Apicomplexa связана со сменой хозяев.
Для фораминифер характерно гетеро-
фазное чередование поколений, по-
скольку мейоз происходит в середине
жизненного цикла.
У некоторых колониальных проти-
стов вегетативные и генеративные клет-
ки могут существовать вместе. Этот тип
клеточной дифференциации можно на-
блюдать, например, у Eudorina и Volvox
(см. рис. 172), а также у некоторых ци-
лиат (например, Zoothamnium alternans;
см. рис. 315а).
Гаплоидные протисты, например
вольвоксовые, хризомонады, большин-
ство динофлагеллат, некоторые гипер-
мастигиды, оксимонады и Apicomplexa,
образуют гаметы непосредственно из
вегетативных клеток или после их би-
нарных делений. У Chlamydomonas
формируются двужгутиковые изогаме-
ты, которые несут на поверхности жгу-
тиков видоспецифичные и специфич-
ные для типа спаривания гликопротеи-
ны. Эти гликопротеины вызывают аг-
глютинацию жгутиков у совместимых
(+/-) клеток. Клетки объединяются в
пары, соединяясь друг с другом при
помощи небольшой папиллы. После
слияния клеток зигота развивается в
толстостенную покоящуюся стадию, в
которой идет мейоз с последующим
формированием гаплоидных вегетатив-
ных клеток.
Более крупные колониальные Vol-
vocida (Eudorina, Volvox) оогамны. По-
ловое размножение у Volvox carteri
инициируется так называемым белком-
индуктором (гликопротеин 30 кД), ко-
торый синтезируется в ответ на повы-
шение температуры окружающей сре-
ды. Он эффективен в очень низких
концентрациях, около 6х10~
,7
М. Поэто-
му количество индуктора, выделяемое
одной колонией, бывает достаточным
для того, чтобы все колонии в конкрет-
ном водоеме начали готовиться к поло-
вому процессу. В присутствии полово-
го индуктора гонидии приступают к
своего рода эмбриогенезу. Гаплоидная
вегетативная клетка дает начало либо
множеству микрогамет, либо одной не-
подвижной макрогамете. У некоторых
видов Volvox оба типа гамет созревают
в одной колонии (например, Volvox glo-
bator); у других в этом участвуют раз-
ные колонии одного клона (например,
Volvox aureus) или колонии разных кло-
нов (например, Volvox perglobator).
Первое деление зиготы сопровождает-
ся мейозом, а дальнейшие митотичес-
кие деления ведут к формированию ко-
лоний, состоящих из гаплоидных веге-
тативных клеток.