Хаусман К., Хюльсман H., Радек Р. Протистология
Подождите немного. Документ загружается.

408 Избранные главы общей протистологии
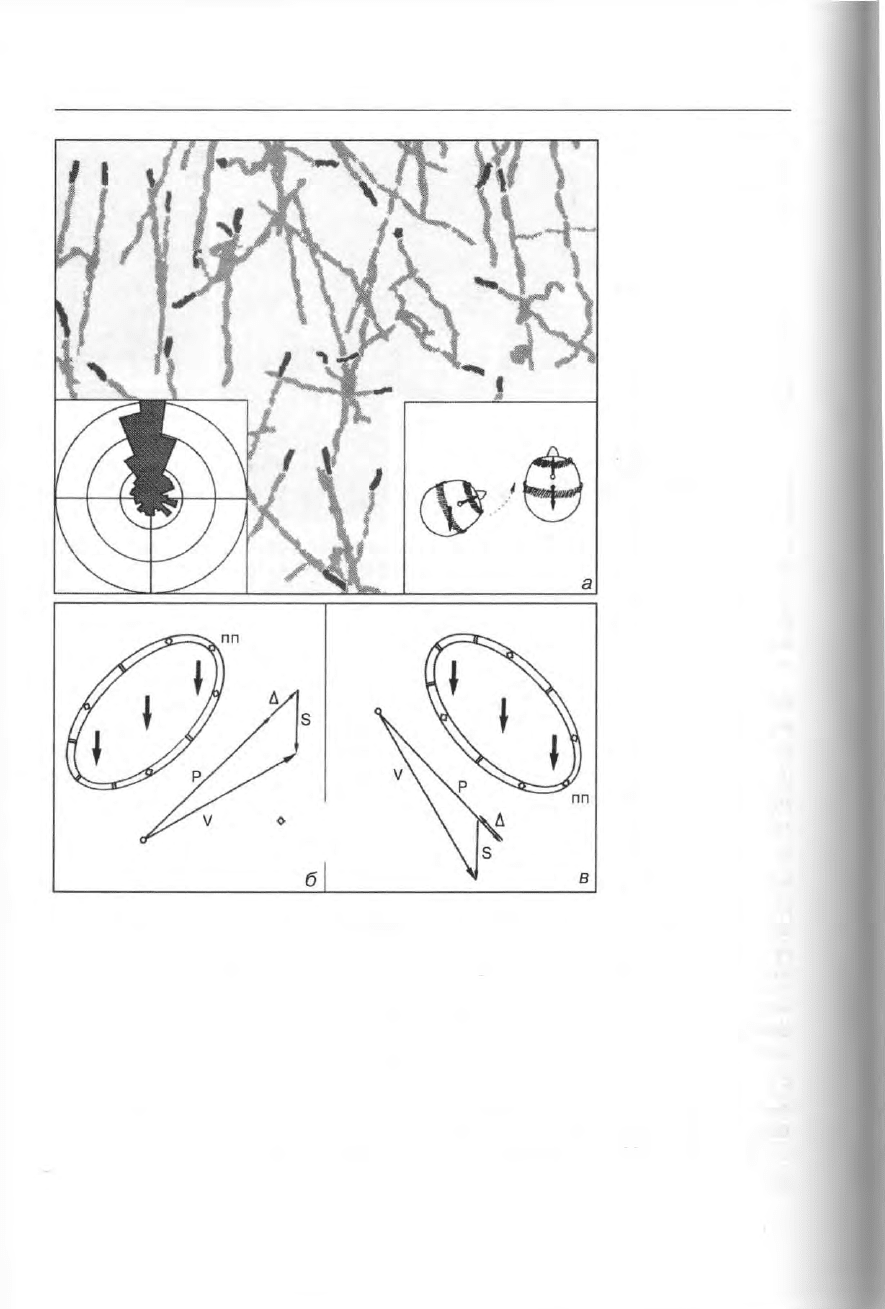
Рис. 344. Реакция ин-
фузорий на гравита-
цию: а — негативный
гравитаксис у Didi-
nium. Затемненные
окончания траекто-
рий движения пока-
зывают, что боль-
шинство клеток при
движении ориенти-
рованы вверх (см. ги-
стограмму траекто-
рий слева). Это пове-
дение можно объяс-
нить (правая врезка)
тем, что центр тяжес-
ти у Didinium (черный
кружок) находится
позади геометриче-
ского центра прило-
жения движущего
усилия (светлый кружок), б, в — гравирецепция и кинезы при движении вверх (б) и вниз
(в), по-видимому, основаны на более высокой плотности цитоплазмы по сравнению с ок-
ружающей средой (пп — передний конец клетки). В мембране имеются чувствительные к
механическому воздействию каналы двух типов: Са
2+
-зависимые и Ю-зависимые; откры-
тие первых вызывает деполяризацию мембраны, а вторых — гиперполяризацию (см. рис.
342). б — при движении вверх давление цитоплазмы на нижнюю мембрану стимулирует
сосредоточенные здесь гравирецепторы гиперполяризации. Соответственно, клетка уве-
личивает силу тяги (Р — propulsion) на величину Д. Скорость перемещения (V) — это век-
торная сумма Р, Д и скорости опускания (S) под воздействием силы тяжести, в — у кле-
ток, плывущих вниз, преимущественная активация гравирецепторов деполяризации вы-
зывает уменьшение силы тяги (Р - Д). Численное значение Д (гравикинез) рассчитывает-
ся из измерений V, Р и S. Данные по поведению многих видов инфузорий указывают, что
Д обычно компенсирует действие гравитации, т.е. представляет собой отрицательный гра-
викинез (из: Machemer and Braucker: Acta Protozool. 31 [1992] 185).
Са-канал
i К-канал

Поведение протистов 409
тикальное скольжение вверх сменяется
строго вертикальным скольжением вниз.
Тем самым,популяция Loxodes удержи-
вает свое пространственное положе-
ние', то есть проявляет нейтральный
гравитаксис.
Механическое и физиологическое
воздействие земного притяжения на
клетку обусловлено направленной вниз
силой, которая является результатом
взаимодействия веса тела (масса х g),
силы сопротивления движению (тре-
ние + динамическое сопротивление. —
Прим. ред.) и архимедовой силы. Вели-
чина этой направленной вниз силы
зависит от разницы в удельной плотно-
сти между клеткой и окружающей жид-
костью и от формы клетки
2
. Сила тяже-
сти воздействует и на специализирован-
ные органеллы, представляющие собой,
вероятно, гравирецепторы (тельце Мюл-
лера у Loxodes и родственных инфузо-
рий; см. рис. 116, 117).
Эксперименты по определению ско-
рости пассивного (под воздействием
силы тяжести) оседания различных ин-
фузорий показали, что скорость актив-
но движущихся вверх клеток увеличи-
вается по сравнению с плывущими го-
ризонтально, а у плывущих вниз умень-
шается, то есть проявляется гравикинез.
Такое поведение объясняют следую-
щим образом: при движении вверх ци-
топлазма клетки оказывает давление на
мембрану нижней стороны тела и де-
формирует ее (рис. 3446), что вызыва-
ет изменение проводимости механоре-
цепторов (рис. 342) и соответствующее
изменение биения ресничек (см. рис.
339). Судя по всему, отрицательный гра-
викинез противодействует оседанию
плавающих протистов.
Еще две реакции — реотаксис и
тигмотаксис — также обусловлены ме-
ханической стимуляцией. Многие пла-
вающие инфузории способны двигать-
ся против течения (положительный ре-
отаксис). Наиболее простое объяснение
реотактического поведения постулиру-
ет возникновение крутящего момента
из-за того, что точка приложения равно-
действующей силы смещена относи-
тельно центра тяжести (см. гравитак-
сис). Тигмотаксис — это проявляемое
организмом стремление «прилипать» к
твердой поверхности при контакте с
ней. Решающим в тигмотаксисе являет-
ся скорее молекулярное взаимодействие
клетки с поверхностью, нежели просто
деформация ее чувствительной мембра-
ны, т.к. инфузории отличают одни твер-
дые предметы от других.
Реакции на температуру
Известно, что особи Paramecium
обычно скапливаются в зоне оптималь-
ной температуры, которая соответству-
ет для них температуре предыдущего
культивирования. Инфузории увеличи-
вают скорость перемещения и уменьша-
ют количество реверсий при движении
к зоне оптимальной температуры, и на-
оборот, при удалении от оптимума ско-
рость движения падает и чаще наблю-
1
Описанное поведение популяция Loxodes проявляет в экспериментах, в которых микрокосм
сотрясают или же помещают в центрифугу для того, чтобы создать контролируемое ускорение. —
Прим. ред.
2
Имеется в виду скорость погружения по закону Стокса. — Прим. ред.

410 Избранные главы общей протистологии
даются реверсии. Таким образом, инфу-
зория осуществляет вероятностное сме-
щение по градиенту стимула. Управля-
ет этим смещением электрофизио-
логический механизм, объясненный
выше.
Paramecium при нагревании изме-
няет распределение трансмембранного
потенциала вдоль переднезадней оси
клетки. Когда температура в экспери-
менте была ниже температуры культи-
вирования, прикладывание подогревае-
мой проволочки к переднему полюсу
клетки приводило к деполяризации
мембраны, а касание той же проволоч-
кой заднего полюса вызывало гиперпо-
ляризацию мембраны. Изучение элект-
рофизиологических основ термочув-
ствительности у Paramecium показало,
что, так же как у беспозвоночных и по-
звоночных животных, терморецепция
сходна с механорецепцией: терморецеп-
торные каналы, расположенные на пе-
реднем конце клетки, Са
2+
-зависимы и
деполяризуют мембрану, а ^-зависи-
мые терморецепторные каналы, распо-
ложенные на заднем конце клетки, вы-
зывают гиперполяризацию мембраны.
Поэтому какое-то время думали, что
одни и те же проводящие каналы,
встроенные в клеточную мембрану,
обеспечивают и механическую, и тер-
мическую чувствительность. Однако
позже это предположение было опро-
вергнуто. Термо- и механорецепторы
реагировали противоположным обра-
зом на повышение температуры: ток
через канал терморецептора уменьшал-
ся, а механорецептора — увеличивался.
Таким образом, клетка протиста имеет
самостоятельные системы проводящих
каналов для преобразования термиче-
ского и механического сигналов, но в
обоих процессах задействует сходные
молекулярные механизмы.
Реакции на химические
раздражители
В процессе передвижения клетки
попадают в растворы разных химичес-
ких веществ, концентрация которых
также различна. При малых размерах
простейшим, по-видимому, трудно реа-
лизовать программу таксиса, то есть
определить химический градиент, ис-
пользуя какой-либо механизм момен-
тальной оценки направления на источ-
ник сигнала, чтобы затем приступить к
строго координированному перемеще-
нию. Считается, что реакции протистов
на химические раздражения всегда яв-
ляются кинезами, то есть на фоне хао-
тического движения устанавливается
тенденция к смещению по градиенту
стимула, причем протист периодически
оценивает интенсивность раздражителя
(см. раздел «Кинезы»), Эксперименты
на Paramecium показали, что некоторые
органические вещества (ацетат, лактат,
фолиевая кислота, инозинмонофосфат и
цАМФ) действуют на этих инфузорий
как аттрактанты. Предполагается, что
эти химические вещества маркируют
скопления бактерий. Отпугивающим
действием обладают, в частности, кви-
нидин, ВаС1
2
и КОН. Как именно дейст-
вуют эти репелленты, неизвестно. Вряд
ли для их распознавания существуют
специфичные рецепторы; эффект от
воздействия репеллента зависит от того,
каков был состав среды до его внесения
и насколько клетка была адаптирована
к этому составу раствора.

Поведение протистов 411
Реакции на химические раздражите-
ли у протистов различаются по задей-
ствованным механизмам: (1) изменение
концентрации неорганических ионов в
среде непосредственно воздействует на
трансмембранный потенциал; (2) органи-
ческие сигнальные молекулы и 0
2
свя-
зываются с рецепторами клеточной мем-
браны, и тогда внутриклеточные мессен-
джеры либо изменяют проводимость
мембранных каналов, либо включают
определенный ионный насос; (3) клетка
приспосабливается к новой интенсивно-
сти сигнала по мере продвижения в сто-
рону увеличивающейся или уменьшаю-
щейся концентрации. Было показано, что
привыкание Paramecium к измененной
концентрации КС1 достигается за счет
открытия или закрытия определенного
количества К
+
-каналов, в результате чего
возмущение трансмембранного потенци-
ала снимается и восстанавливается зна-
чение потенциала покоя.
Перемещение спорозоитов Plasmo-
dium являет пример хемотаксиса. После
выхода из ооцисты они пассивно дрей-
фуют в гемолимфе и затем, привлечен-
ные определенными химическими веще-
ствами, мигрируют в слюнные железы
комара. Перемещение при этом имеет
характер кинеза: в целом хаотично ме-
няя направление движения, спорозоит
отслеживает концентрацию сигнального
вещества и постепенно смещается в зону
оптимальной его концентрации.
В то время как химические сигна-
лы являются решающими при обнару-
жении пищи, сами пищевые частицы
инфузории, по-видимому, отбирают по
размеру. Paramecium поглощает несъе-
добный материал, например частички
кармина, угля и железные опилки так
же, как и бактерий. Didinium, который
питается преимущественно парамеция-
ми, игнорируя большинство других ин-
фузорий, распознает жертву при контак-
те. Блуждая по всему объему воды хищ-
ник случайно натыкается на потенци-
альную жертву. Считается, что после
столкновения из его хоботка выстрели-
вают специфичные экструсомы, с помо-
щью которых жертва оценивается «на
вкус». При одобрении выбора выстре-
ливают уже смертоносные токсицисты,
которые немедленно выводят из строя
механизм электроконтроля биения рес-
ничек Paramecium.
Взаимное узнавание клеток, компе-
тентных к половому взаимодействию, во
многих случаях достигается при участии
сигнальных молекул (см. рис. 307), ко-
торые либо встроены в мембрану ресни-
чек (Paramecium), либо секретируются в
среду, где служат ориентиром для хемо-
кинеза полового партнера (Blepharisma,
Euplotes). У Chlamydomonas задейство-
ваны оба эти механизма. Выделяемые в
среду аттрактанты (аттрактанты инфузо-
рий называют гамонами) принадлежат к
гетерогенному семейству половых феро-
монов, которые действуют с высокой
специфичностью в крайне низких кон-
центрациях (> 10
ч
М) и давно известны
у насекомых. У протистов после взаимо-
идентификации половых партнеров про-
исходит агглютинация жгутиков или рес-
ничек и прекращение их работы, затем
следует слияние клеток.
Реакции на свет
Многие протисты, в том числе и
бесцветные формы, реагируют на види-
мый свет. При усилении освещенности

412 Избранные главы общей протистологии
Amoeba proteus увеличивает скорость
передвижения (положительный фотоки-
нез), но через 10 минут адаптируется к
новым условиям, и скорость ее локомо-
ции падает до нормальной. Как содер-
жащие хлореллу особи Paramecium bur-
saria, так и лишенные ее скапливаются
в освещенных участках (фотоаккумуля-
ция), где они снижают скорость плава-
ния и даже вовсе останавливаются (от-
рицательный фотокинез). Это поведе-
ние характеризуется суточным ритмом
(фотоаккумуляция проявляется сильнее
днем, нежели ночью. — Прим. ред.).
Показано, что изменение поведения
здесь контролируется изменением вели-
чины мембранного потенциала покоя
(при этом водитель ритма сугубо эндо-
генный. — Прим. ред.). При очень вы-
сокой интенсивности белого света Para-
mecium bursaria и бесцветная Parame-
cium multimicronucleatum скапливаются
в затененных местах за счет увеличения
скорости движения и уменьшения чис-
ла реверсий в освещенной зоне (поло-
жительный фотокинез). Для Paramecium
определили, что фототрансдукция спек-
трочувствительна, то есть деполяризу-
ет или гиперполяризует клеточную мем-
брану в зависимости от спектрального
состава света. Это наблюдение указыва-
ет, что механизм фотокинеза соответ-
ствует общей схеме: воздействие на
трансмембранный потенциал изменяет
рисунок в целом хаотического переме-
щения инфузории. Деполяризующая
фоточувствительность Paramecium bur-
saria была максимальной в мембране
тела клетки в пределах вестибулума.
Гиперполяризующая мембрану фото-
чувствительность отмечена у интакт-
ных клеток, но эта реакция отсутство-
вала у лишенных ресничек особей.
Можно предположить, что при фото-
трансдукции именно мембрана ресни-
чек обеспечивает гиперполяризующую
реакцию. И в мембране тела клетки, и
в мембране ресничек при помощи ме-
ченых антител были обнаружены ро-
допсиноподобные фоторецепторные
молекулы.
На границе света и тени перепад в
интенсивности освещенности иногда
достигает величины нескольких поряд-
ков. Пересекающие эту границу фото-
чувствительные клетки испытывают
шок и проявляют реакцию избегания.
Механизм реакции избегания при фото-
шоке был изучен у жгутиконосца СЫа-
mydomonas (рис. 345). При восприятии
светового сигнала фоторецептор депо-
ляризует мембрану и генерирует увели-
чение концентрации Са
2+
. Быстрый рост
концентрации Са
2+
в жгутиках до уров-
ня в 10" М вызывает переключение
биения жгутиков с ресничного типа,
толкающего клетку вперед, на жгутико-
вый, толкающий клетку назад. Нор-
мальное биение восстанавливается пос-
ле того, как уровень Са
2+
в жгутиках
возвращается в норму (10~
8
М). Реакция
фотошока известна и у инфузорий, в
том числе не содержащих фотосимби-
онтов. Весь этот каскад процессов от
восприятия раздражителя до локомо-
торного ответа очень напоминает реак-
цию избегания Paramecium (см. рис.
255), и, думается, этот механизм прин-
ципиально сходен у всех ресничных и
жгутиковых протистов.
Скопление пигмента под плазма-
леммой вовсе не обязательно присут-
ствует у клетки, которая способна про-
являть фотокинез или реакцию фотошо-
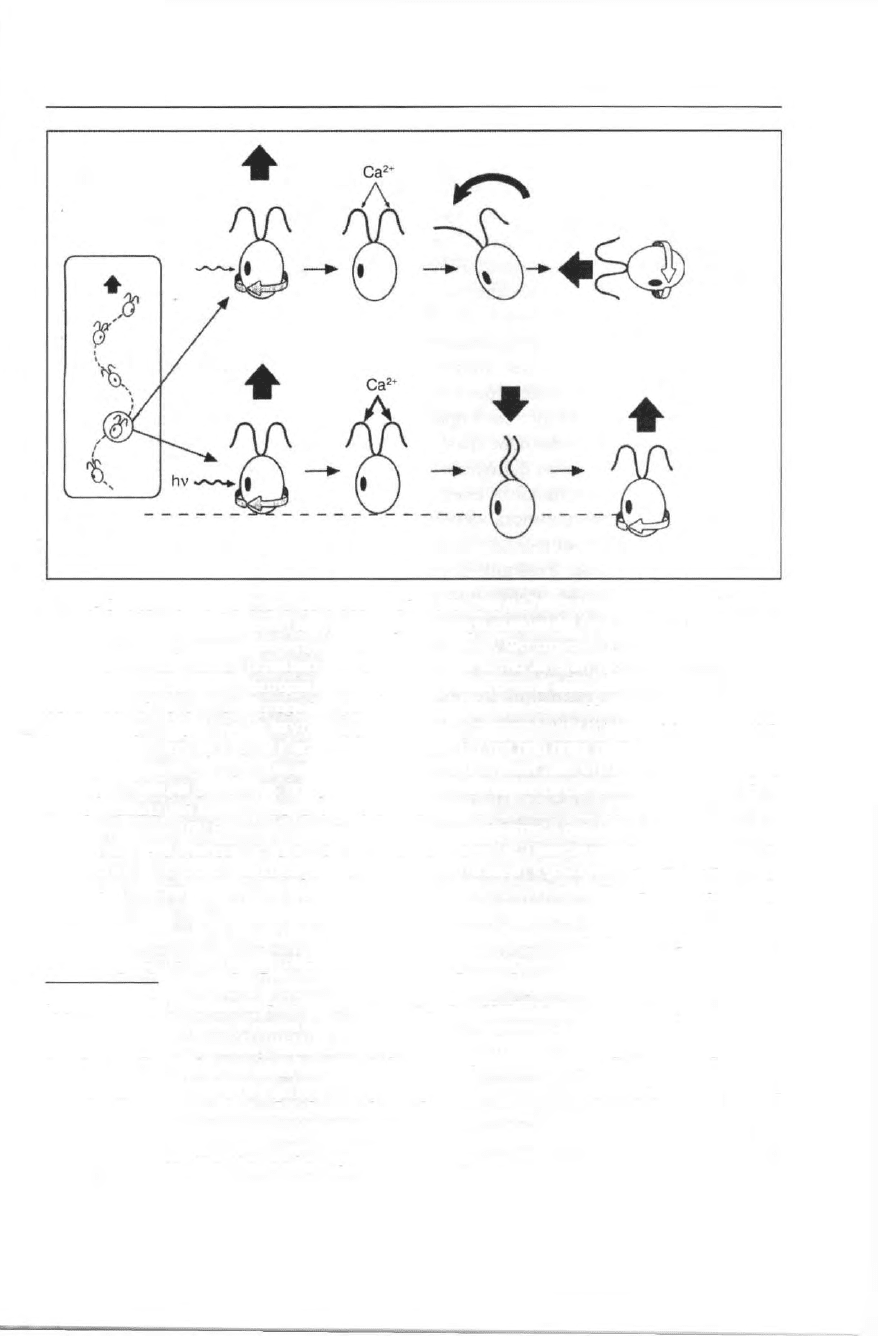
Поведение протистов 413
|фототаксис
Фотошок
а б в г
Рис. 345. Сравнение фототаксиса с реакцией фотошока у Chlamydomonas. Двужгутиковые
клетки двигаются по винтовой спирали вокруг вектора смещения (врезка). Глазок на эква-
торе клетки непрерывно сканирует окружающую среду. Фототаксис: при своем вращении
клетка однажды примет позицию, когда пигментное пятно затенит фоторецептор (а). Из-
менение освещенности фоторецептора родопсинового типа вызывает деполяризующий
рецепторный потенциал, который порождает потенциал действия, за которым, в свою оче-
редь, следует вброс Са
2+
в жгутики (б). Различие в двигательном ответе жгутиков на Са
2
*
приводит к повороту клетки к источнику света (в). До тех пор пока освещенность фоторе-
цептора не меняется при вращении клетки, она продолжает двигаться параллельно пучку
света (г). Реакция фотошока: при резком скачке интенсивности света (а) потенциал дейст-
вия, открывающий кальциевые каналы, столь велик, что концентрация Са
2+
в жгутиках превы-
шает пороговую (б). В результате оба жгутика переключаются с ресничного типа биения на
жгутиковый, что приводит к попятному движению клетки (в). Прекращение действия стимула
или адаптация к продолжающемуся воздействию света ведет к удалению избытка Са
2+
из жгу-
тиков, что восстанавливает их нормальную активность (г) (из: Witman: TICB 4 [1993] 403).
ка, однако такое пятно, по-видимому, дающих фототаксисом
1
. Перемещение,
всегда присутствует у протистов, обла- координированное по отношению к ис-
1
Для организма, проявляющего только фотокинез, не имеет значения, в каком направлении рас-
положен источник света. У такого протиста световоспринимающая система будет включать только
фоторецептор(ы). Организму, обладающему фототаксисом, необходимо определять направление
на источник света. Организация световоспринимающей системы неизбежно будет более слож-
ной: она должна включать и фоторецептор, и экран (либо светопоглощающий, либо отражаю-
щий). Фоторецептор и экран часто разнесены в клетке, хотя, разумеется, их расположение ско-
ординировано. Экран ярко окрашен и хорошо заметен при наблюдении в световой микроскоп.
Именно экран, безотносительно того, расположен ли фоторецептор рядом или на удалении, на-
зывается глазок, глазное пятно или стигма. — Прим. ред.

414 Избранные главы общей протистологии
точнику света, изучалось у фототроф-
ных протистов. У Chlamydomonas име-
ется глазок; этот жгутиконосец при ло-
комоции вращается вокруг своей про-
дольной оси, в том числе когда он на-
правляется к источнику света (рис. 345).
Глазок расположен у экватора клетки и
содержит родопсиноподобный белок, к
которому с одной стороны прилегают
поглощающие свет гранулы. Вслед-
ствие этого, когда свет падает сбоку на
клетку, вращающуюся вокруг свой про-
дольной оси, светопоглощающие грану-
лы периодически затеняют фоторецеп-
тор. Изменение интенсивности света,
вероятно, меняет проводимость канала
фоторецептора, что ведет к сдвигу мем-
бранного потенциала. Реакция двух
жгутиков на изменение мембранного
потенциала и вброс Са
2+
не идентична.
В результате траектория искривляется.
Клетка продолжает поворачивать до тех
пор, пока вектор ее смещения не при-
мет положение, параллельное лучам
света. Биение жгутиков становится пол-
ностью симметричным, и клетка плы-
вет прямо к источнику света (положи-
тельный фототаксис) или от него (отри-
цательный фототаксис).
Среди инфузорий фототаксисом до-
стоверно обладают несколько пигменти-
рованных форм — Stentor, Fabrea и
Chlamydodon — и один бесцветный ги-
стиофаг — Ophryoglena. Фотореакции
последней весьма примечательны. Ее
фототаксис меняется в ходе жизненного
цикла (рис. 346). Теронты — активно-
плавающая фаза цикла — обладают по-
ложительным фототаксисом. Поглотив
пищу, теронты превращаются в крупных
трофонтов и через несколько часов осе-
дают на дно, становясь томонтами, кото-
рые обладают уже отрицательным фо-
тотаксисом. Расселительная стадия
Ophryoglena, по-видимому, не имеет ни-
какой фоточувствительности. Предпола-
гается, что эта инфузория определяет
направление на источник света при по-
мощи так называемой либеркюновой
органеллы, расположенной в области ве-
стибулума. К этой яркой светопреломля-
ющей выпукло-вогнутой структуре с
одной стороны прилегают световоспри-
нимающие кристаллы или гранулы.
ч> ч>
0° о°
270°
180° 180°
Теронты Томонты
Рис. 346. Смена положительного фототак-
сиса на отрицательный у инфузории
Ophryoglena в зависимости от стадии жиз-
ненного цикла. Круговые гистограммы по-
казывают пропорции траекторий движения
по отношению к падающему белому свету
(стрелки вверху) у теронтов и томонтов
Ophryoglena (из: Kuhlmann: Europ. J.
Protistol. 29 [1993] 344).

Экология протистов 415
Экология протистов
Исследователь, изучающий эколо-
гию протистов, сталкивается с целым
рядом специфических трудностей.
Большой помехой являются микроско-
пические размеры самих объектов. Так-
же очень сложно выявить структуру ес-
тественных сообществ протистов. Ведь
эти организмы часто населяют лишь
особые микроместообитания, в которых
быстро меняются как обилие отдельных
видов, так и таксономический состав
сообщества (рис. 347). Эфемерность
сообществ протистов создает дополни-
тельную помеху еще и потому, что их
точная таксономическая идентифика-
ция зачастую весьма затруднительна, и
выполнить определение тут же в поле-
вых условиях далеко не всегда пред-
ставляется возможным. Последнее —
как раз одно из серьезнейших препят-
ствий при изучении экологии протис-
тов, и преодолеть его удается лишь спе-
циалистам, имеющим достаточный
опыт в таксономии. Перечисленными
трудностями отчасти объясняется отно-
сительно медленный прогресс в эколо-
гии протистов. Тем не менее, накопле-
ние знаний идет и, в частности, стано-
вится все очевиднее, что протисты, осо-
бенно их планктонные сообщества,
играют в природе очень важную роль.
Среда, в которой обитают свободно-
живущие протисты, постоянно находит-
ся под воздействием как прокариотных,
так и высших эукариотных организмов,
равно как и самих протистов. Другими
словами, среда, в которой протисты ра-
стут и размножаются, находится под
влиянием не только абиогенных, но и
биогенных факторов. Примерами вкла-
да протистов в формирование поверх-
ности Земли являются не только отло-
жения силиката и карбоната кальция
(в виде их скелетов и панцирей), кото-
рые сформировали, в частности, извест-
няки Альп, но также и фотосинтетичес-
кая активность микроводорослей,кото-
рые создали современную, содержащую
кислород и озон атмосферу нашей пла-
неты.
Было бы несправедливо относить
влияние протистов на биосферу только
к минувшим эпохам. Становится все
более очевидно, что равновесие углеро-
да в биосфере в значительной степени
находится под контролем гетеротроф-
ных и фотоавтотрофных микроорганиз-
мов морского планктона.
Помимо важности участия протис-
тов в глобальных процессах формиро-
вания среды, следует помнить, что эко-
логия протистов все чаще рассматрива-
ется как дисциплина, которая изучает
процессы, определяющие пути эволю-
ции других организмов.
Факторы,
определяющие
распространение
Встречаемость и распределение
протистов определяется абиотическими
и биотическими факторами. Абиотиче-
ские факторы имеют физическую и хи-
мическую природу. К биотическим фак-
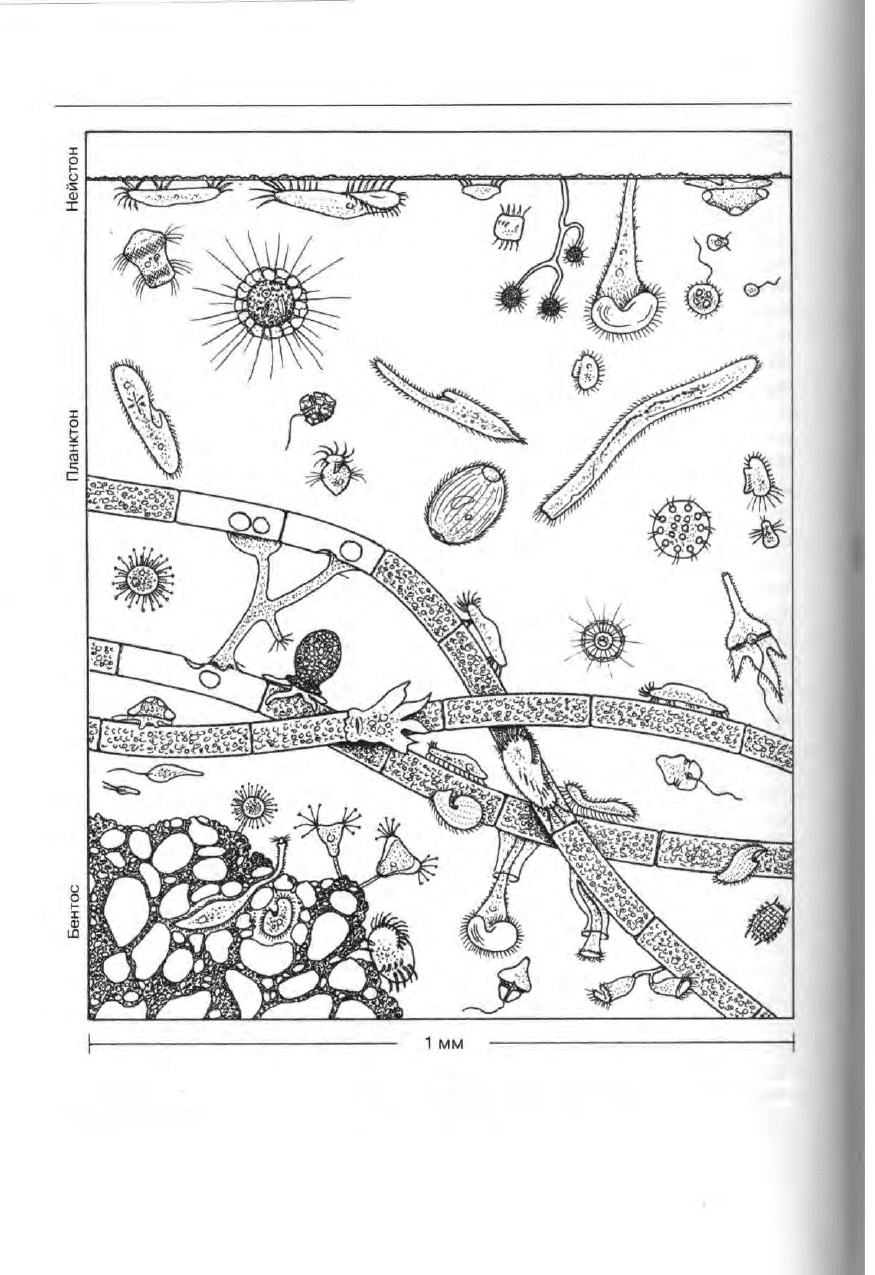
416 Избранные главы общей протистологии
Рис. 347. Схема пространственного распределения одноклеточных организмов в водной
среде. Представители всех групп свободноживущих протистов обитают на поверхностной
пленке, в планктоне, в бентосе, на водных организмах (по нескольким авторам).
Экология протистов 417
торам относятся: наличие пищи, конку-
ренция, взаимоотношения типа хищ-
ник—жертва и др. Комбинация абиоти-
ческих факторов определяет, смогут ли
конкретные виды протистов существо-
вать в данном биотопе, тогда как био-
тические факторы более важны в опре-
делении относительного обилия той
или иной популяции.
Абиотические факторы
Протисты обнаруживаются во всех
биотопах и местообитаниях. Хотя каж-
дый вид толерантен к ограниченному
диапазону значений конкретного факто-
ра, несколько видов охватят весь био-
сферный диапазон изменчивости этого
фактора (рис. 348). Информация о вли-
янии абиотических факторов на проти-
стов большей частью получена в ходе
лабораторных экспериментов и в мень-
шей степени — при полевых наблюде-
ниях. Абиотические параметры, изме-
ренные в естественных местообита-
ниях, относятся обычно к большим
объемам воды и далеко не всегда харак-
теризуют микробиотоп, в котором оби-
тает популяция определенного вида.
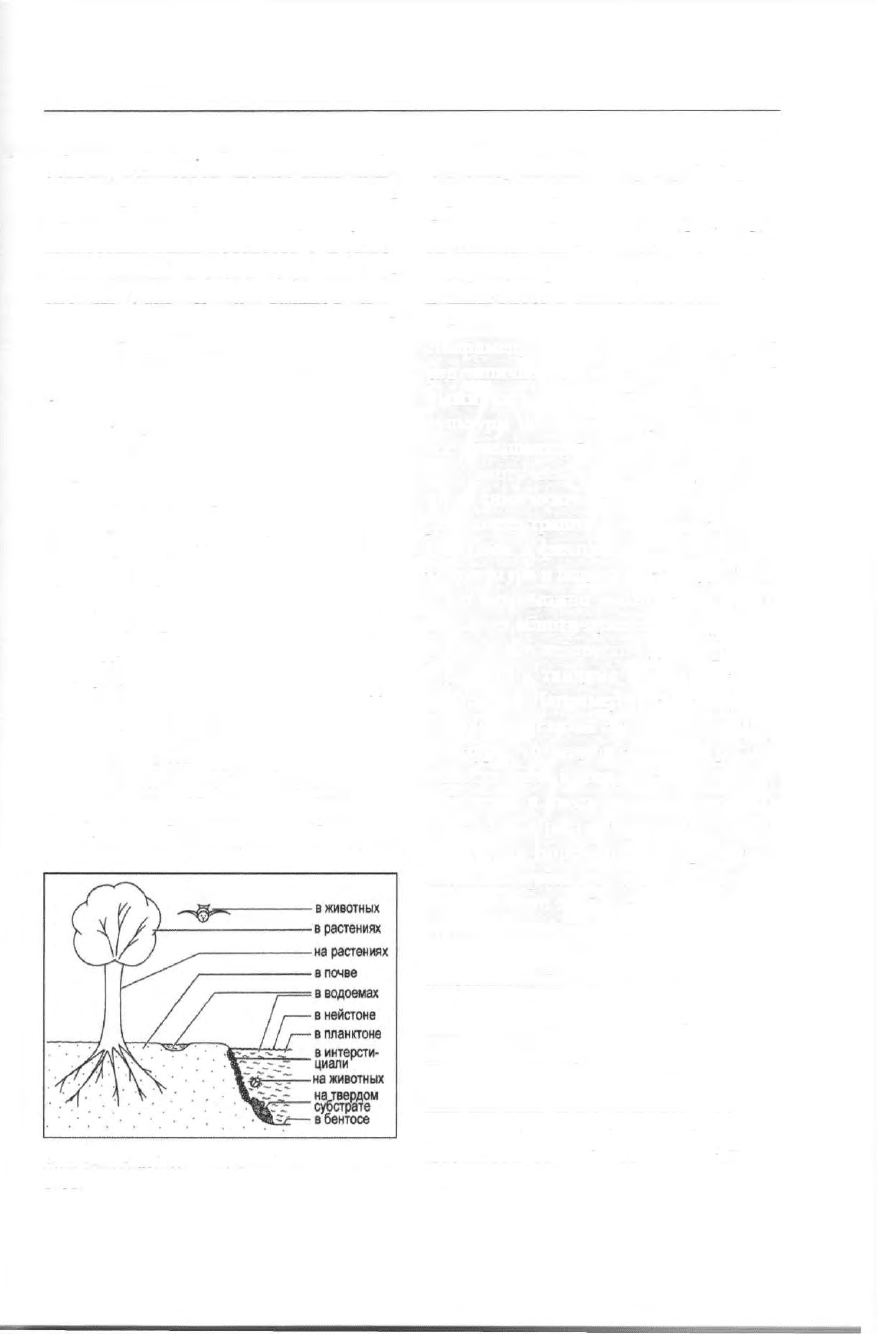
Рис. 348. Наиболее важные биотопы проти-
стов.
Из лабораторных экспериментов
известно, что цисты инфузорий Colpoda
переживают очень низкую температуру.
Это отнюдь не означает, что род Colpo-
da типичен для арктических областей.
С другой стороны, отсутствие опреде-
ленных видов в данном местообитании
ничего не говорит об их потребностях.
Например, было показано, что целый
ряд типично пресноводных инфузорий
выживают в морской воде, если их
культуры постепенно адаптировать ко
все повышающейся солености.
Абиотические факторы подразделя-
ют на химические, такие как влажность,
рН, концентрация ионов и растворен-
ных газов, и физические, такие как свет,
температура и подвижность воды. Зача-
стую невозможно выделить вклад от-
дельного абиотического фактора, ско-
рее, они все действуют однонаправлен-
но, часто в тандеме с биотическими
факторами. Например, рост освещенно-
сти обычно связан с повышением тем-
пературы, что ведет к снижению раство-
римости газов в воде, росту уровня ме-
таболизма и увеличению потребности
протистов в растворенном кислороде.
Даже если определенные организмы
способны временно переживать эти ус-
ловия, ничего нельзя сказать об их ко-
нечных шансах на выживание. Толерант-
ность Paramecium multimicronucleatum
к повышенной температуре увеличива-
ется только с одновременным уменьше-
нием обилия пищи (бактерий) или уве-
личением рН среды. Как бы то ни было,
базовые представления о наиболее важ-
ных лимитирующих факторах, регули-
рующих встречаемость протистов, явля-
ются необходимой предпосылкой для
понимания их экологии.