Kenneth E. Gonsalves, Craig R. Halberstadt, Cato T. Laurencin, Lakshmi S. Nair. Biomedical Nanostructures
Подождите немного. Документ загружается.

This chapter aims to provide an up-to-date review of some of the significant
developments in our understanding of the ECM and its interactions with cells
from the macro- to nanoscale.
9.2 CELL MICROENVIRONMENT
Cells have diameters typically in the tens of microns and are not the only
constituents of tissues. A large portion of tissue volume is often occupied by
extracellular space, which contai ns a dynamic three-dimensional network of
macromolecules that assemble to form the cell microenvironmentthe ECM.
Constant remodeling alters the constituent molecules and thus dictates the
properties of the cellular environment.
The ECM is a diverse composite structure of cell-secreted proteins,
glycosaminoglycans (GAGs), proteoglycans, and glycoproteins. Tissue-specific
heterogeneity in its histoarchitecture has evolved to serve numerous functions
ranging from supporting cells to intracellular signaling. Numerous proteins
play a role in the activation of signaling pathways, with cell surface receptors
serving as primary conduits for signal transduction between neighboring cells,
secreted signaling molecules, and ligands in the surrounding matrix. These
extrinsic factors extend profound influences on both gene expression and cell
behavior and ultimately come together to form a dynamic cell microenviron-
ment essential for tissue development and function.
9.2.1 ECM Compositional Diversity
Although the major constituents of ECM found in all metazoa can be grouped
into four classes: collagens, glycoproteins, GAGs, and proteoglycans,
evolutionary pressures have resulted in substantial diversity in the ECM
structure. The many variations in ECM form may be explained by the
numerous variants of each constituent class and the ability of cells to secrete
different proportions of these ECM components, which then form various
architectural configurations. ECM compositional diversity reflects the broad
range of biofunctional requirements of the different organ systems.
9.2.1.1 Constituent Macromolecules Extensive evolutionary changes
have occurred in metazoa, generating diversified matrix molecules with different
structural properties and signaling domains. For example, fibrillar type I
collagens found in tendons form rope-like structures to carry tensile loads [8],
while nonfibrillar type IV collagens form a meshworks that act as size-selective
filtration barriers in the basement membrane [9]. Many of the constituent
macromolecules are the result of chimeric aggregations of structural domains
that have formed by exon shuffling [10] and numerous isoforms have resulted
through differential splicing of these domains and posttranslational modifica-
tions [11]. Collagens, for instance, have achieved considerable diversity, as more
226 BIOMEDICAL NANOSTRUCTURES
than 27 distinct types have been characterized [12, 13]. Similar diversity through
differential splicing has been found in numerous glycoproteins of the ECM,
including fibronectins, laminins and tenascins, whereas GAG variation and
number account for proteoglycan heterogeneity [11, 14].
9.2.1.2 Developmental Diversity In addition to the specific diversity
found in the individual constituent macromolecules of the ECM, the matrix as
a whole, can display considerable compositional diversity. Through differ-
ential production of particular ECM components, specific regions or
boundaries within the tissue may be delineated.
ECM regulation of cell behavior has been observed in a number of
developmental processes [5, 6, 15–17]. As developing cells move among
microenvironments within the developing tissue, each with different markers/
signals, they modulate their differentiating state and behavior accordingly.
ECM signaling generates a unique biological and structural ‘‘niche’’ that
directs cellular differentiation and maintains cellular phenotype. This cellular
modulation is accompanied by dramatic changes in the types, ratios, and
geometrical arrange ments of the synthesized ECM macromolecules, giving rise
to the specific biofunctional pro perties of the tissue in question.
Endochondral ossification is one developmental process that requires the
coordination of a vast array of microenvironments and cell types. In the
beginning, the hyaluronate- and type I collagen-rich matrix provides a
supportive environment for mesenchymal cell migration and proliferation
[18]. As bone development progresses, the mesenchymal cells migrate and
aggregate into condensations that form the template for future skeletal
elements [19, 20]. The cells at the center of these condensations then
differentiate into chondrocytes, which subsequently leads to the removal of
hyaluronate from the surrounding microenvironment and the secretion of large
amounts of macromolecules including aggrecan and collagen II [21]. These
macromolecules act as molecular markers that help distinguish the differ-
entiating chondrocytes from the surroun ding undifferentiated mesenchymal
cells. Once fully differentiated, further complex transitions occur in the
arrangement and composition of the cartilag inous ECM. The ECM
surrounding the chondrocytes calcifies and the cells apoptose [17]. This
calcified matrix favors vascular invasion from the bone collar and allows
chondroclast and osteoblast invasion [15]. At this point, collagen synthesis
reverts from type II to type I as the surrounding cartilaginous ECM degrades
and is replaced with bone ECM [21]. Changes in composition are therefore
important factors in the control of organogenesis as well as provide suitable
structural properties to the developing skeletal tissues.
9.2.1.3 Tissue-Specific Diversity In complex organisms, the ECM of
distinct tissue types has evolved into different forms. This is significant since
the structure and function of a specific tissue will depend on the relative
proportions of the constitutive molecules present. This is evident in tissue types
ECM INTERACTIONS WITH CELLS FROM THE MACRO- TO NANOSCALE 227
like articular cartilage or arterial walls, where ECM compositional diversity
helps to engender their inherent structure and properties.
The unique blend of macromolecules, together with the ECM configuration,
gives articular cartilage its impor tant physical propertiescompressive
stiffness and friction-reduced movement of joints. In articular cartilage, the
predominant collagen, type II, along with smaller amounts of other collagens
(V, VI, IX, X, and XI), forms a fibrillar network [22] that is embedded with
multimolecular aggregates that self-assemble from the proteoglycan, aggrecan,
and the GAG, hyaluronan [23]. The fibrillar structure of the collagen network
provides tensile strength to the tissue and resistance to the swelling pressures
of the embedded proteoglycans, whilst the aggrecan surface, being highly
sulfated and negatively charged, attracts cations to the matrix and in turn
increases the osmotic swelling pressure [24]. This swelling prop erty allows the
dissipation of compressive loads through changes in osmotic and hydrostatic
pressures.
Unlike the functional requirements of cartilage tissue, arterial walls must
resist tensile forces, provide a barrier to blood cells, and be elastic enough to
accommodate hemodynamic pressure [25] associated with blood flow from the
heart. A hierarchical arrangement of endothelial cells (ECs), smooth muscle
cells (SMCs), and ECM proteins come toget her to form the arterial walls in
vascular systems [16]. Arterial walls are organized into three distinct layers: the
intima, consisting of ECs and a hydrated matrix rich in hyaluronan and
proteoglycans; the media, separated from the intima by a dense internal elastic
lamina and composed of layer s of SMCs and elastic fibers; and the adventitia,
separated by an elastic lamina and composed primarily of fibrillar collagen
and fibroblasts [26]. The integrated composition of collagens, fibrillins, and
elastins help maintain vascular homeostasis and impart specific properties to
the arterial walls. The predominant collagens type I and III assemble
into fibrils that provide tensile strength to the walls [16], whereas type
IV and VIII collagens within the basement membranes under the ECs, form
three-dimensional networks that serve to anchor the cells and help control
vascular permeability [26]. Fibrillins self-associate to form microfibrillar
networks that stabilize elastic fiber structure [27, 28] and elastins assemble
into elastic fibers that provide elasticity needed to accommodate hemodynamic
pressure differences from blood flow [29].
ECM compositional diversity has therefore evolved as a mechanism to
guide cell behavior through the organizational arrangement of developmental
stage- and tissue-specific structures with the biofunctional properties of these
tissues being dependent on the biochemical composition and architectural
composition.
9.2.2 Nanoscaled Structures of the ECM
Interactions between cells and ECM occur over multiple length scales, where
the hierarchical assembly of the various ECM constituents plays an essential
228 BIOMEDICAL NANOSTRUCTURES
role in numerous cellular processes. Whether it is an intricate glycopr otein
network pro viding adhesion points for cell binding and migration or a
negatively charged matrix-embedded proteoglycan providing charge-selective
filtration properties to the ECM, the constituent molecules that mobilize to
form the nano- to macroscale structures of the ECM can be grouped into four
major groups: proteins (collagens and elastins), GAGs, proteoglycans, and
glycoproteins.
9.2.2.1 ProteinsCollagens and Elastins Numerous proteins are con-
stituted in the ECM; however, the primary ones are collagens and elastins.
Collagens are the most abundant proteins in the human body and comprise a
highly diverse superfamily of ECM proteins. Of the 27 different collagen types
(IXXVII), 42 distinct a-chains have been described [12, 30–33]. All collagen
molecules consist of three homo- or heterotrimeric a-chain subunits that wind
together to form triple-helix conformations [34]. Each a-chain subunit contains a
(Gly-X-Y)
n
repetitive sequence motif, where glycine is present every third
position and X and Y are often the amino acids proline and hydroxyproline [35].
Hydroxyl groups in hydroxyproline are essential in forming H-bonds that
stabilize the collagen triple-helix conformation, whereas lysine and hydroxyly-
sine residues present in the structure are integral in forming covalent cross-links
within and between collagen molecules in supramolecular form [36]. Most
collagens associate into highly organized supramolecular structures and their
triple-helical domains can vary in length and can be continuous or interrupted
with nonhelical domains depending on collagen type.
The collagen superfamily can be divided into several subfamilies based on
supramolecular structure and other properties. Table 9.1 presents a summary
of these subfamilies and their known supramolecular structures.
The synthesis of collagen pro-alpha chains occurs by several co- and
posttranslational modifications, in addition to established pathways for
secretory proteins. These include the hydroxylation of secreted prolines and
lysines, the glycosylation of selected hydroxylysine and asparagines residues,
disulfide bond formation, and chain self-assembly into the procollagen triple
helix. The procollagens are then secreted into the ECM where the protective N
and C propeptides are cleaved and the collagen molecules are assembled into
stable cross-linked fibrils or other supramolecular structures depending on
collagen type. Further details of collagen biosynthesis may be found in the
following reviews [13, 32, 36] and text [37, 40]. This section, however, will
instead focus on the hierarchical structures of collagen and how they self-
organize to interact with cells.
Collagens are primarily responsible for the structural integrity of ECM or
the anchoring of cells to the matrix and have a tendency to form fibrils,
filaments, or networks alone or in conjunction with other ECM proteins.
Fibril-forming collagens are normally associated with the structural
functions, whereas nonfibrillar collagens are often involved in the regulatory
functions.
ECM INTERACTIONS WITH CELLS FROM THE MACRO- TO NANOSCALE 229
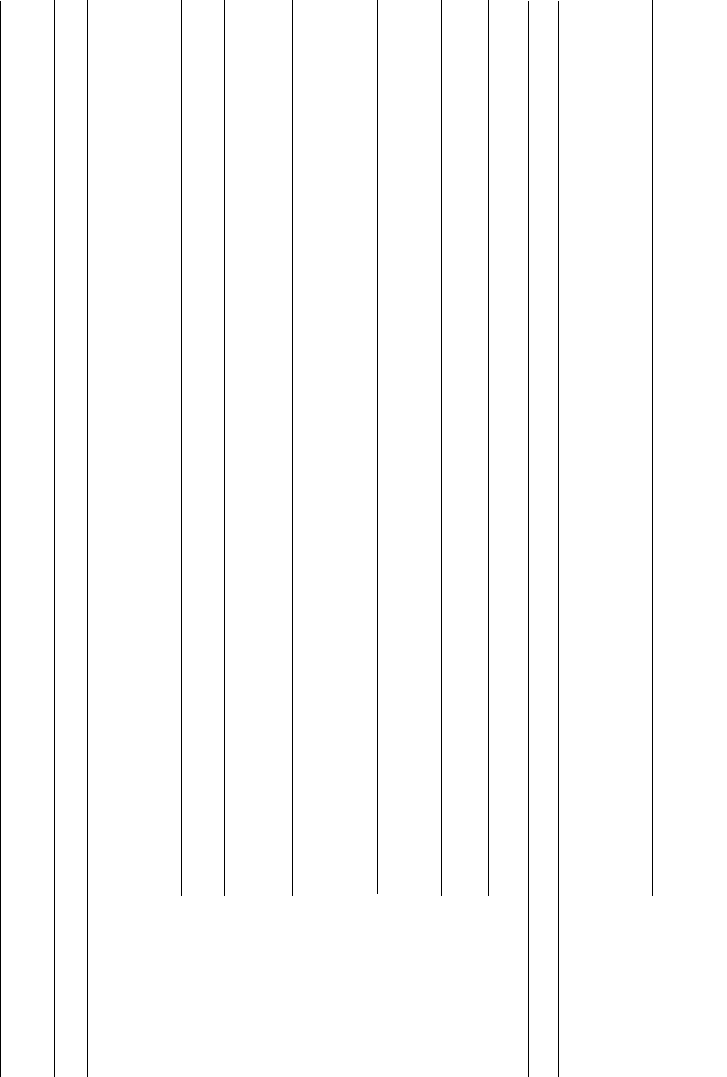
TABLE 9.1 Collagen Subfamilies and Known Supramolecular Structures
Subfamilies
Collagen
type Structural features Representative tissues Function
Fibrillar collagens
Fibril forming I 300 nm-long, staggered
large-diameter
banded fibrils
Skin, tendon, bone,
lung, cornea
Supporting fibers
recognized by
integrin cell
surface receptors
II 300 nm-long, staggered
small-diameter banded fibrils
Cartilage, vitreous
humor
Supporting fibers
III 300 nm-long, staggered
small-diameter fibrils; often
copolymer with type I
Skin, muscle, vascular,
lung, kidney
Small supporting
fibers
V 390 nm-long fibrils with globular
N-terminal extension; often
copolymers with type I and III
Cornea, teeth, bone,
skin, smooth muscle,
tendon, liver
Small supporting
pericellular fibers;
interacts with
thrombospondin
XI Small-diameter fibrils;
can form molecules
with type II, IX and V
Cartilage, vitreous
humor
Possible core for
type II regulating
fibril diameter
XXIV Small-diameter fibrils;
coexpressed with type I
Developing
cornea and bone
Possible regulator
of type I fibrillogenesis
XXVII Small-diameter fibrils Cartilage, eye, ear, lung Unknown
Nonfibrillar collagens
FACIT
a
and related
structural collagens
IX FACIT; N-terminal
globular domain;
bound GAG
Cartilage, vitreous humor Type II collagen
fibril association and
interaction with
proteoglycans
230

XII FACIT Skin, tendon,
cartilage, periodontal
ligament
Type I collagen
association and
interaction with
different ECM
components
XIV FACIT Fetal skin,
tendon, cartilage
Similar to type XII
XVI FACIT Heart, lung, pancreas,
placenta
Stabilizing fibroblasts
in dermal matrix
XIX FACIT Vascular, neuronal, and
mesenchymal basement
membrane zones
Cell-cell/cell-matrix
adhesive protein;
association with
heparan sulfate
proteoglycans
XX FACIT Corneal epithelium, skin,
cartilage, tendon
Regulation of fibrillar
diameter; similar to
collagen XII and XVI
XXI FACIT Vascular walls, smooth
muscle cells
Participation in ECM
assembly of vascular
networks during
angiogenesis
XXII FACIT Tissue junctions Cell adhesion
ligand for
skin epithelial cells
and fibroblasts
XXVI FACIT Testis, ovary Possible role in
development of
reproductive tissues
231
(continued )

TABLE 9.1 (Continued )
Subfamilies
Collagen
type Structural features Representative tissues Function
Sheet forming IV 400 nm-long fibrils
forming meshwork
sheets
Basement membrane Meshwork scaffolding;
multiple cell-binding
sites
VIII 130 nm-long fibrils
forming hexagonal
network
Sclera, Descemet’s
membrane
Regulation of matrix
environment; fibroblastic
cell layers and
Descemet’s membrane
thickness
X 130 nm-long fibrils
forming hexagonal
network
Hypertrophic
cartilage
Endochondral bone
development; associates
with type II
collagen fibrils
Beaded filaments VI 510 nm-diameter beaded
filaments; periodic
globular domains
Vessels, skin,
intervertebral disc
Interfaces with major
collagen fibrils and cells;
multiple RGD motifs
recognized by integrin
receptors
Anchoring VII 420 nm-long collagenous
domains; forms
anchoring filaments
Subbasal l
amina of skin
Anchoring of dermal
epithelial cells to
underlying stroma
Transmembrane XIII Cell membrane protein Fibroblasts, cartilage
growth plate,
epidermis, hair
follicles
Cellular
adhesion and
migration; binds to
fibronectin, heparin and
basement membrane
components
232

XVII Cell membrane protein Dermalepidermal
junction of skin in
hemidesmosomes
Linkage of basal
cells to stroma
XXIII Cell membrane protein Metastatic tumor cells,
cornea, lung
May assist in enhancing
the stability of corneal
interfacial areas
XXV Cell membrane protein Neurons May play a role in adheren
junctions between neurons
Multiplexins XV Core protein of
chondroitin sulfate
Heart, placenta,
endometrium, skeletal
muscles
Regulates critical
basement membrane
functions; stabilizes
muscle cells and
microvessels
XVIII Core protein of
chondroitin sulfate
Lung, liver, kidney Regulates critical basement
membrane functions;
maintains structural
integrity
Data complied from multiple references and the references therein [13, 30, 32, 35–39].
a
Fibril-associated collagens with interrupted triple helices (FACIT).
Fibril-forming collagens (I, II, III, V, XI, XXIV, and XXVIII) represent
major products synthesized by connective tissue cells and have the unique
ability to associate into highly ordered fibrils and fibers. Their structural unit
generally consists of a 300 nm-long, 1.5 nm-diameter triple helix, and the
presence of the triple-helical domain provides these fibrils with stiff, rod-like
regions. In addition, fibrils may be heterotypic aggregates of more than one
collagen type organized into discrete tissue-specific configurations where their
structure and biomechanical properties are governed by the specific ratios of
collagen involved [35]. For example, diameter control was found to be an
inherent property of the specific proportions of collagen type for heterotypic
fibrils of embryonic cartilage formed from collagen types II, IX, and XI [41].
Interactions with fibril-associated collagens with interrupted triple-helix
(FACIT) collagens and small proteoglycans have also been shown to regulate
fibril properties [42]. FACITs (IX, XII, XIV, XVI, XIX, XX, XXI, XXII, and
XXVI) are another subclass of collagens that do not form fibrils themselves,
but are found attached to the surface of preexisting collagen fibrils. They are
comprised of relatively short triple-helical (collageneous) domains connected
by nontriple-helical (noncollageneous) sequences. FACITs help modulate
ECM structure by forming molecular bridges between fibrillar collagens and
other ECM components. Specifically, FACITs like collagen XVI may play an
active role in anchoring microfibrils to basement membranes [43], whi le others
like GAG carrying collagen IX help modulate the surface charge properties of
fibrils [44].
Sheet-forming collagens (IV, VIII, and X) like those involved in forming
basement membrane structures (IV) or hexagonal networks (VIII and X) can
act as anchorage for cells, serve as molecular filters, or provide permeable
barriers for developing embryos. Type IV collagen molecules in the basement
membrane have collageneous domains longer than those of fibril-forming
collagens, 400 nm long [45], and can self-assemble to form cross-linked
network structures in which the monomers associate in end-to-end interactions
and the helical domains intertwine to form super- coiled structures [36]. The
other sheet-forming collagens type VIII and X are made up of almost the same
sized 130-nm-long rod base units, where the collageneous domains contain
eight imperfections in similar positions in the Gly-X-Y sequences [30] and
assemble to form hexagonal lattices of type VIII or X collagen.
Beaded filament collagens (VI) are heterotrimeric molecules made up from
three unique protein chains with very short triple-helical domains and large N-
and C-terminal globular domains, which assemble into beaded microfibrils
[36]. Type VI is the only known collagen to form beaded filaments and the
presence of arginineglycineaspartate (RGD) motifs within its structure
plays a central role in its interaction with other ECM constituents like
fibronectin and with integrin cell surface receptors [30].
Anchoring collagens (VII) form anchoring fibrils that link the basement
membrane to anchoring plaques in the underlying ECM [46]. Type VII collagen
is a major component of anchoring fibrils, and the 420 nm-long triple-helical
234 BIOMEDICAL NANOSTRUCTURES
domains are longer than any other collagen [30]. It is a homotrimeric molecule
consisting of collagenous central domains flanked by two noncollageneous
domain ends. Within the extracellular space, these type VII collagen molecules
form antiparallel dimers, which then assemble laterally in a nonstaggered
manner to form anchoring fibrils [47].
Transmembrane collagens (XIII, XVII, XXIII, and XXV) are homotrimeric
cell surface molecules with multiple extracellular triple-helical domains that are
connected to single hydrophobic transmembrane domains [35]. Within their
ectodomains, which reside in the extracellular space, the triple-helical domains
provide rigid rod-like structures, whereas the interruptions between these
helical regions provide structural flexibility [48]. These collagens function as
cell surface receptors and their ectodomains can be cleaved off from the cell
surface to produce soluble extracellular molecules.
Multiplexins (XV and XVIII) are homotrimeric collagens with multiple
triple-helical domains and interruptions. They have a large N-terminal
globular domain, a highly interrupted triple helix, and a large C-terminal
globular domain. In addition, multiplexins often carry GAG chains and are
therefore referred to as proteoglycans.
For more comprehensive coverage of structure and function of the collagen
superfamily, the reader is referred to previous reviews [13, 30, 32–34, 36, 48, 49].
Elastin is the major protein of elastic fibers in connective tissues like blood
vessels and skin and is extremely rich in the hydrophobic amino acids glycine,
valine, alanine, and proline [50, 51]. It is synthesized when the enzyme lysyl
oxidase covalently cross- links its soluble precursor, tropoelastin, and forms an
insoluble polypeptide [29]. Elastin, in association with other microfibrillar
components, assembles to form elastic fibers, which then form a randomly
oriented interconnected network. Elastin fibrillogenesis begins as 1012-nm-
diameter fibrillin microfibril bundles are secreted into the extracellular space
near the cell surface and assemble into aggregates [29]. Fiber development
progresses, as tropoelastins deposited at specific locations within the microfibril
bundles progressively displace the microfibrils to the outer region of the fibers
and covalently cross-link to form the elastin core. These elastin fibrous
networks are highly insoluble and have the ability to undergo elastic recoil [52].
The unique intermolecular desmosine and isodesmosine cross-links and the
high content of hydrophobic amino acids within elastin’s structure account for
these properties [53, 54]. Additionally, elastin peptides are known to enhance
signaling pathways, with Val-Gly-Val-Ala-Pro-Gly (VGVAPG) known to
upregulate metalloproteinases [55] and promote chemotactic activity with
fibroblasts [56].
9.2.2.2 GAGs GAGs are long polymer chains of specific repeating
disaccharides that are important in regulating biological functions. This
may consist of creating permissive environments for cell migration and
macromolecular transport through regulation of the size and properties of
ECM spaces or binding to signaling molecules like growth factors to
ECM INTERACTIONS WITH CELLS FROM THE MACRO- TO NANOSCALE 235