Kenneth E. Gonsalves, Craig R. Halberstadt, Cato T. Laurencin, Lakshmi S. Nair. Biomedical Nanostructures
Подождите немного. Документ загружается.

modulate signaling pathways. This group of complex carbohydrates is
classified based on the nature of their repeating disaccharide units and
includes chondroitin sulfate, dermatan sulfate, heparan sulfate, hyaluronan,
and keratan sulfate [57]. Due to their high sulfate content and/or the presence
of uronic acid, GAGs bare many negative charges that enable them to
associate with numerous ligands by electrostatic interactions. All GAGs are
found in tissues as covalently attached components of proteoglycans, with the
exception of hyaluronan. Hyaluronan is unique since it is secreted directly
into the extracellular space by the cell membrane bound enzyme hyaluronan
synthase and is able to function as a f ree carbohydrate [58, 59]. It is flexible,
bending and twisting into many conformations forming random coils, and
can self-associate to form networks. It has a strong affinity to water due to the
large number of anionic residues on its surface and can bind cations by COO
groups on its surface. Additionally, it can form superaggregates with a
number of proteoglycans including aggrecan.
9.2.2.3 Proteoglycans Proteoglycans are a diverse group of molecules
that are abundant in the ECM and expressed on cell surfaces. They represent
a glycoprotein subset in which structure and function are mediated by protein
cores and covalently linked GAG side chains. Numerous peptide motifs exist
within these molecules and many of these are often found in the same protein
core including epidermal growth factor (EGF) repeats, hyaluronan-binding,
immunoglobulin-like, leucine-rich repeats, and sugar-binding lectin domains
[60]. Several classes are present in the ECM and these may be divided into
two subfamilies based on the typical features of the protein core arrangement
(Table 9.2) [61].
The first family, the small leucine-rich proteoglycans, includes proteoglycans
like biglycan, decorin, fibromodulin, and lumican. While the second family, the
modular proteoglycans, can be further divided into the nonhyaluronate-
binding proteoglycans like perlecan and agrin and the hyaluronan/lectin-
binding proteoglycans like aggrecan, versican, and CD44 [2]. This diversity in
protein core domains and GAG side chains enables proteogl ycans to have a
high affinity to binding of various ligands and accounts for many specific
interactions in the matrix. Perlecan, for instance, is embedded in basement
membranes and gives them a fixed negative electrostatic charge, which is
responsible in part for their charge-selective filtration properties [63]. Whereas
the small leucine-rich proteoglycan, decorin, can bind and regulate the
fibrillogenesis of collagen [64, 65], other proteoglycans, like heparan sulfate
proteoglycans, participate in fibroblast growth factor (FGF) signaling by
directly interacting with FGFs and its receptors on the cell surface,
subsequently increasing FGF’s affinity for its associated receptors [57, 66].
9.2.2.4 Glycoproteins With the ability to organize collagens, elastins,
proteoglycans, and cells into ordered structures, glycoproteins are also essential
building blocks of the ECM. These macromolecules vary in size, structure, and
236 BIOMEDICAL NANOSTRUCTURES

TABLE 9.2 General Properties of Selected Proteoglycans
Proteoglycan
Protein core
(kDa)
GAGs
(number)
Representative
tissue Function
Small leucine-rich proteoglycans
Biglycan 3840 Chondroitin
sulfate/
dermatan
sulfate (2)
Muscle, bone,
cartilage,
epithelia
Associated
with cell
surfaces
Decorin 3640 Chondroitin
sulfate/
dermatan
sulfate (1)
Collagenous
matrices, bone,
teeth
Binds collagen
fibrils and
modifies
their assembly;
binds to
TGF-beta
Fibromodulin 4243 Keratan
sulfate (24)
Cartilage,
skin, tendon
Binds collagen
I and II;
reduces size
of collagen
fibrils
Modular proteoglycans
Nonhyaluron
an binding
Agrin 200250 Heparan
sulfate (3)
Synaptic
sites of
neuromuscular
junctions,
renal
basement
membranes
Involved in
the aggregation
of acetylcholine
receptors
Perlecan 400467 Chondroitin
sulfate/
heparan (3)
Basement
membranes,
cell surfaces,
cartilage
Self-associates;
binds laminin
in basement
membranes;
binds basic
fibroblast
growth
factor
Hyaluronan
and
lectin
binding
Aggrecan 220250 Chondroitin
sulfate (>150)
and Keratin
sulfate
(100
150)
Cartilage,
brain,
blood
Binds
hyaluronan,
hydrates and
fills
ECM
Versican 260370 Chondroitin
sulfate/
dermatan
sulfate
(1030)
Blood
vessels,
brain,
cartilage,
skin
May bind
hyaluronan
Data complied from multiple references [2, 40, 61, 62].
ECM INTERACTIONS WITH CELLS FROM THE MACRO- TO NANOSCALE 237
distribution and are composed of a protein and covalently linked oligosac-
charide(s). Glycoproteins are made up of multiple subunits colored with an
assortment of distinct peptide motifs that interact with not only themselves, but
also cells and other matrix proteins [6]. They may exist as large molecules with
extended conformations that span distances of several hundred nanometers. A
variety of glycoproteins are found in the ECM including fibronectin and
laminin.
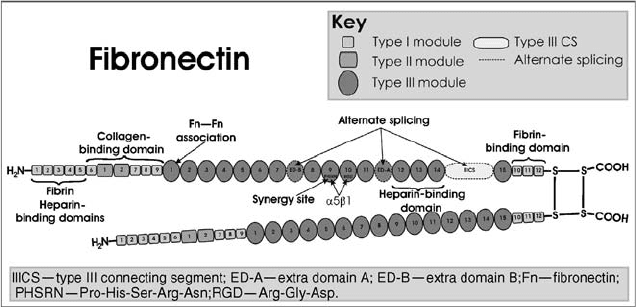
Fibronectin is one of the best studied glycoproteins, has multiple
isoforms, and is composed of three modules, repeating motifs of type I, II,
and III, separated by connecting sequences [67]. It is a large glycoprotein
with numerous binding sites for collagens, proteoglycans, and cell surface
receptors like integrins (Fig. 9.1). The type III module is the most abundant
module in its structure and includes many aspartate-containing sequences
such as RGD, which mediate cellular adhesion through integrins [68, 69].
Fibronectin is critical for numerous cellular functions and is one of the
earliest macromolecules to be laid down in the ECM. It is required for the
assembly of several ECM proteins like collagen I [70] and plays a role in
regulating the incorporation of morphogens and growth factors into the
ECM [71]. Its distribution in areas of skeletogenesis suggests that it may be
involved in early stages of bone development [72] as well as continued
mature bone homeostasis by regulating the differentiation and survival of
osteoblasts [73].
Laminins are the primary adhesive matrix proteins in the basement
membrane, exist in several isoforms, and are large heterotrimeric glycoproteins
constituted by three genetically distinct polypeptide alpha, beta, and gamma
FIGURE 9.1 Schematic representation of fibronectin dimer. Depicts individual modules
as well as a variety of binding domains specialized for particular macromolecules.
Alternate splicing accounts for the differences in the two similar chains, which are joined
by two disulfide bonds near the C-termini.
238
BIOMEDICAL NANOSTRUCTURES
chains that assemble to form a cruciform structure with globular domains at
each arm end (Fig. 9.1) [74, 75]. They contain multiple binding domains that
participate in numerous biological activities (network and filament formation,
cell binding through cell surface receptors, and binding to other matrix
proteins in supramolecular assemblies), with the ECM structure-forming
domains generally located on the short arms of all three subunits and cell-
interacting domains mapped onto the N- and C-terminal domains of the alpha
subunits [74]. For example, within the ba sement membrane, all three short
arms (alpha1, beta1, and gamma1) of laminin-1 are involved in laminin self-
assembly [76], while binding with entactin/nidogen is restricted to the short arm
region of the gamma1 chain [77], and integrin cell surface receptors to the
laminin G-domain-like (LG) modules of the alpha subunits [78, 79]. Such
binding interactions are essential for proper assembly and function of the
basement membrane, where type IV collagen, laminins, and entactins are
thought to be the primary basement membrane scaffold, fibrous two-
dimensional supramolecular networks where entactin binds the laminin and
collagen networks with stable noncovalent bridges [78, 80].
9.2.3 Putting It All Together—Hierarchical Assembly
Hierarchy of scales is apparent in the biological structures formed during tissue
development and homeostasis. Within the ECM, the multifunctional nature of
matrix components is a direct reflection of their modular and hierarchical
architecture. The mult iple binding domains, chemistry, and topology of the
various ECM constituents help regulate matrix assembly, stabilize ECM
architecture, and modulate cellmatrix interactions. Some examples include
fibrous structures that provide strength, elasticity, or cell permissivity,
proteoglycans that act as reservoirs for morphogens or regulate osmotic
pressure, and glycoproteins that stabilize fibrous structures or act as binding
sites for cells. All of these structures combine together to give tissues their
specialized properties. The significance of this relationship between ECM
hierarchy and function is apparent in tissues like articular cartilage.
Articular cartilage is an avascular tissue containing a relatively sparse number
of chondrocyte cells that covers the subchondral bone in diarthrodial joints [22].
In association with synovial fluid, it forms a wear-resistant, friction-reduced,
load-bearing surface only a few millimeters thick [81, 82]. Its matrix and cells are
organized into distinct zones: superficial, middle, deep, and calcified zones
(Fig. 9.2) and these zones exhibit variations in biochemical composition, cell
morphology, and cellmatrix structural organization [81, 83].
The superficial zone consists of a densely packed collagen fibrillar network
with flattened chondrocytes that are alig ned tangentially with the articulat-
ing surface [84]. Collagen fibril alignment within this zone enables the
accommodation of high shear and tensile forces and type II collagen form s the
bulk of the fibrils. Microfibril assembly is mediated in part by type XI collagen
and decorin with the former regulating lateral growth and the latter regulating
ECM INTERACTIONS WITH CELLS FROM THE MACRO- TO NANOSCALE 239
fibril diameter [41]. Fibrils are thinnest in this zone and measure approximately
20 nm in diameter [83].
The middle zone represents a transitional region from the tangential fiber
orientation of the superficial zone to the radial fiber orientation of the deep
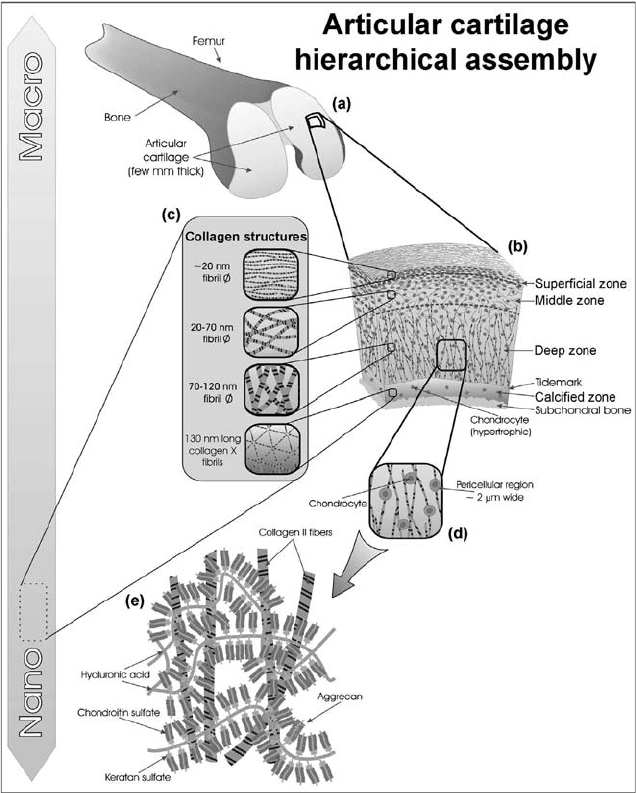
FIGURE 9.2 Hierarchical organization of articular cartilage over different length
scales. Articular cartilage forms a wear-resistant, load-bearing surface that covers bone
in diarthrodial joints (a). It is organized into distinct zones (b), where the organization
of the collagen structures varies between zones (c). Resident chondrocytes are encased in
pericellular regions (d), which are surrounded by a well-defined matrix nanoarchitecture
of aggrecan/hyaluronic acid superaggregates and macrofibrillar collagen networks (e).
240
BIOMEDICAL NANOSTRUCTURES
zone. Cells exhibit a round morphology and their density is lower than that in
the superficial zone [85, 86]. The ECM within this region is aggrecan rich and
the collagen fibrils are larger in diameter, randomly arranged, and loosely
packed [84, 87].
Within the deep zone, the collagen fibers are perpendicularly oriented
with the subchondral bone and attain a maximum diameter size that ranges
from 70 to 12 0 nm [83]. Cell density is lowest in this region [86], whereas
aggrecan content is at a maximum. Cartilage compressive stiffness is provided
by the swelling pressure of hydrated aggregates of aggrecan and hyaluronan
bound to the porous collagen II structure within this zone, where swelling is
restricted by the collagen network. This hydrated fibrous proteoglycan
structure, in conjunction with synovial fluid and articulating joint movements,
allows nutrients to circulate and nourish cells [24].
The calcified zone provides a buffer region between the uncalcified
cartilage and subchondral bone. The chondrocytes that reside in this region
are in a hypertrophic phenotypic state. These cells secrete the hexagonal sheet
forming collagen type X and participate in calcifying the ECM within this
region [83].
Additionally, within the different zones a pericellular matrix region,
approximately 2 mm wide, surrounds the chondrocytes [83]. This region varies
in its structure compared to that of the surrounding matrix. Type VI collagen
and decorin are concentrated in this region and assemble to form a
microfilamentous network. The cell surface proteoglycan perlecan is also
present within this region. It cross-links with ECM components such as type VI
collagen, thereby linking cell surfaces with matrix organization.
Therefore, the highly specialized composi tion and structure of articular
cartilage has evolved to enable joint mobility and gives an excellent example of
the relationship between the ECM hierarchy and tissue function.
9.3 CELLECM INTERACTIONSTHE MULTIDIMENSIONAL MAP
Beyond its role in providing structural support and organization for resident
cells and imparting tissues with specific biofunctional properties, the ECM also
serves in an instructive capacity. A vast amount of information is encoded in
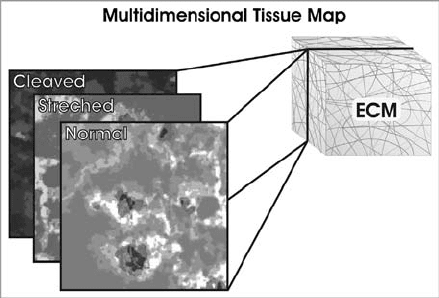
the matrix architecture and this forms a multidimensional map (Fig. 9.3). Cells
use this map to guide their behavior. The information within it is expressed in
the form of signaling gradients, which may exist as chemical distributions or
differential expressions of ECM components. Cell surface receptors interpret
these gradie nts and respond accordingly with directionality being dictated by
signaling gradient changes. A bidirectional flow of information between the
cells and the ECM ensues where changes in the local microenvironment result
in dynamic releases of information. This map may be perceived in a variety of
different ways and its context is dependent on external events as well as specific
cell types.
ECM INTERACTIONS WITH CELLS FROM THE MACRO- TO NANOSCALE 241
9.3.1 Signaling Gradients
In order to direct cells into the desired mode of behavior, signaling cues need to
be display ed within an appropriate context. This requires both spatial and
temporal positioning of signals from different sources. Consequently, each cell
type has evolved to produce their own network of precisely encoded ECM
proteins. This enables the configuration of the structural properties and
information content of distinct ECM microenvironments. The three-dimen-
sional matrix architecture assembled from ECM constituents regulates the
spatial and temporal signals that are derived from localized biochemical
distributions and mechanical forces. Biochemically, the ECM interacts with
cells to provide information on the microenvironment through soluble and
insoluble effectors, while physically it achieves this action through structural
and mechanical restraints. The mechanisms to establish gradients of signaling
molecules within the matrix architecture coincide with the deposition of ECM
constituents. This may involve sequestering effector molecules from cells,
secreting them in activated or deactivated states, or varying concentrations
within the local microenvironment. Autoregulation of these mechanisms
occurs through incremental changes in which different levels of regulation help
maintain a homeostatic balance between ECM remodeling and signaling
molecule release during processes like morphogenesis and tissue remodeling.
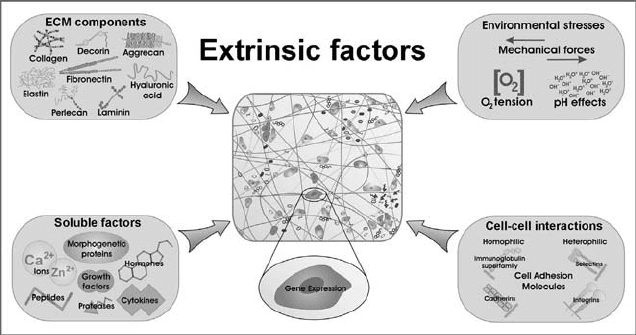
A variety of extrinsic factors control cellular behavior. The major sources of
these external signals are soluble factors, ECM components, environmental
stresses, and cellcell interactions (Fig. 9.4) [88]. The modulation and
FIGURE 9.3 Multidimensional tissue map. By providing signaling gradients to cells,
natural tissues effectively provide multiple behavioral maps, whose context is dependent
on external events. These gradients present themselves in the form of different signaling
molecules or binding opportunities that result from changes in the ECM through
processes such as stretching and proteolytic cleavage. (Modified from Reference 7 with
permission from AAAS).
242
BIOMEDICAL NANOSTRUCTURES
presentation of these external factors within the local ECM microenvironments
is of equal importance to the presence of the external signals themselves. This
chapter, however, will primarily focus on those sources associated with ECM
structures.
9.3.2 Soluble Factors
Secreted by cells, soluble factors (Fig. 9.4) feed into co mplex and overlapping
signaling pathways that often share common receptors. The modulation of
cellular activity by soluble factors is complex. Not only is the presence of the
factors required, but they must be present at the appropriate times,
concentrations, and ratios. These secreted molec ules have a tendency to
diffuse great distances, which potentially makes them unavailable or available
in ineffective concentrations [89]. Consequently, a number of mechanisms have
evolved to modulate their spatial and temporal distributions. These mechan-
isms include the production of molecules in an inactive form such as
Transforming growth factor TGF-beta [90], which activates only at the
appropriate site of action; the requirement of cofactors such as heparan sulfate
for recept or activation [57], and the localization of effector molecules like Ca
2+
by bind ing to specific matrix constituents [91]. A direct or indirect relationship
FIGURE 9.4 Extrinsic factors influencing cellular behavior. Four major sources of
external signals regulate cell behavior: soluble factors, ECM components, environ-
mental stresses, and cellcell interactions. Soluble factors include growth factors,
cytokines, morphogenetic proteins, peptides, hormones, proteases, and ions. ECM
components include proteins, GAGs, proteoglycans, and glycoproteins. Environmental
stresses include mechanical forces (dynamic and static), oxygen tension, and pH effects.
Cellcell interactions can be either homophilic or heterophilic and are regulated by cell
adhesion molecules. These extrinsic factors in turn extend profound influences on gene
expression and cell behavior, forming a dynamic cell microenvironment essential for
tissue development and function.
ECM INTERACTIONS WITH CELLS FROM THE MACRO- TO NANOSCALE 243
can exist between ECM components and many of these soluble factors. For
instance, interactions with GAGs and proteoglycans can lead to dynamic
concentrations of free signaling molecules [88]. Additionally, signaling
synergies often exist with ECMsoluble factor interactions. For example,
cell adhesion has been shown to enhance autophosphorylation of the EGF
receptor in response to its associated ligand [92]. Of the numerous soluble
factors, growth factors are indicative of the necessity for spatial and temporal
distribution in the modulation of cellular activities.
9.3.3 Growth Factors
Growth factors are specialized polypeptide molecules that bind to cell surface
receptors and trigger intracellular signaling pathways [12]. The binding of growth
factors to ECM structures is a major mechanism for regulating growth factor
activity. Growth factors are essential for establishing signaling gradients during
matrix synthesis. ECMgrowth factor association enables the storage of large
quantities of signals in readily available form. This association with matrix
structures means the signals can prevent or slow down diffusion, thereby
decreasing the loss of information [93]. Additionally, ECMgrowth factor
binding enables the transmission of signals to cells in contact with the same matrix
region later, which is essential for the development of complex cellular structures.
Not all growth factors found wi thin the matrix architecture are in an active
state. This inertness may be attributed to a variety of factors. The growth
factors may be concealed within the matrix only to be revealed during matrix
degradation or cell movement. For instance, proteolysis of ECM-bound
vascular endothelial growth factor (VEGF) by matrix metalloproteinase
MMP-9 releases and increases VEGF bioavailability [94]. Similarly, an
epidermal growth factor-like domain DIII, released throu gh cleavage of
laminin-5 by MMP-2, stimulates cell migration and changes in gene expression
only upon its degradation [95]. A group effect may also occur where multiple
signals from the matrix counteract each other and signaling activity would
depend on the ratios of various growth factors (i.e. biological activity woul d
require perturbations). For example, during angiogenesis, antiangiogenic
factors dominate until the release of angiogenic growth factors by ECM
proteinases causes a proangiogenic environment to ensue [96–100]. Finally, the
growth factors can be latent with limited biological activity. For instance,
TGF-beta is secreted by the majority of cells as latent complexes that are
unable to interact with their corresponding signaling receptors until activated
by processes like proteolysis [101].
Growth factor activity is prim arily controlled by two biochemical
mechanisms, release from the matrix architecture and activation. Release
involves the dissociation of growth factors from matrix binding sites, while
activation involves the dissociation of mature growth factors from dormant
proteins or the induction of conformal changes that allow its binding to
244 BIOMEDICAL NANOSTRUCTURES
receptors [93]. The release of growth factors from the matrix into biologically
active form may occur via proteolytic enzymes. For example, cell surfa ce
localized matrix metalloproteinases such as MMP-9 and MMP-2 can release
and activate TGF-beta by proteolytically cleaving its latent form [102]. This
proteolytic release and activation of stored growth factors generates rapid
localized signals. Proteolytic modulation is faster than sim ple control of gene
expression, which is of particular importance in wound healing and immune
system control [93].
9.3.4 ECM Components
ECM components can serve as landmarks for cells by leaving spatial and
temporal cues. As discussed earlier, as cells migrate through the microenviron-
ments within developing tissue, each with different ECM landmarks, they
modulate their behavior accordingly, adjusting gene expression in response to
the environmental cues. This creates a feedback loop as the responding cells
also influence the surrounding ECM by differentially expressing particular
ECM components and/or proteases. The stage- and tissue-specific assembly of
matrix components therefore requires an intricate interplay between the
residing cells and the signaling cues. With respect to the ECM components,
these cues may emanate from the molecular composition, that is, multiple
binding domains that may be apparent on the protein surface or concealed as
cryptic sites or from the surface topography.
9.3.4.1 Binding Domains Numerous proteins are expressed in the ECM
and each contains multiple motifs. These motifs are made up of specific amino
acid sequences that present multiple binding sites for cell interactions with many
distinct cell surface receptors. These interactions may directly or indirectly
influence cellular activities such as adhesion, migration, proliferation, differ-
entiation, and apoptosis. For instance, alpha5-beta1 integrin cell surface
receptors bind directly to RGD sites on fibronectin molecules (10th type III
repeating unit) [103], whereas tenascin indirectly interferes with cellular adhesion
by binding to fibronectin [104], thus altering its ability to promote cell adhesion.
The spatial proximity and regularity of binding domains has also been found to
elicit different cellular responses. The migration of fibroblasts has been found to
be a function of the average surface binding site density where cell speed varies
with the spatial presentation of RGD binding sites and affected cell adhesion
[105]. Colocalization of synergistic binding sites can also modulate cellular
responses. For example, the synergistic Pro-His-Ser-Arg-Asn (PHSRN) site in
the central cell-adhesive domain of fibronectin (9th type III repeating unit),
although itself not biologically active, has been found to substantially enhance
the cell-adhesive activity mediated by the alpha5-beta1 integrin cell surface
receptor to its associated RGD binding site (10th type III repeating unit) [106].
The spacing between the PHSRN and RGD sites in the native conformation of
fibronectin is approximately 4 nm [107].
ECM INTERACTIONS WITH CELLS FROM THE MACRO- TO NANOSCALE 245