Рубин А.Б. Современные методы биофизических исследований (Практикум по биофизике)
Подождите немного. Документ загружается.


где ф — электротонический потенциал, измеряемый по отношению
к
уровню потенциала
покоя,
х — расстояние по оси клетки. К за-
висит от проводимости мембраны, а также электропроводности
наружной и внутренней среды:
где г
м
— сопротивление мембраны на единицу длины; r
t
и г
о
—
сопротивление внутренней и внешней среды на единицу длины клет-
ки.
При г
0
< /-; X =
\
r
rjr
i
.
Решение уравнения
(IV.4.1)
имеет вид
ср =
ф
о
ехр(
—
х/'к).
Видно, что электротонический потенциал уменьшается с увеличе-
нием
расстояния, ослабляясь до ф
о
/е при х = X.
При
анализе переходных процессов, вызванных пропусканием
импульсов тока,
следует
учитывать, что электротонический потен-
циал является функцией
двух
переменных: координаты х и вре-
мени
/. Распределение электротонического потенциала во времени
по
длине клетки описывается уравнением
З
2
ф ф т дер
а*
2
х
2
х
2
at
где т -•• /\,С
М
— постоянная времени клеточной мембраны.
На
основе кабельной теории разработан относительно простой
метод измерения сопротивления и емкости цилиндрической клетки
конечных размеров с помощью
двух
микроэлектродов. В середи-
ну клетки, общая длина которой составляет 21, вводят токовый
микроэлектрод; точке инъекции тока
соответствует
координата
х
-— 0. Если ввести измерительный микроэлектрод на расстоянии
0,38/ от места нахождения токового электрода, то полное сопротив-
ление клеточной мембраны
будет
определяться выражением, по
форме совпадающим с законом Ома: R = rjl — U (x
uv
) I (0), где
U (*кр) — электротонический потенциал, измеряемый на критиче-
ском расстоянии х = 0,38 /; / (0) — сила тока, пропускаемого
через микроэлектрод. Сопротивление мембраны на единицу длины
г
м
имеет размерность Ом-см, R выражено в омах. Из кинетической
кривой
нарастания электротонического потенциала в точке х =
— 0,38 / при пропускании прямоугольного импульса тока можно не-
посредственно оценить постоянную времени мембраны (т). За время
т электротонический потенциал достигает 0,63 максимального зна-
чения.
Критическое расстояние 0.38/ рассчитано для клеток харо-
вых водорослей, у которых 1/К —
0,5ч-3,0.
Значение x
KV
несколько
варьирует в зависимости от сопротивления торцов клетки.
Для измерения пассивных электрических свойств волокон мож-
но
использовать и
другие
методы, однако в этом
случае
необходимо
равномерное протекание тока через клеточную поверхность. Для
этой
цели в крупные нервные волокна вводят центральный осевой
электрод, а снаружи помещают
другой
металлический электрод,
161

что обеспечивает эквипотенциальность внутренней
и
наружной
среды.
Метод
изолирующих мостиков.
Введение осевого электрода
на
всю длину клетки представляет сложную,
а
иногда
и
невыполнимую
задачу.
Гораздо проще создать равномерную плотность тока
на не-
большом участке, отделенном
от
остальной поверхности клетки
изо-
лирующими перегородками. Этот прием широко используют
при
работе
с
клетками
харовых
водорослей. Длинную изолированную
клетку помещают
в
камеру
с
тремя отсеками. Промежутки
между
отсеками заполнены вазелином, который обеспечивает электриче-
скую изоляцию
между
растворами
в
центральном
и
боковых отсе-
ках
(рис. 57). При
наложении разности потенциалов
между
боко-
выми электродами
и
электродом, находящимся
в
центральном отсе-
ке,
путь
тока замыкается через клетку.
Так как
ширина зазора
меж-
ду изолирующими перегородками
(2—3 мм)
существенно меньше
постоянной
длины
X,
плотность мембранного тока
на
центральном
изолированном сегменте клетки
будет
равномерной. Мембранный
потенциал изолированного участка
и
изменения, вызванные
про-
пусканием тока, регистрируют
с
помощью единственного внутри-
клеточного микроэлектрода.
В
опытах
с
мышечными волокнами
в ка-
честве
изолирующей среды используют раствор сахарозы. Примене-
ние
изолирующих мостиков позволяет осуществлять электрическую
стимуляцию возбудимых клеток, измерять
их
сопротивление
и ем-
кость.
Если центральный участок клетки харовой водоросли предвари-
тельно деполяризовать
0,1 М
раствором
KG. то.
пропуская
на
этом
участке
входящий
ток,
можно наблюдать гиперполяризационный
ответ, заключающихся
в
скачкообразном увеличении мембранного
сопротивления
и
переходе
мембранного потенциала
на
более отрицательный уровень.
Изменение
сопротивления
мембраны
при
гиперполяри-
зационном
ответе
(в
общем
случае
при
пропускании
по-
стоянного тока) измеряют
путем
наложения
на
фикси-
рованный
ток
дополнительной
низкоамплитудной перемен-
ной
составляющей. При проте-
кании
через мембрану элек-
трического тока
/
мембран-
ный
потенциал можно пред-
ставить
в
виде суммы омиче-
ского падения напряжения
IR
M
и
эффективной
ЭДС
мембраны
Е: V = //?„ + Е,
Рис.
57.
Трансклеточное пропускание
то-
ка
с
помощью изолирующих мостиков:
Мэ
—
измерительный микроэлектрод,
ЭС
электрод сравнения,
V -
измеряемый
мем
бранный
потенциал.
/ -
регистрируемый тран-
смембранный
ток, Ян —
сопротивление
на
грузки
(~1 ГОм), Е-
генератор прямоуголь-
ных импульсов
162

где
R
M
—
мембранное сопро-
тивление. Сопротивление
R
M
или
обратную величину
—
проводимость
g
M
—
рассчиты-
вают
по
закону
Ома: R
M
=
=
g
-i = AWA/, где AV —
смещения потенциала клетки,
вызываемые слабыми импуль-
сами тока
А/.
Определение
мембранной
емкости.
Эквивалентная
схе-
I
и
1ш
1
Рис.
58.
Эквивалентная
схема
измери-
тельной
и
токовой цепей
в
методе
изо-
лирующих
мостиков
ма рассмотренной цепи
по-
казана
на рис. 58.
Здесь
R
M
и С
м
обозначают сопротивление
и
емкость мембраны,
R
H
—
резистор, стабилизирующий
ток в
цепи.
Сопротивление утечки
R
y
отражает протекание небольшой
части тока
в
обход
клетки
по
границе
с
изолирующим слоем.
Сопротивление наружной среды
и
электрода сравнения, находя-
щегося
в
центральном отсеке, объединены
на
схеме
в
резистор
R
c
.
Включение напряжения
Е
сопровождается заряжением емкости
мембраны.
При
условии,
что R
H
> R
M
и R
y
> R
M
,
характерное
время экспоненты
т
равно /?
М
С
М
.
При
плохом качестве изоляции
между
отсеками камеры (низкое
R
y
)
экспериментальное значение
т
окажется заниженным
по
сравнению
с
постоянной времени мембра-
ны
/?
М
С
М
. Фазе плавного нарастания потенциала
до
максимума
может предшествовать небольшой быстрый скачок потенциала, обус-
ловленный падением напряжения
на
суммарном сопротивлении
среды
и
внешнего электрода сравнения
R
c
.
Падение напряжения
на
внешнем электроде сравнения
накладывается
на
измеряемый
потенциал мембраны,
что
огра-
ничивает точность измерений
со-
противления
и
емкости
по рас-
смотренным выше схемам. Ошиб-
ку измерений можно дополни-
тельно понизить, разделив
пол-
ностью токовую
и
измеритель-
ную цепи.
Для
этого вводят
до-
полнительный электрод,
а
элек-
трод сравнения изолируют
от
общего заземления
(рис. 59). _
сп
,.
г
u
r
Рис. 59.
Измерение электрических
Стандартный прием СОСТОИТ
в
парам
е
Т
ров клетки четырехэлектрод-
том,
что
потенциалы измери-
тельного микроэлектрода
и
электрода сравнения измеряют
двумя
предусилителями,
а
затем
находят разностный сигнал.
р рр
ным
методом
(с
использованием
двух
микроэлектродов):
/.
2 —
токовые электроды.
3, 4 —
измери-
тельные электроды,
5 -
электрометриче-
ские усилители, 6—дифференциальный
уси-
литель,
7 —
регистратор
тока,
/?н —
рези-
стор, стабилизирующий
ток в
цепи
163

Измерения
на
перемен-
ном
токе.
Для тестирова-
ния
пассивных электриче-
ских характеристик кле-
точных мембран наряду с
постоянным
током исполь-
зуют
переменный ток раз-
личных частот. Оба подхо-
да — анализ изменений по-
тенциала в
ответ
на про-
пускание ступени тока и
построение частотных за-
висимостей импеданса кле-
ток
— достаточно схожи,
хотя
измерения на перемен-
ном
токе в широком диапа-
зоне частот
несут
большую
информацию
об эквивалент-
ной
электрической
схеме
мембранной системы и механизмах ионного транспорта.
Для измерений импеданса с помощью четырехэлектродной схе-
мы (рис. 59) через клетку пропускают переменный ток известного
значения
и регистрируют падение напряжения на мембране, созда-
ваемое этим током.
Импедансом
мембраны!
называют ее комплексное
сопротивление для переменного тока частоты ш:
Z
= д« +
1Л.Ц).
В простейшем
случае
частотная зависимость импеданса клетки
описывается эквивалентной схемой, включающей параллельно сое-
диненные элементы R и С, а также последовательно соединенное со-
противление R
n
внутренней и внешней среды (рис. 60, А). Импеданс
параллельного участка цепи Z'. определяемый условием 1/Z' --
=
1/R + коС,
R
1
Рис.
60.
Частотная зависимость импеданса
мембраны
харовых
водорослей.
А —
экви-
валентная
схема
мембраны
с
последователь-
ным
сопротивлением
(R
u
)
внешней среды;
Б
—
зависимость модуля импеданса
(Z
u
)
от частоты переменного тока, пропускаемо-
го через мембрану
Z'
=
|+(О
2
С
2
/?
2
— I •
С
учетом
R
u
действительная часть импеданса всей цепи составит
В эксперименте измеряется
модуль
импеданса, численно равный
сумме
векторов R
a
и Х
а
: Z
M
•=
ния
для
и
Х
ш
, получим
z
м
Х'^. Подставляя выраже-
(IV.4.2)
164

Зависимость модуля импеданса от частоты, описываемая этим
уравнением, показана на рис. 60, Б. На высоких
частотах
Z
M
=
r
- R п.
т
-
е
-
все
напряжение падает на водном растворе, окружаю-
щем клетку. На постоянном токе Z
M
= R
n
+ R- Частотно-зави-
симый спад импеданса определяется ростом проводимости кон-
денсатора с повышением частоты. Для линейного участка спада
Z
M
в логарифмических координатах угловой коэффициент близок
к
— 1, т. е. при возрастании частоты на один порядок происходит
10-кратное уменьшение импеданса. Соблюдение данного условия
в
эксперименте
свидетельствует
об адекватности упрощенной
эквивалентной схемы мембраны для рассматриваемого частотного
диапазона.
По
измерениям импеданса на линейном
участке
спада Z
M
можно
рассчитать емкость мембраны. Формула для расчета получается
из
соотношения
(IV.4.2):
=
to/?
Простейшая эквивалентная
схема
клетки часто оказывается
недостаточной для объяснения электрических свойств клеток на
переменном токе. В соответствии с этим кривые зарядки мембранной
емкости при пропускании постоянного тока в ряде
случаев
лучше
аппроксимируются суммой
двух
экспонент. Для интерпретации
свойств растительных клеток привлекают сложные эквивалентные
схемы. Это обусловлено существованием
двух
последовательно
расположенных мембран — плазмалеммы и тонопласта, а также
неоднородностью свойств мембраны в пределах одной клетки. Про-
странственная неоднородность емкости и проводимости установлена
для клеток
харовых
водорослей, которые в определенных условиях
формируют у поверхности чередующиеся кольцевые зоны выделе-
ния
и поглощения Н
+
. Дополнительным фактором, объясняющим
расхождение частотных характеристик целой клетки и простей-
шего электрического аналога,
служит
зависимость емкости клеточ-
ных мембран от частоты пропускаемого тока.
§
5.
Ионные
токи
возбудимых
мембран
Современные представления о механизмах транспорта ионов через
мембраны возбудимых клеток сложились главным образом на осно-
ве данных, полученных
методом
фиксации
напряжения.
Сущность
метода состоит в измерении токов, протекающих через клеточную
мембрану в условиях, когда мембранный потенциал автоматически
с помощью усилителя обратной связи поддерживается на любом
заданном уровне.
В покоящейся клетке ток через мембрану не
течет,
так как сум-
ма токов, переносимых ионами разных видов, равна нулю. При сме-
165
щении
потенциала от уровня потенциала покоя возникает трансмем-
бранный
ток, который состоит из емкостного и ионного токов:
где /
м
— суммарный ток;
C
M
dV/dt
— ток зарядки мембранной
емкости; / — ионный ток. Типичный эксперимент по фиксации
напряжения
состоит в том, что разность потенциалов
между
внут-
ренней
и наружной средой клетки изменяют скачком до нового зна-
чения
и, поддерживая потенциал на заданном уровне, регистриру-
ют динамику электрического тока, текущего через мембрану. В мо-
мент ступенчатого переключения потенциала происходит быстрая
перезарядка С
м
; при этом регистрируется волна емкостного тока.
За
доли или сотые доли миллисекунды устанавливается новое зна-
чение мембранного потенциала, емкостный ток прекращается
(так
как
dV/dt
= 0) и измеряется только ток, обусловленный дви-
жением ионов через мембрану.
Компоненты
ионного
тока.
В классических исследованиях
А. Ходжкина и А. Хаксли, выполненных на гигантских аксонах
кальмара, установлено, что суммарный ионный ток, текущий через
мембрану после ступенчатой деполяризации, складывается из
трех
компонентов: быстрого Иа
+
-тока, задержанного
К
+
-тока
и
постоянного во времени и небольшого по значению
тока
утечки.
Проводимость утечки не зависит от мембранного потенциала.
Ходжкин и Хаксли объясняли появление тока утечки движением
ионов
через поврежденную область мембраны. Для измерения то-
ка
утечки обычно сдвигают мембранный потенциал в сторону гипер-
поляризации.
В этом
случае
после кратковременной волны емкост-
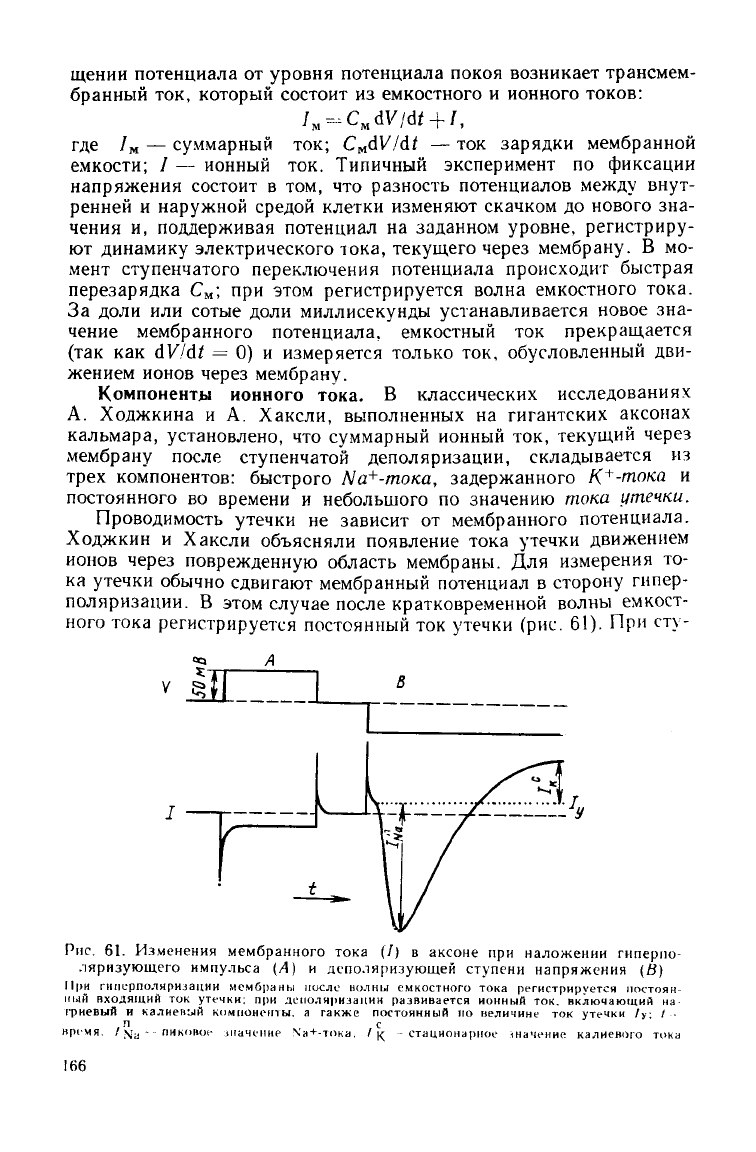
ного тока регистрируется постоянный ток утечки (рис. 61). При сту-
Рпс. 61.
Изменения
мембранного
тока
(7) в
аксоне
при
наложении
гнперпо-
ляризующего
импульса
(А) и
деполяризующей
ступени
напряжения
(й)
11|>и
гиперполиризации
мембраны мосле волны
емкостного
тока
регистрируется
постоян-
ный
входящий ток
утечки;
при деполяризации развивается ионный ток. включающий на
фиевый и калиевый
компоненты,
а также постоянный по величине ток утечки /> / •-
п
с
время.
/ \т
у
-
пиковое
1Наченне \а
+
-тока.
/ (
-стационарное
«наченне калиевого
166

Рис.
62.
Эквивалентная
схе-
ма мембраны аксона:
о
и 1 —
обозначения наружного
раствора
и
аксоплазмы;
С
м
~
емкость мембраны;
l/#Na
H
1.7?
к —
проводимости мембра-
ны
для Na+ и К+. ЧНу — "Р"
водимость утечки;
£м
а
и £^ —
равновесные потенциалы
для
Na+
и К
+
;
£у--потенциал
ре
версии тока утечки; V—напря
жение
на
мембране
пенчатон деполяризации помимо тока
угечки развиваются токи
Na+ и К
+
,
характеризующиеся сложной кинетикой.
Натриевый
и
калиевый токи разви-
ваются независимо, поэтому
с
помощью
различных приемов
удается
разделить
суммарную кривую ионного тока
на
компоненты
и
охарактеризовать
по от-
дельности свойства системы
Na
+
- и
К
+
проводимости.
Так, при
замене
в
среде
Na
+
на
непроникающие катионы
холина Ыа
+
-компонент ионного тока
практически исчезает
и
регистрируется
только К
+
-ток, имеющий выходящее
направление
Суммарный ионный
ток в
аксонах
удается
разделить
на
компоненты
и с
помощью
блокаторов
Ыа
+
-
и
К
+
-каналов.
Натриевый
ток
подавляется тетродотоксином, К
+
-ток исчезает
при
введении внутрь аксона тетраэтиламмония, высоких концентраций
Cs~.
а
также
при
обработке волокон 4-аминопиридином.
С
помощью
блокаторов
Na
+
- и
К
+
-токов
удалось
вычленить
Са
2+
-токи
в
мемб-
ране нейронов
и
подробно охарактеризовать свойства Са
2+
-кана-
лов.
К
блокированию ионных токов прибегают
и для
вычленения
воротных токов, связанных
с
открыванием каналов.
Потенциал
реверсии ионного тока. В
опытах
на
аксонах каль-
мара установлено,
что
потенциал,
при
котором начальный ионный
гок
изменяет свое направление, близок
по
величине
к
равновесному
потенциалу для№
+
. рассчитываемому
по
формуле Нернста
(IV.1.3).
Этот
результат,
легко объяснимый
на
основании принятой эквива-
лентной схемы мембраны
(рис. 62),
показывает,
что в том
случае,
когда мембранный потенциал равен равновесному потенциалу
Na+(£
Na
),
движущая сила
для
Na+ равна нулю. Поэтому действую-
щую силу, определяющую направление движения ионов через
мем-
брану, можно выразить
в
виде разности
между
мембранным
по-
тенциалом
V и
равновесным потенциалом
для
данного иона
£;.
Для Na+ действующая сила равна
V — £
Na
.
Ионные
проводимости возбудимой мембраны. Так как
после
ступенчатого смещения мембранного потенциала
на
заданный
уро-
вень действующая сила постоянна, очевидно,
что
изменения тока
Na+
обусловлены изменением проводимости мембраны
для Na
+
:
fsa
=
gNa
(
V —
£ыа).
Таким
образом, если суммарная кривая мембранного тока разделе-
на
на
компоненты,
то
можно проследить изменения проводимости
мембраны
для Na+ и К
+
во
времени:
gNa
=
/
N
a/(V
—
£
Na
);
дк =
167
Сумма парциальных ионных проводимостей
g
t
равна общей
про-
водимости мембраны.
Проводимость
для
иона
i,
определяемую соотношением
g,- =
=
1\!{V — £;),
называют
хордовой проводимостью. Ее следует от-
личать
от дифференциальной проводимости G;,
определяемой
по уг-
лу наклона вольт-амперной характеристики:
G
;
= d/j/dV. Их
различие можно рассмотреть
на
примере вольт-амперной кривой
для пикового
Na
+
-
тока.
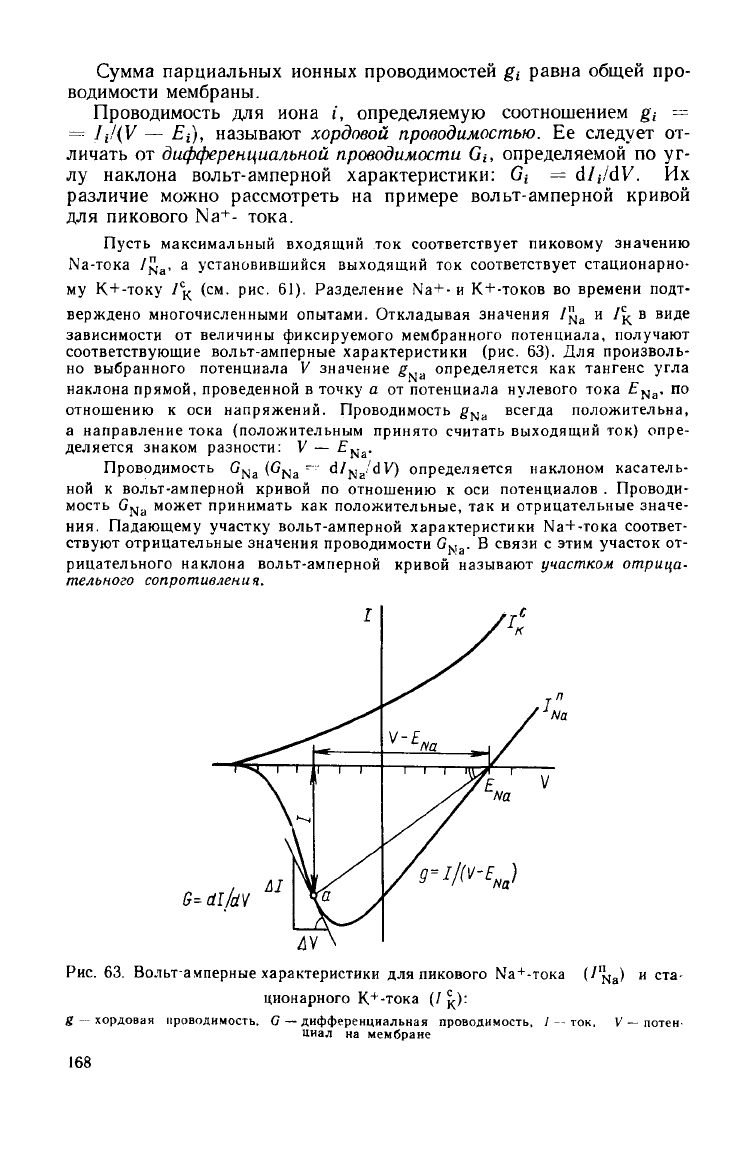
Пусть максимальный входящий
ток
соответствует
пиковому значению
Na-тока
1^
а
, а
установившийся выходящий
ток
соответствует
стационарно-
му К
+
-току
/
с
к
(см. рис. 61).
Разделение Na+-
и
К
+
-токов
во
времени подт-
верждено многочисленными опытами. Откладывая значения
У^
а
и /^ в
виде
зависимости
от
величины фиксируемого мембранного потенциала, получают
соответствующие вольт-амперные характеристики
(рис.
63). Для
произволь-
но
выбранного потенциала
V
значение
g
определяется
как
тангенс
угла
наклона
прямой, проведенной
в
точку
а от
потенциала нулевого тока
£
Na
. по
отношению
к оси
напряжений. Проводимость
g
Na
всегда
положительна,
а направление тока (положительным принято считать выходящий
ток)
опре-
деляется знаком разности:
V —
£[yj
a
-
Проводимость
G
Na
(G
Na
-
dJ^JdV)
определяется наклоном касатель-
ной
к
вольт-амперной кривой
по
отношению
к оси
потенциалов
.
Проводи-
мость
G[y|
a
может принимать
как
положительные,
так и
отрицательные значе-
ния.
Падающему
участку
вольт-амперной характеристики Ыа+-тока соответ-
ствуют
отрицательные значения проводимости G
Na
.
В
связи
с
этим участок
от-
рицательного наклона вольт-амперной кривой называют
участком
отрица-
тельного
сопротивления.
Рис.
63.
Вольт-амперные характеристики
для
пикового Na
+
-TOKa
(/^
а
) и ста-
ционарного
К
+
-тока
(/^)
:
g
-
хордовая
проводимость.
С —
дифференциальная проводимость, /--ток.
V —
потен-
циал
на
мембране
168

Кинетика ионных токов.
Значения проводимостей
g^
a
и g
K
изменяются
во
времени
и
зависят
от
потенциала.
При
ступенчатой
деполяризации мембраны
g
Na
быстро возрастает
до
максимума,
а
затем снижается
до
низкого постоянного уровня.
В
отличие
от
этого
£к
возрастает
от
низкого уровня
до
стационарного максималь-
ного уровня. Хотя нарастание
g
Na
происходит гораздо быстрее,
чем
нарастание g&,
оба
процесса развиваются после некоторого периода
задержки.
В мембране
на
единичной площади поверхности содержится
ог-
раниченное число проводящих элементов. Поэтому, если
все
прово-
дящие каналы открыты, достигается максимальный уровень прово-
димости, обозначаемый
g
Na
и g
K
для
ионов
Na и К.
Промежуточ-
ные состояния проводимости,
при
которых открыта лишь часть
про-
водящих элементов, удобно представить
в
виде произведения макси-
мальной проводимости,
на
безразмерные параметры, варьирующие
в
пределах
от
нуля
до
единицы. Параметры, определяющие уровень
Na+-
проводимости, получили название
т и h;
уровень К
+
-проводи-
мости определяется параметром
п. При
ступенчатом смещении
мем-
бранного потенциала каждый
из
этих параметров совершает относи-
тельно медленный
переход
к
новому равновесному значению.
Изменение
т, п и h
можно формально рассматривать
как
следст-
вие перераспределения некоторых заряженных внутримембранных
частиц
между
внутренней
и
наружной границами мембраны под дей-
ствием электрического поля. Используя
такую
интерпретацию,
зна-
чения
тип
рассматривают
как
вероятность нахождения положитель-
но
заряженных
т- и
л-частиц
у
внешней поверхности мембраны.
При
больших отрицательных потенциалах
все
положительно заря-
женные подвижные частицы сосредоточены
у
внутренней поверхно-
сти мембраны; этому состоянию
соответствуют
значения
тип, рав-
ные нулю,
и
значение
h,
равное единице.
При
ступенчатой деполя-
ризации
клетки управляющие частицы начинают переходить
к
внеш-
ней
поверхности, причем m-частицы перемещаются
на
порядок
бы-
стрее,
чем п-и
/г-частицы. Такой
переход
соответствует
увеличе-
нию
параметром
т и п и
уменьшению параметра
h.
Увеличение
т
после ступенчатой деполяризации описывает фазу
нарастания
g
Na
, т. е.
активацию Ыа+-проводимости; медленное
уменьшение
h
описывает фазу спада
g
Na
, т. е.
инактивацию
Na+-
проводимости. Увеличение
п
отвечает активации /С
+
-проводимости.
В модели Ходжкина
и
Хаксли мембранный
ток
изображают
в
виде суммы емкостного тока, Ыа+-тока,
К
+
-
тока
и
тока утечки:
'м
= С
м
— +
g~
Na
m
3
h (V — Es.)
+gKn*
(V — Ек) +"£
у
(V-£
y
),
где
т, п и h
описываются однотипными уравнениями
dm/dt
^a
m
(l —
т)~$
т
т.
169
Л
1
1
-/о
1
1 1
-50
-20
20
60 V.MS
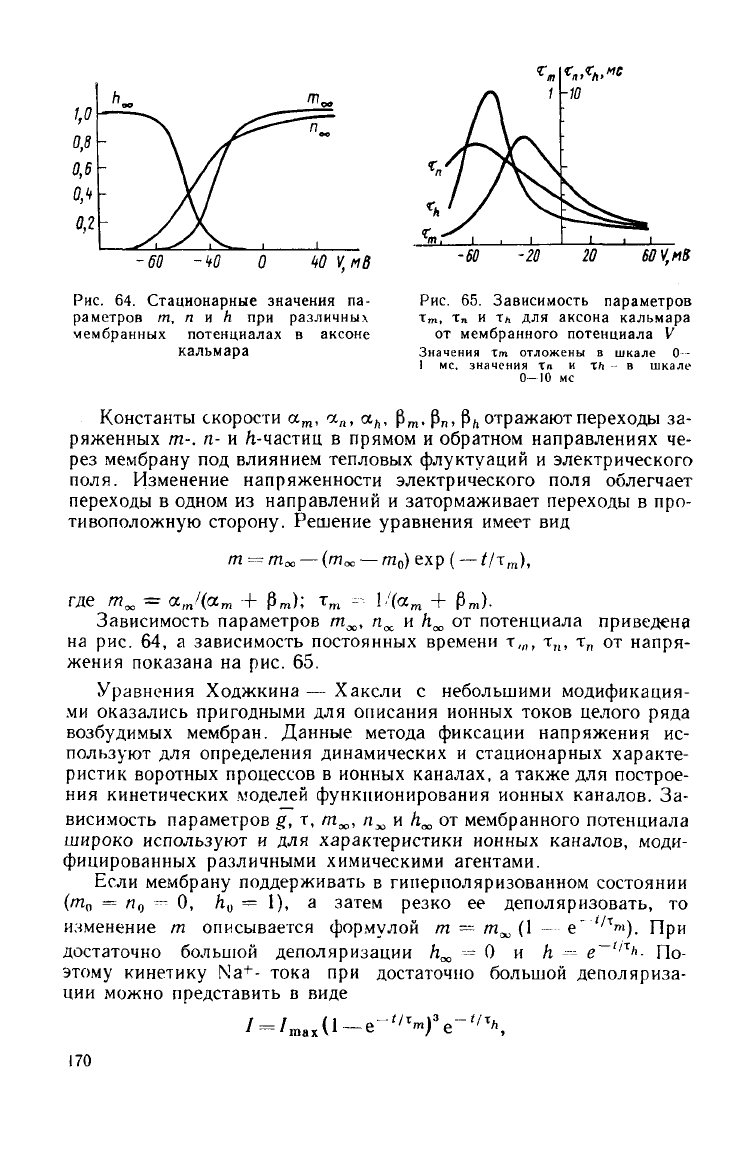
Рис.
64.
Стационарные значения
па-
раметров
m, n и h при
различных
мембранных потенциалах
в
аксоне
кальмара
Рис.
65.
Зависимость параметров
т»,
х
п
и тл для
аксона кальмара
от мембранного потенциала
V
Значения
Тт отложены в шкале О-
1
мс. значения Хп и xh - в шкале
О—10 мс
Константы скорости а
т
, а.
п
, а
Л
, р
т
. Р„, P
h
отражают переходы за-
ряженных т-. п- и дг-частиц в прямом и обратном направлениях че-
рез мембрану под влиянием тепловых флуктуации и электрического
поля.
Изменение напряженности электрического поля
облегчает
переходы в одном из направлений и затормаживает переходы в про-
тивоположную сторону. Решение уравнения имеет вид
т
=
rrioo
—
(moo
— т
0
) ехр (—
t/x
m
),
где т^ -
aj(a
m
+ p
m
); т
т
^ \>(а
т
+ р
т
).
Зависимость параметров т^, п
х
и h^ от потенциала приведена
на
рис. 64, а зависимость постоянных времени т,„, т„, т
л
от напря-
жения
показана на рис. 65.
Уравнения Ходжкина — Хаксли с небольшими модификация-
ми
оказались пригодными для описания ионных токов целого ряда
возбудимых мембран. Данные метода фиксации напряжения ис-
пользуют для определения динамических и стационарных характе-
ристик
воротных процессов в ионных каналах, а также для построе-
ния
кинетических моделей функционирования ионных каналов. За-
висимость параметров g, т, т^, п
х
и h
x
от мембранного потенциала
широко
используют и для характеристики ионных каналов, моди-
фицированных различными химическими агентами.
Если мембрану поддерживать в гиперполяризованном состоянии
(т„ — л
0
~ 0, h
0
= 1), а затем резко ее деполяризовать, то
изменение т описывается формулой т — т^{\ — -• е"
'
/т
"
!
).
При
достаточно большой деполяризации h
x
- 0 и h --
e~''
x
i>-
По-
этому кинетику Na
+
- тока при достаточно большой деполяриза-
ции
можно представить в виде
170