Хаусман К., Хюльсман H., Радек Р. Протистология
Подождите немного. Документ загружается.


108 Эволюция и таксономия
Некоторые лишенные фрустулы
диатомовые обитают как облигатные
эндосимбионты в морских протистах
(например, в крупных фораминиферах)
и в тканях многоклеточных животных.
Их и динофлагеллат рода Symbiodinium
называют зооксантеллами. Зооксантел-
лы играют важную роль в карбонатной
биоминерализации при строительстве
известковых раковин.
Диатомовых традиционно подразде-
ляют на два порядка: Centrales и Реп-
nales. Однако известно, что Centrales,
скорее всего, не является монофилети-
ческим таксоном, поэтому в дальней-
шем будем использовать только нефор-
мальные названия: центрические диато-
мовые и пеннатные диатомовые.
Центрические диатомовые
Створки центрических диатомовых
имеют радиальную симметрию. Боль-
шинство — не обладающие подвижно-
стью одиночные или колониальные
планктонные формы. Половой процесс
в форме оогамии. Сперматозоид несет
единственный жгутик аббератного стро-
ения: отсутствуют центральная пара
микротрубочек и переходная спираль,
кинетосома состоит из девяти дуплетов
вместо триплетов (рис. 70).
Примеры: Biddulphia, Coscinodiscus,
Odontella, Rhizosolenia.
Пеннатные диатомовые
Пеннатные диатомовые имеют вы-
тянутый в плоскости шва билатерально
симметричный панцирь
1
. Преимуще-
ственно бентосные формы, но некото-
рые колониальные виды обитают и в
толще воды. Жгутиковые стадии неиз-
вестны. Половой процесс изогамный,
гаметы амебоидные, всегда с пластида-
ми. Эпивальва и гиповальва часто несут
продольную щель («шов»), которая иг-
рает определенную роль в локомо-
ции — быстром скольжении по субстра-
ту. Биомеханическая основа этой фор-
мы подвижности еще неизвестна; клю-
чевую роль, по-видимому, играет
выделение через шов полисахаридов.
Примеры: Bacillaria, Navicula, Nit-
zschia, Pleurosigma, Pseudo-nitzschia.
Класс HETEROMONADEA Leedale,
1983 (Xanthophyceae) —
ЖЕЛТО-ЗЕЛЕНЫЕ ВОДОРОСЛИ,
КСАНТОФИТЫ
Жгутиковые клетки ксантофитовых
очень похожи на хризомонад по ультра-
структурным признакам и гетероконтно-
му расположению жгутиков. Похожий
на пенек укороченный рулевой жгутик
совершенно гладкий и оканчивается
терминальной нитью, акронемой. Как и
у хризомонад, этот жгутик имеет базаль-
ное утолщение напротив расположенно-
го в хлоропласте глазка. Существенное
отличие от хризомонад — полное отсут-
ствие фукоксантина, что в сочетании с
наличием (3-каротина и ксантофиллов
дает более желто-зеленую окраску кле-
ток. Кроме этого, они отличаются и по
строению эндогенных цист. Желто-зеле-
ные водоросли с нитчатыми талломами
(Tribonema, Vaucheria) хорошо извест-
1
Авторы пособия не различают билатеральную симметрию и двулучевую симметрию. В данном
случае следует говорить о двулучевой симметрии. Билатеральная симметрия присуща очень немно-
гим протистам. Таковы, например, дипломонады и некоторые из высших феодарий. — Прим. ред.

Система протистов 109
Рис. 72. Heterokonta, Eustigmatophyceae: схема строения зооспоры эустигматофита на про-
дольном срезе, вз — вздутие в основании жгутика, мае — мастигонемы, ми — митохонд-
рия, хл — хлоропласт, ст — стигма, я — ядро (по Нибберду). Увел.: 4 400х.
ны; имеется также несколько жгутико-
вых видов. В настоящее время наибо-
лее интенсивно изучаются амебоидные
формы. Ксантофитовые имеют невысо-
кую численность и ограниченное рас-
пространение; их экологическое значе-
ние, по-видимому, невелико.
Примеры: жгутиконосцы: Chloro-
meson; амебоидные: Chlamydomyxa,
Rhizochloris, Reticulosphaera.
Класс EUSTIGMATOPHYCEAE
Hibberd & Leedale, 1971 —
ЭУСТИ ГМ АТОФИТОВЫ E
Эустигматофитовые представляют
собой небольшой таксон, включающий
6 родов и около 15 видов. Все являют-
ся фототрофными коккоидными форма-
ми. Некоторые виды образуют зооспо-
ры (10-15 мкм длиной) с одним или
(редко. — Прим. пер.) двумя неравны-
ми жгутиками (рис. 72). Уникальная
особенность жгутикового аппарата зак-
лючается в том, что вдоль основания
переднего жгутика идет Т-образное
вздутие, которое формирует тесный
контакт с передним концом зооспоры
(рис. 72), где расположен очень круп-
ный лежащий вне хлоропласта глазок
(стигма)
1
. Единственный пластидный
компартмент окружен тремя мембрана-
ми и содержит продольные ламеллы из
трех неплотно прилегающих друг к дру-
гу тилакоидов. Наличие трехчленных
мастигонем и спирали в переходной
зоне жгутика указывает на близкое род-
ство с типичными гетероконтами.
Примеры: Eustigmatos, Chlorobotrys,
Monodopsis.
Класс LABYRINTHULEA Cienkowski,
1867 (Labyrinthulomycota) —
ЛАБИРИНТУЛЫ
Этот таксон содержит лишь один
отряд Labyrinthulida с двумя семейства-
ми Labyrinthulidae и Thraustochytriidae,
в которых насчитывается около 40 ви-
дов. Эти организмы встречаются пре-
имущественно в морских прибрежных
и эстуарных местообитаниях, реже в
пресных водах или в засоленных по-
чвах. Их можно обнаружить на водо-
рослях и остатках сосудистых растений
или в органическом детрите. Считает-
В отличие от хризомонад, жгутиковое вздутие фоторецепторного аппарата находится на дви-
гательном, а не на рулевом жгутике. — Прим. пер.
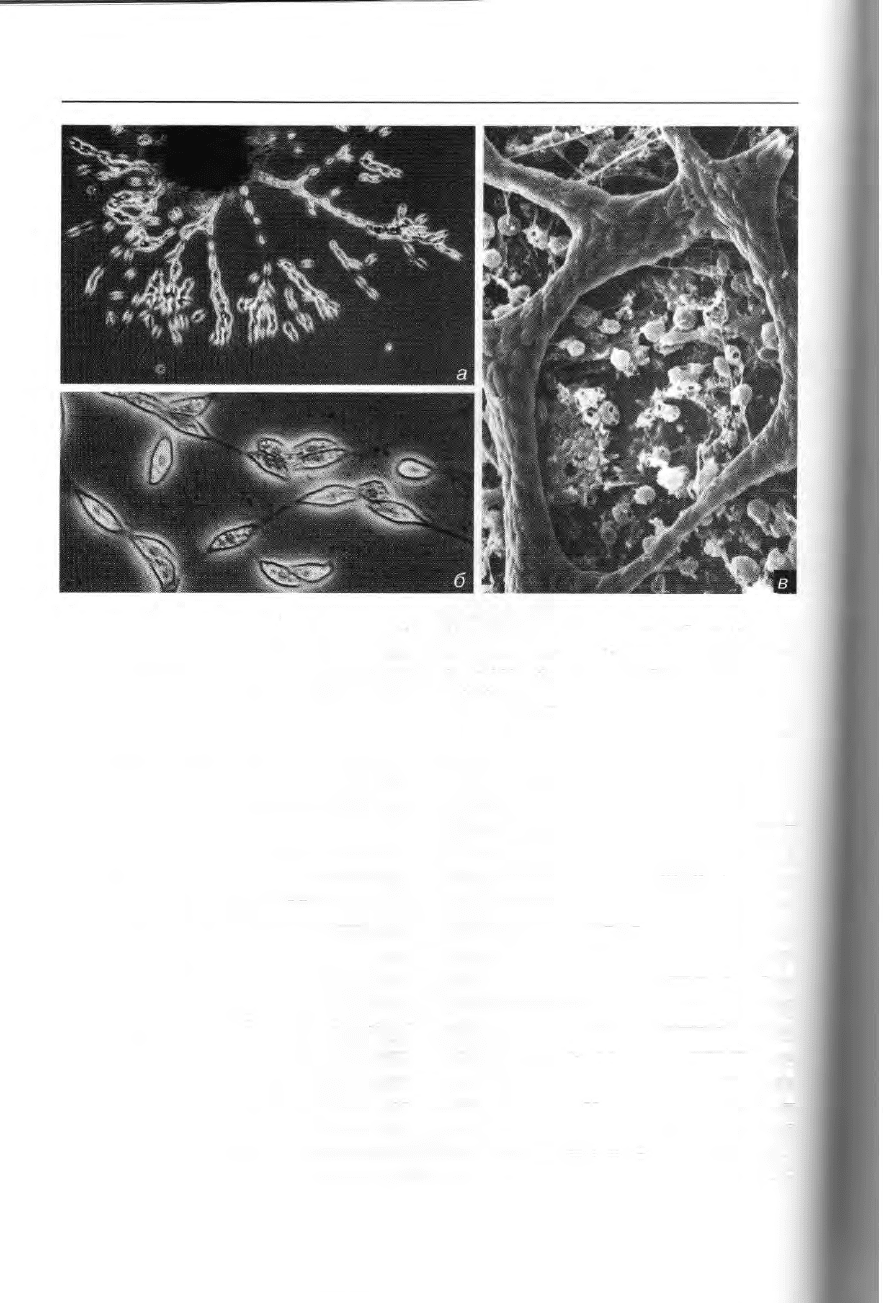
110 Эволюция и таксономия
Рис. 73. Heterokonta, Labyrinthulea: а — плазмодиальная сеть Labyhnthula coenocystis; б —
веретеновидные ядросодержащие тела, мигрирующие внутри округлых в сечении нитей
филоподиальной сети; в — плотно упакованные ядросодержащие тела внутри цилиндри-
ческих филоподий — сканирующая электронная микроскопия (а и б — из: Stey: Zellforsch.
Mikrosk. Anat. 102 [1969] 387). Увел.: a — 190x, б— 800x, s — 330x.
ся, что два вида: Labyrinthula zosterae и
L. macrocystis — вызывают карлико-
вость нескольких видов морских трав
из семейства зостеровых. Эти и некото-
рые другие представители рода Laby-
rinthula способны пронизывать клеточ-
ную стенку у наземных растений, гри-
бов, диатомовых и других организмов.
На стадии трофонта лабиринтулы
представляют собой блуждающие плаз-
модии (рис. 73). Веретеновидные ядро-
содержащие тела («клетки») постоян-
ной формы или меняющие свои очерта-
ния каким-то образом скользят внутри
составляющих сеть тяжей, которые на-
поминают псевдоподии. Совокупность
тяжей называется эктоплазматической
сетью. Покровом эктоплазматической
сети служит плазмалемма. Каждая вере-
теновидная «клетка» одета уплощенной
цистерной, которая на электронограм-
мах читается как оболочка из двух па-
раллельных мембран с небольшим про-
светом между ними. В просвете распо-
лагаются чешуйки. Цистерна, охватыва-
ющая «клетку», не сплошная: в одной
или нескольких точках внешняя и внут-
ренняя ее мембраны соединяются, фор-
мируя пору. Через эту пору цитоплазма
«клетки» сообщается с цитоплазмой
сети (рис. 74). На ультраструктурном
уровне пора весьма сложно организова-
на. Она получила название ботросома,
или сагеногенетосома, сагеноген. Бот-
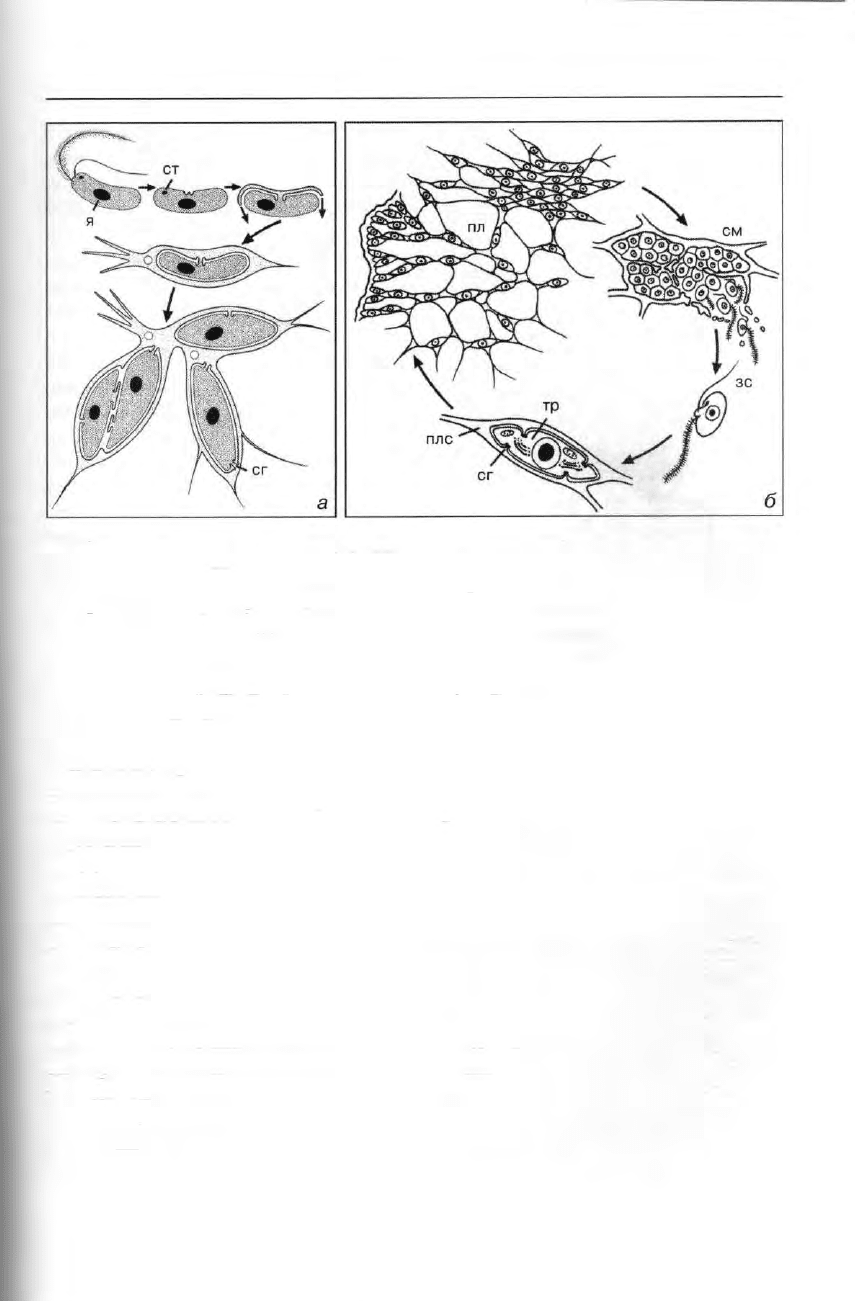
Система протистов 111
Рис. 74. Labyrinthulea: а — модель развития эктоплазматической сети у лабиринтулид. ст —
глазок, я — ядро, сг — сагеногенетосома (= ботросома). б — жизненный цикл Labyrinthula:
плазмодии (пл), содержащие веретеновидные «клетки»-трофонты, формируютспорогенные
массы (см), в которых клетки претерпевают мейоз; выходящие зооспоры (зс), по-видимому,
развиваются в новые клетки-трофонты (тр), плс — плазмодиальная сеть (по Портеру).
росомы и та оболочка из двух мембран,
которую они принизывают, в совокуп-
ности представляют собой апоморфный
признак лабиринтул. Следует вновь
подчеркнуть, что «клетки» не являются
истинными клетками. Каждый ядросо-
держащий участок цитоплазмы лишь
частично отграничен от обобщенной
сети цитоплазматических тяжей. Саму
же сеть вполне допустимо рассматри-
вать как своеобразную псевдоподиаль-
ную конструкцию.
Механизм движения неизвестен.
Ультраструктурные исследования по-
зволяют предположить, что задейство-
ваны расположенные в тяжах сети ак-
тиновые микрофиламенты.
Размножение идет за счет бинарно-
го деления «клеток» внутри сети или
путем плазмотомии плазмодия. Не до
конца понятный жизненный цикл не-
сомненно включает половой процесс;
у наиболее изученных видов отмечены
многоклеточные цисты, спорангии (со-
русы), а также и двужгутиковые зоо-
споры (рис. 74). Для понимания род-
ственных связей лабиринтул особенно
важны признаки строения зооспор.
Последние представлены гетероконт-
ными клетками с трехчленными масти-
гонемами на переднем жгутике. Эти
морфологические черты зооспоры вы-
дают принадлежность лабиринтул к
Heterokonta. Имеющееся у зооспор пиг-
ментное пятно, подстилающее рулевой
жгутик, — тоже признак, характерный
для способных к фотосинтезу жгутико-
носцев из состава гетероконтов.
Примеры: Labyrinthula, Labyrithu-
loides, Thraustochytrium.

112 Эволюция и таксономия
Рис. 75. Heterokonta, Raphidomonadea: схе-
ма строения клетки Chattonella sp. с харак-
терными кольцевыми диктиосомами (д) у
поверхности ядра (я), к — корешок, мц —
мукоцисты, пл — пластиды (по Хейвуду).
Увел.: 3 700х.
Класс RAPHIDOMONADEA
(Chloromonadea) Heywood &
Leedale, 1983 — РАФИДОМОНАДЫ
Рафидомонады (около 20 видов) —
исключительно монадные, обычно дор-
совентрально уплощенные организмы
(рис. 75). Их клетки, лишенные чешуек
и клеточной стенки, достигают нети-
пично большого для фитофлагеллат
размера, до 90 мкм. Форма клетки от-
носительно постоянна благодаря нали-
чию упругой пелликулы. Мукоцисты и
несколько вариантов веретеновидных
трихоцист с невыясненной функцией
располагаются в поверхностной зоне
цитоплазмы. Гетероконтные жгутики
выходят из вентрального углубления на
переднем полюсе клетки; их кинетосо-
мы связаны корешковыми структурами
с поверхностью ядра. Длинный направ-
ленный назад и преимущественно пас-
сивный гладкий жгутик не имеет ба-
зального вздутия. Более короткий и
очень быстро вращающийся передний
жгутик несет трубчатые мастигонемы,
которые синтезируются в цистернах эн-
доплазматического ретикулума или
диктиосомах. Стигма отсутствует. К пе-
редней поверхности крупного ядра
прилегает слой диктиосом, что являет-
ся характерной чертой рафидофитовых.
Ряд крупных везикул, производных ап-
парата Гольджи, постоянно трансфор-
мируется в сократительные вакуоли, по
крайней мере, у пресноводного рода
Vacuolaria.
Рафидомонады — фототрофы, но
возможно имеют филогенетические свя-
зи с гетеротрофными тауматомонадами
(из Cercozoa). Многочисленные пласти-
ды ярко-зеленого цвета с хлорофилла-
ми а и с, а также с дополнительными
пигментами из группы каротиноидов —
ксантофиллами и фукоксантинами. Хло-
ропласта содержат стопки из трех ти-
лакоидов и опоясывающую ламеллу.
Питательные вещества представлены
липидами и запасаются в виде капель.
Массовое развитие Heterosigma ака-
shiwo, Chattonella antiqua, Ch. marina,
Fibrocapsa japonica в рыбоводных хо-
зяйствах и естественных биотопах мо-
жет приводить к гибели рыбы в резуль-
тате воздействия выделяемых водорос-
лями полиэфирных токсинов, совокуп-
но называемых бревитоксин.
Система протистов 113
Примеры: Chattonella, Heterosigma,
Gonyostomum, Fibrocapsa, Vacuolaria.
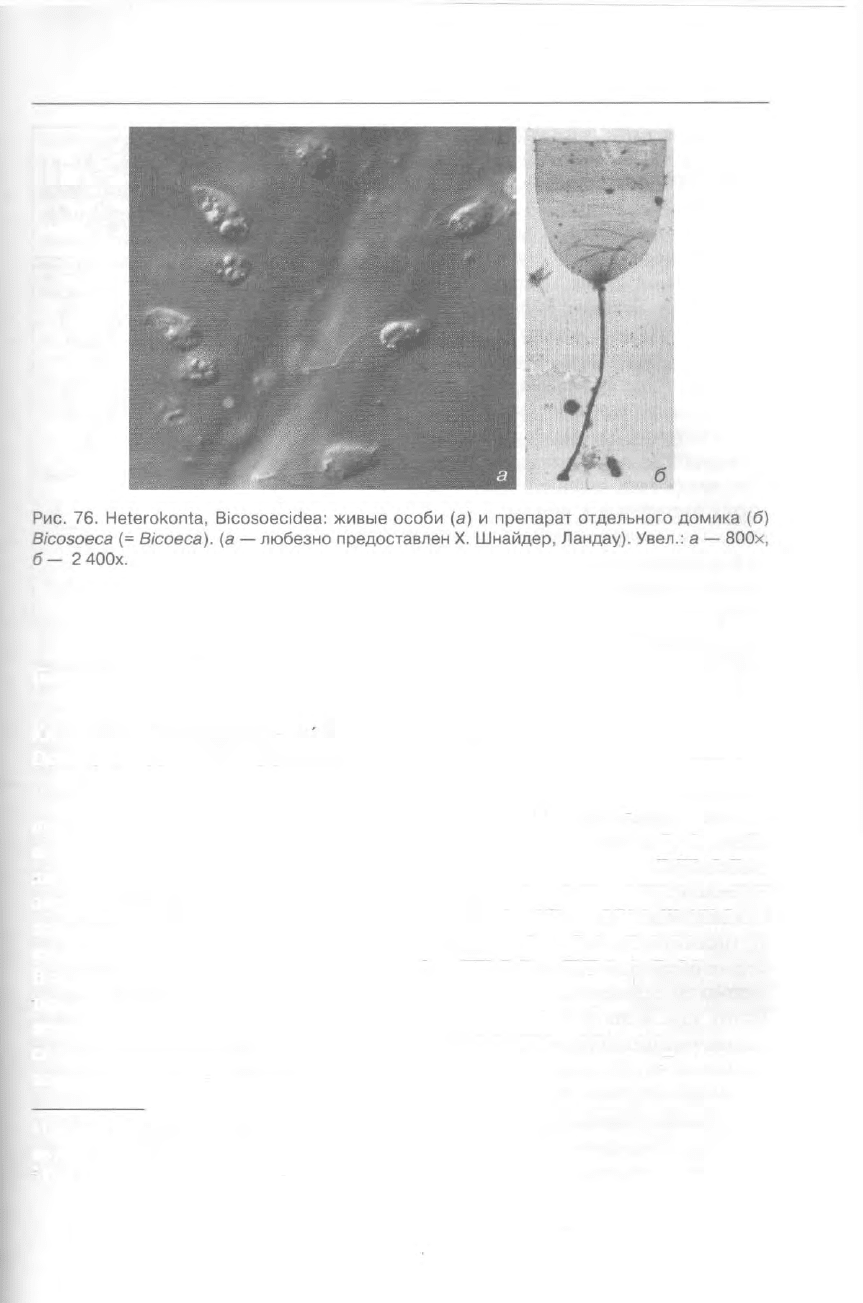
Класс BICOSOECIDEA Grasse &
Deflandre, 1952 — БИКОЗОЕЦИДЫ
Около 40 видов этого таксона
представлены одиночными клетками
или колониями и характеризуются на-
личием бокаловидной раковинки, или
лорики, построенной из хитина
1
(в
противоположность силикатным доми-
кам хоанофлагеллат и хризомонад).
Все бикозоэциды — одноядерные ге-
тероконтные жгутиконосцы. Гладкий
жгутик идет назад в ложбинке тела и
служит для прикрепления клетки к ос-
нованию домика. Этот жгутик спосо-
бен сокращаться и втягивать клетку
внутрь лорики. Второй, более длин-
ный жгутик направлен вперед. Он не-
сет один или два ряда трубчатых мас-
тигонем и служит для собирания час-
тиц пищи, в первую очередь бактерий
(рис. 76, 273е). Размер клеток около
5 мкм. Они ведут свободно-подвиж-
ный или сидячий образ жизни, в пос-
леднем случае прикрепляясь лорикой
к субстрату. Бикозоециды входят в со-
став гетеротрофного нанопланктона
морских и пресных вод
2
и, очевидно,
представляют собой одну из ключевых
в трофическом отношении групп мик-
роскопических организмов.
Примеры: Bicosoeca (= Bicoeca),
Cafeteria, Pseudobodo.
1
По последним данным, эта группа включает около 60 видов, отнесенных к 12 родам. Многие
не имеют раковинки. — Прим. пер.
:
Известны и почвенные бикозоециды. — Прим. пер.
114 Эволюция и таксономия
Класс HYPHOCHYTRIOMYCETES
Patterson, 1982 —
ГИФОХИТРИДИЕВЫЕ
Гифохитридиевые — небольшой и
не слишком известный таксон, вклю-
чающий на сегодняшний день пример-
но 25 видов. Название группы — про-
изводное от имени одного из пяти ее
родов. Это сапрофиты, некрофаги или
паразиты, которые обильно развивают-
ся в морских, пресноводных и назем-
ных биотопах на самых различных
органических субстратах: ракообраз-
ных, водорослях, оомицетах, разлагаю-
щихся растительных остатках, мелких
яйцах беспозвоночных-, пыльце и т.д.
Гифохитридиевые, по-видимому, не
способны контролировать численность
популяции хозяина и не являются се-
рьезными вредителями. Таким обра-
зом, их экологическое значение неве-
лико.
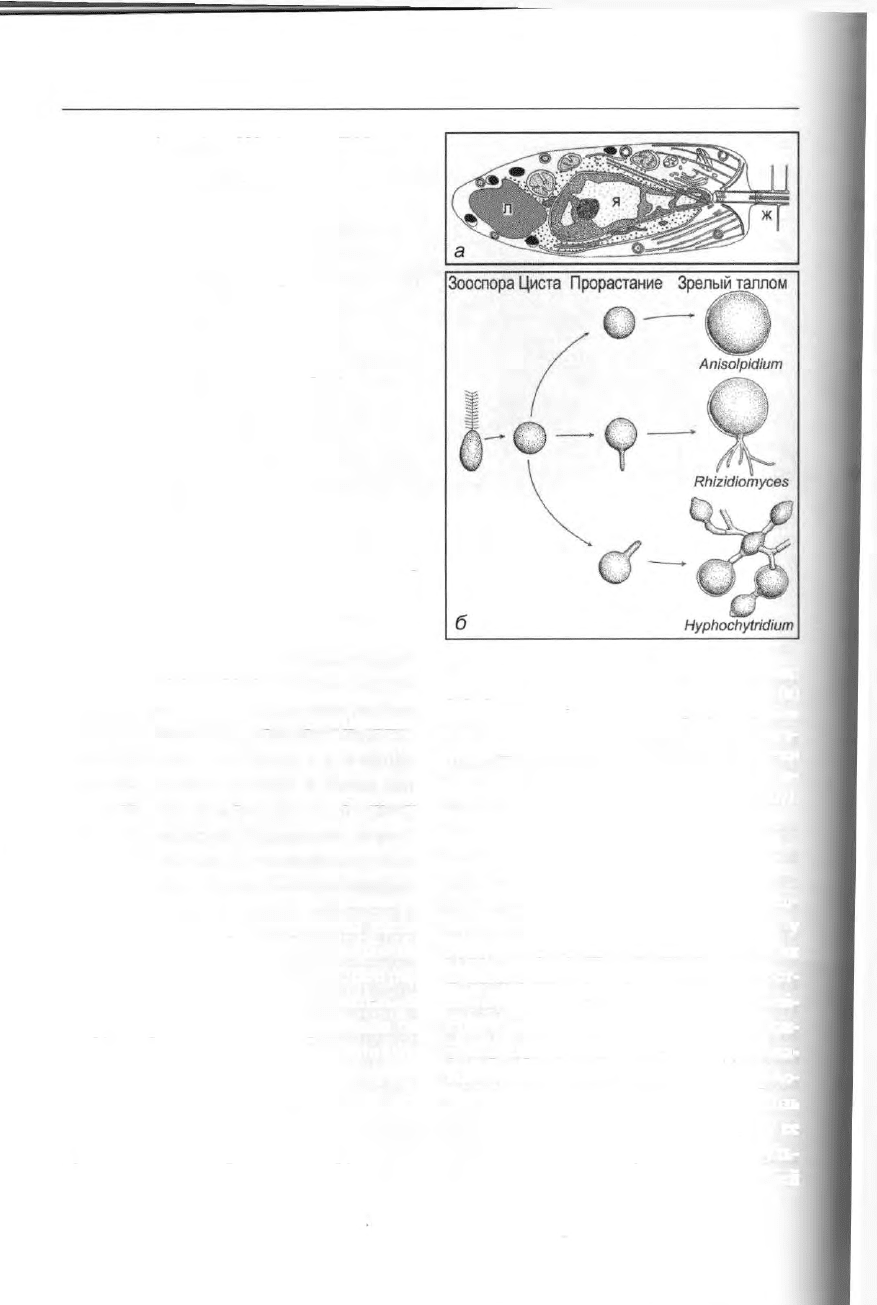
Жизненный цикл включает подвиж-
ные зооспоры и неподвижные много-
ядерные талломы. Зооспоры длиной
4-6 мкм (рис. 77а) несут единственный
жгутик, занимающий апикальное поло-
жение; второй жгутик редуцирован до
кинетосомы. Как и в других классах
Heterokonta, жгутик несет мастигонемы.
Отличительная черта устройства клет-
ки гифохитридиевых — это неравно-
мерное распределение рибосом. Все ри-
босомы сосредоточены исключительно
вокруг ядра, и эта зона отграничена от
остальной цитоплазмы цистернами
ЭПР.
После оседания в подходящем мес-
те в тканях хозяина или на мертвой
органике, зооспора инцистируется и
затем трансформируются в проросток.
Рис. 77. Heterokonta, Hyphochytriomycetes:
схема строения зооспоры Hyphochytrium (а)
и схема стадий жизненного цикла с тремя
различными типами таллома (б), ж — жгу-
тик, л — липидные капли, я — ядро. Точки
вокруг ядра в цитоплазме обозначают ри-
босомы (а — по Уэллсу, б — по Фулеру).
Увел.: а — 5 ОООх.
Он окружен клеточной стенкой из цел-
люлозы и хитина, по крайней мере, у
наиболее изученного Rhizidiomyces. Как
показано на рисунке 776, эти пророст-
ки формируют специфичные для семей-
ства трубки, а позднее систему ризои-
дов. Ризоиды часто несут продолгова-
тые вздутия, в частности, у рода Hypho-
chytrium. Ризоиды прорастают сквозь
стенку клетки хозяина и поглощают ее
содержимое. В экспериментах по куль-
тивированию выделение следующей
Система протистов 115
генерации зооспор начинается через
18-48 часов. Зооспорогенез идет по-
средством отделения многоядерных
фрагментов зрелого таллома, которые
выходят из общей материнской оболоч-
ки. Половой процесс и формирование
цист покоя также имеют место.
Примеры: Anisolpidium, Hyphochyt-
rium, Rhizidiomyces.
Класс OOMYCETES Dick, 1976 —
ООМИЦЕТЫ, ВОДНЫЕ ПЛЕСЕНИ
Оомицеты насчитывают в своем
составе около 800 видов и 65 родов и
представляют собой одну из основных
групп гетеротрофных гетероконтов.
Они распространены всесветно и
встречаются практически во всех вод-
ных и наземных экосистемах. Если
упомянутые выше гифохитридиевые
безвредны, то продвижение оомицетов
зачастую опустошительно. В частно-
сти, к оомицетам относятся: Phyto-
phthora infestans — возбудитель фито-
фтороза картофеля, вызвавший Вели-
кий голод в Ирландии, также называе-
мый Ирландский картофельный голод
(The Great Irish Potato Famine), в
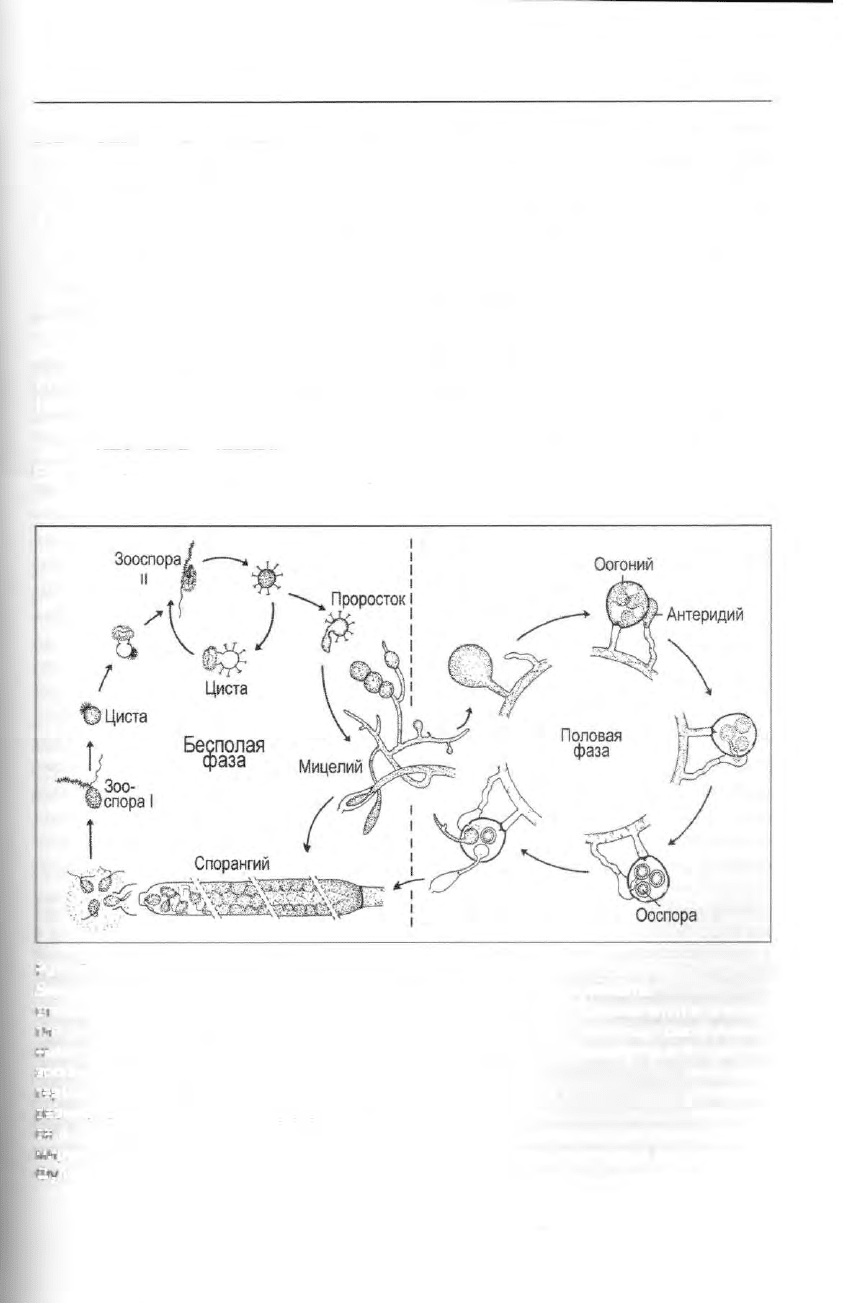
Рис. 78. Heterokonta, Oomycetes: бесполые и половые стадии в жизненном цикле
Saprolegnia; заметим, что другие роды Oomycetes существенно отличаются по особенно-
стям жизненного цикла. При бесполом размножении деление цитоплазмы внутри споран-
гия приводит к образованию зооспор. Первичные зооспоры (I) с апикально расположен-
ными жгутиками превращаются в цисты, которые затем дают инфекционные вторичные
зооспоры (II) с боковыми жгутиками. Они могут снова инцистироваться или формировать
-ерминативную трубку (проросток), которая развивается в гифы (мицелий). При половом
размножении на гифах образуются многоядерные антеридии и оогонии; они развивают-
ся в контакте друг с другом, и в них синхронно идет мейоз. После оплодотворения фор-
мируются толстостенные устойчивые ооспоры, которые затем прорастают в мицелий (по
Фулеру).

116 Эволюция и таксономия
1840-е годы
1
; Pythium spp., основной
патоген рассады многих культурных
растений; Plasmopara viticola — па-
разит винограда; Saprolegnia spp. и
Achlya spp., которые поражают лососе-
вых рыб в садковых хозяйствах. По-
мимо большой группы облигатных и в
высшей степени патогенных парази-
тов, в том числе перечисленных выше,
среди оомицетов есть и сапрофиты
(см. рис. 238).
Жизненный цикл (описанный здесь
для Saprolegnia) включает бесполые
стадии: подвижные зооспоры и непод-
вижные несептированные гифы с вер-
хушечным ростом, на которых форми-
руются зооспорангии; а также половые
стадии с образованием гаметангиев:
женских оогониев и мужских антери-
диев (рис. 78). Как зооспорангии, так и
гаметангий отделяется от ценотической
гифы поперечной стенкой (септой).
Мейоз идет при формировании гамет;
все другие стадии жизненного цикла
диплоидны. Первичные зооспоры —
клетки яйцевидной формы с двумя апи-
кальными жгутиками — высвобожда-
ются группами из зооспорангиев и бы-
стро инцистируются. После эксцисти-
рования они претерпевают серию цик-
лов репродукции, которые включают
стадию бобовидных вторичных зоо-
спор с теперь уже вентрально выходя-
щими жгутиками и стадию шиповатых
цист
2
. В ходе созревания цисты форми-
руется зародышевая трубка, которая за-
тем прорастает сквозь покровы хозяи-
на или в разлагающуюся органику и
развивается в так называемый споран-
гиофор — мицелий с ветвящимися не-
септированными гифами и апикальны-
ми зооспорангиями. После выхода зоо-
спор в пустой зооспорангии врастает
несущая его гифа, которая формирует
внутри новый зооспорангии, но уже
меньшего размера. С наступлением по-
ловой репродуктивной фазы в оогони-
ях (отсюда и название класса) развива-
ются гаплоидные ооциты. Они опло-
дотворяются через оплодотворитель-
ные трубки, прорастающие из
антеридия в оогоний. Высвободившие-
ся из него толстостенные зиготы (оос-
поры) устойчивы к высыханию и пере-
падам температуры и представляют со-
бой стадию покоя. Они распространя-
ются - животными-переносчиками
(птицами, насекомыми и др.) или вет-
ром. При благоприятных условиях оос-
пора прорастает, формируя новую заро-
1
Фитофтороз опустошал картофельные поля Европы в 1840-е годы. Самыми разрушительными
последствия были для Ирландии, где картофель был главной сельскохозяйственной культурой и
всецело доминировал в рационе жителей. Последовавший голод привел к высокой смертности
населения и спровоцировал массовую эмиграцию ирландцев в США. Численность населения
сократилась на четверть в период с 1845 по 1852 г. — Прим. ред.
1
Вторичная зооспора (бобовидная форма, латеральные жгутики) оседает на субстрат и превра-
щается в форму, называемую «шиповатая циста». Если субстрат неподходящ, она трансформи-
руется обратно во вторичную зооспору, которая переплывает на новый субстрат. Этот обрати-
мое преобразование может осуществляться несколько раз. Когда подходящий субстрат найден,
шипы «цисты» замыкаются на нем, а сама «циста» дает начало таллому. Эта цепь событий не
ведет к увеличению количества репродуктивных единиц и потому здесь нельзя говорить о раз-
множении. Вторичная зооспора обеспечивает расселение сапролегнии в водной среде, а ооспо-
ра (см. ниже) — в воздушной среде. — Прим. ред.

дышевую трубку и затем трехмерную
сеть мицелия с зооспорангиями.
В жизненном цикле других предста-
вителей этого класса процесс формиро-
вания первичных и вторичных зооспор,
да и сама стадия зооспор, может отсут-
ствовать. Если зооспоры имеются, они
несут латерально выходящие жгутики.
Зооспорангии распространяются пре-
имущественно ветром, что можно счи-
тать адаптацией к наземному образу
жизни.
Стенка гиф состоит из (3-1,3- и
(3-1,6-глюканов с небольшими вставка-
ми целлюлозы; у некоторых представи-
телей также имеется хитин в составе
стенки. Поглощение питательных ве-
ществ из клеток хозяина идет осмот-
рофно. Характерные запасные пита-
тельные вещества — гликоген и мико-
ламинарин. Последний сходен с запас-
ными веществами бурых водорослей и
диатомовых. Эта специфика метаболиз-
ма и последние молекулярные исследо-
вания указывают на то, что оомицеты
произошли от водорослеподобного
предка, который утратил пластиды и
стал гетеротрофом.
Примеры: Achlya, Albugo, Haptoglos-
sa, Plasmopara, Phytophthora, Pythium,
Saprolegnia.
Тип ALVEOLATA Cavalier-Smith,
1992 — АЛЬВЕОЛЯТЫ
Молекулярные данные (в первую
очередь, построение матриц генных
расстояний для рРНК и белков) показа-
ли, что три крупных таксона — Cilio-
phora, Apicomplexa и Dinoflagellata, —
а также часть бывшего таксона Asceto-
spora (Haplospora) и две небольшие
Система протистов 117
Таблица 11. Предполагаемые
филогенетические отношения
между группами Alveolata
(по разным авторам)
Dinoflagellata
Perkinsozoa
Colpodellidae
Apicomplexa
Ciliophora
группы жгутиконосцев (Perkinsozoa и
семейство Colpodellidae) составляют
единый монофилетический таксон
(табл. 11). Таким образом, те структур-
ные комплексы, которые, согласно преж-
ним воззрениям, имели независимое
происхождение, сейчас считаются гомо-
логичными, унаследованными от обще-
го предка. Так, три разительно различ-
ные по внешней морфологии группы —
инфузории с множественными реснич-
ками, динофлагелляты с двумя жгутика-
ми и лишенные жгутиков Apicom-
plexa — несут в кортикальной цито-
плазме гомологичную структуру, назы-
ваемую соответственно: пелликулярные
альвеолы, амфиесмальные пузырьки и
внутренний мембранный комплекс. Эта
структура унаследована от жившего
порядка 1200 млн лет назад общего
предка, который, имея два жгутика, по-
ходил на современного Perkinsus; спе-
цифическая же особенность его суб-
мембранных покровов состояла в при-
сутствии слоя вакуолей. Кроме того, го-
мологичными структурами, несущими
сходные функции, вероятно, являются
парасомальные мешочки цилиат, окай-
мленные ямки динофлагеллат и микро-
поры Apicomplexa (рис. 79).