Hogg S. Essential microbiology
Подождите немного. Документ загружается.

JWBK011-11 JWBK011-Hogg August 12, 2005 19:21 Char Count= 0
a
c b a
d c b a
e d c b a
50
40
30
20
10
a
b
c
d
e
20 40 60
Time (min)
Percentage of
recombinants
w
ith Hfr
characters
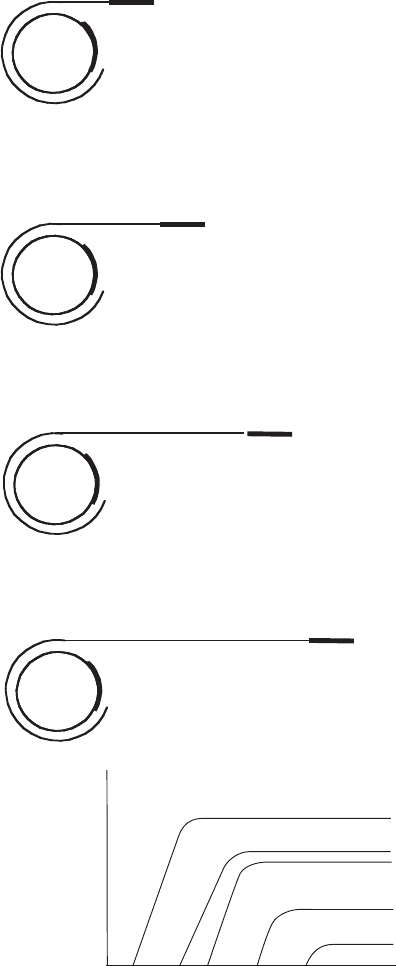
Figure 11.31 In interrupted mating experiments, the conjugation tube is broken after
different time periods. The time at which different genes are transferred reflects their relative
positions on the bacterial chromosome. By plating out onto selective media, the order in
which the different genes are transferred can be determined. The graph shows that the first
genes to enter the F
−
cell are those present in the highest proportion of recombinants
308

JWBK011-11 JWBK011-Hogg August 12, 2005 19:21 Char Count= 0
GENETIC TRANSFER IN MICROORGANISMS 309
Box 11.9 It’s that man again!
Not content with going down in history as the man who first demonstrated conju-
gation in bacteria, Joshua Lederberg was also, in 1952, one of the co-discoverers
of transduction, along with Norton Zinder. Lederberg was to become a dominant
figure in microbial genetics for over half a century. He was even responsible for
coining the term ‘plasmid’!
The integration of the F plasmid into the bacterial chromosome is reversible; thus
Hfr cells can revert to F
+
. Excision of the integrated plasmid is not always precise, and
sometimes a little chromosomal DNA is removed too. When this happens, the plasmid,
and the cell containing it, are called F
(‘F prime’); transfer of the plasmid to an F
−
cell
takes with it the extra DNA from the host chromosome. The recipient genome thus
becomes partially diploid (merodiploid), because it has its own copy, plus the ‘guest’
copy of certain genes.
Transduction (Box 11.9)
In the third form of genetic transfer in bacteria, bacteriophages act as carriers of DNA
from one cell to another. In order to appreciate the way in which this is done, it is
necessary to recall the sequence of events in phage replication cycles discussed in the
previous chapter (see Figure 10.11).
Generalised transduction occurs in virulent phages, that is, those with a lytic life
cycle. Sometimes, the enzymes responsible for packaging phage DNA into its protein
coat package instead similarly sized fragments of degraded chromosomal DNA (Figure
11.32). Despite containing the wrong DNA, this transducing phage particle is still infec-
tive, since this is dependant on its protein element. Thus following infection of another
bacterial cell, the DNA can be incorporated by recombining with the homologous seg-
ment in the recipient cell. Since any chromosomal fragment can be mistakenly packaged
in this way (as long as it finds an area of homology), all genes are transferred at a similar
(low) frequency.
Specialised transduction results in a much higher efficiency of transfer for specific
genes, however it is limited to genes having a particular chromosomal location. Recall
from Chapter 10 that in lysogenic life cycles, the phage DNA is integrated into the
host chromosome, and later, perhaps after many rounds of cell division, excised again
before re-entering a lytic cycle. If this excision does not happen precisely, some of the
adjoining chromosomal DNA, carrying a gene or two, may be incorporated into the
phage particle (we saw a similar mechanism in the case of F
plasmid formation). Upon
infecting another cell, the transduced genes would undergo recombination and become
incorporated into the recipient’s chromosome (Figure 11.33). Although limited to genes
in the vicinity of the lysogenic phage’s integration, this is a highly efficient form of
transfer, since the genes become stably integrated into the host cell.
Transduction experiments, like those involving conjugation, can be used to determine
the relative positions of genes on a bacterial chromosome.
JWBK011-11 JWBK011-Hogg August 12, 2005 19:21 Char Count= 0
310 MICROBIAL GENETICS
a)
b
)
c)
f)
e)
d)
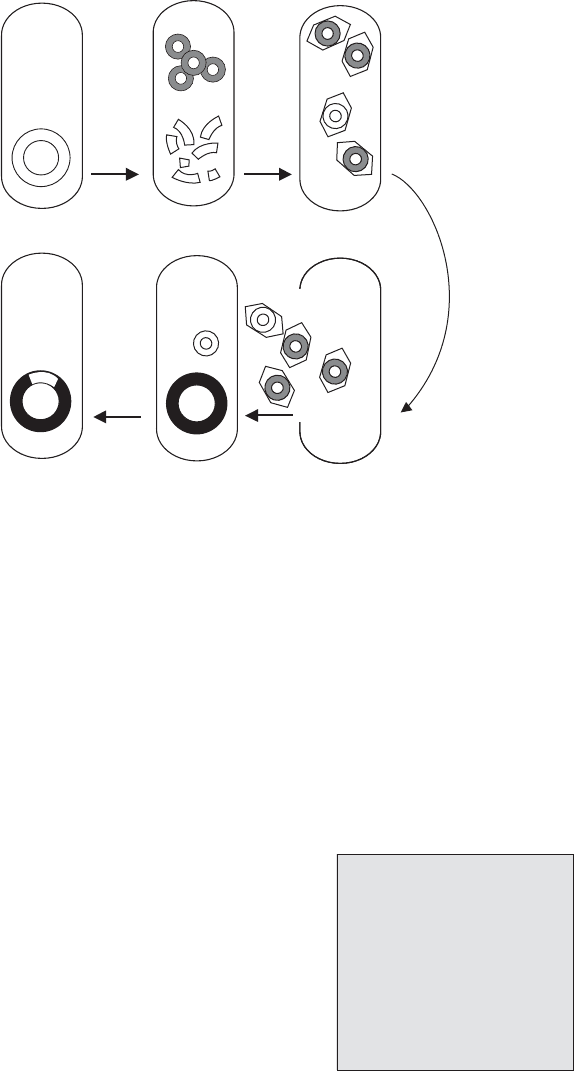
Figure 11.32 Generalised transduction. During the lytic cycle of a phage, the host DNA is
degraded (b), and a fragment may be mistakenly packaged into a newly synthesised phage
particle (c). Upon infecting a new host cell, the transducing phage releases its DNA (e);
although unable to replicate, this can undergo recombination with a homologous sequence
on the host chromosome (f)
Transposable elements
An unusual type of genetic transfer which takes place within an individual cell involves
sequences of DNA called transposable elements. One type is known as an insertion
sequence (IS), a relatively short piece of chromosomal or plasmid DNA which contains
a gene for the enzyme transposase (Figure 11.34). This recognises, cuts and re-ligates
the insertion sequence anywhere in the bacterial genome. In so doing, it may interrupt a
Transposable elements
that also carry genes
other than those re-
quired for transposition,
such as genes for an-
tibiotic resistance or tox-
ins, are known as trans-
posons.
gene sequence, and thereby cause a mutation. Unlike re-
combination events, no homology is required between
the transposable element and the point at which it in-
serts. This relocation of a transposable element from one
place in the genome to another is termed conservative
transposition.Inreplicative transposition, the element
remains in its original position and a copy is made and
inserted elsewhere in the genome. Insertion sequences are
flanked by inverted sequences some 9–41bp in length,
which are thought to be essential for the recognition of
the sequence by the transposase.
JWBK011-11 JWBK011-Hogg August 12, 2005 19:21 Char Count= 0
Integration of phage
and chromosomal
genes into new host
chromosome
Chromosomal
DNA excised
with prophage
Integrated
prophage
Phage
replication and
packaging
Cell lysis and
release of phages
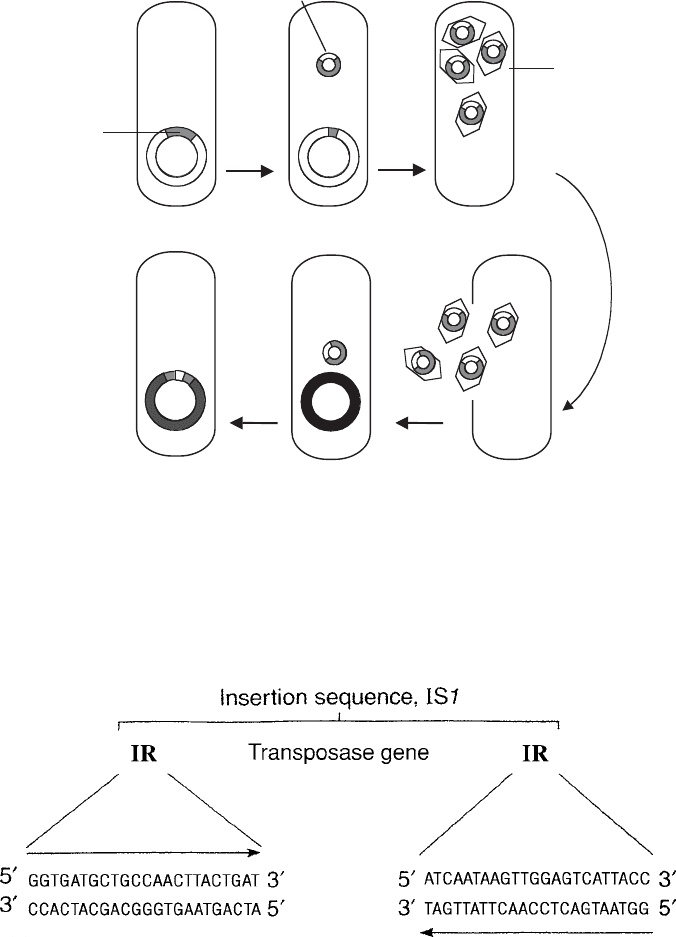
Figure 11.33 Specialised transduction. During the replication cycle of a lysogenic bacte-
riophage, phage DNA is incorporated into the host chromosome (see Figure 10.11). When
a lytic cycle resumes and the phage DNA is excised, it may take with it an amount of sur-
rounding chromosomal DNA. This is packaged into phage particles and infects new host
cells, where it is integrated into the bacterial chromosome. Only genes surrounding the site
of phage integration may be transduced in this way
Figure 11.34 Transposable elements. The insertion sequence IS 1 of E. coli is 768bp in
length and is flanked by 23bp inverted repeat sequences. The IS contains the gene for a
transposase enzyme, which catalyses the movement of the insertion sequence from one loca-
tion to another. By integrating at random points in the genome where there is no sequence
homology, IS sequences may disrupt functional genes and give rise to mutations
311

JWBK011-11 JWBK011-Hogg August 12, 2005 19:21 Char Count= 0
312 MICROBIAL GENETICS
Test yourself
1 In the - model of DNA replication, each strand serves as
a
for the synthesis of a stand. Each daughter molecule
comprises one
strand and one newly synthesised strand.
2 During DNA replication, the point where the two strands become separated
is called the
.
3
III forms a second strand by adding complementary
nucleotides in the
→
direction.
4 During DNA replication, the
must be synthesised dis-
continuously, as a series of
fragments.
5 Mistakes in DNA replication are largely corrected by the cell’s
enzymes. Any errors that persist may lead to .
6 The function of genes was expressed in the one
, one,
hypothesis.
7 The flow of information: DNA → mRNA → protein is often referred to as
the
of biology.
8 In the genetic code, many amino acids are encoded by more than one triplet
sequence; the code is therefore said to be
. Three of the
triplet combinations do not code for an amino acid, but instead serve as
codons.
9 The enzyme RNA polymerase uses a single-stranded
template to
synthesise a complementary strand of
.
10 Transcription begins at a
sequence, situated of the gene.
11 In bacteria, proteins with related functions may be encoded together; the
result of transcription is a
mRNA.
12 In eukaryotes, genes are usually discontinuous; coding regions called
are interspersed with non-coding .
13 Molecules of tRNA act as adapter molecules during translation; at one end
they have a three-base
complementary to a triplet codon, and at
the other end carry the corresponding
.
14 Lactose acts as an
for the three genes that make up the lac operon.
It neutralises the effects of a
protein encoded by the I gene.

JWBK011-11 JWBK011-Hogg August 12, 2005 19:21 Char Count= 0
TEST YOURSELF 313
15 The trp operon contains five genes involved in the synthesis of . The
presence of this substance activates a
, which prevents transcription
of the operon by binding to the
sequence.
16 A
mutation alters the of a gene and will change
the sense of the encoded message. Such a mutation arises through
or of DNA.
17 A
mutation changes a normal codon into a codon, and
results in the premature termination of translation.
18
such as 5-bromouracil mimic the structure of normal
nucleotide bases and become
into the DNA structure.
19 The
test is used to assess the mutagenicity of a substance.
20 In Griffith’s famous experiment,
cells of the virulent
S-strain appeared to pass on the ability to synthesise a capsule to the non-
virulent R-strain. Griffith coined the term
for the factor
responsible.
21 Transformation only occurs between related cells as it depends on the donor
DNA finding a
sequence on the host chromosome with which to
.
22
experiments can be used to map the order in which genes
on a bacterial chromosome are transferred by
.
23 In
transduction, chromosomal genes close to the point of integra-
tion of the
may be excised along with it and be transferred to
another host cell.
24 In
transduction, fragments of DNA are mistakenly pack-
aged into phage coats and can be transferred to another bacterial cell.
25
are sequences of DNA that can move from one location
on a chromosome to another.
JWBK011-11 JWBK011-Hogg August 12, 2005 19:21 Char Count= 0
314

JWBK011-12 JWBK011-Hogg August 12, 2005 20:21 Char Count= 0
12
Microorganisms in Genetic
Engineering
Introduction
In the last 30 years or so, there has been a revolution in the field of genetics, which has
had a profound effect on virtually every other area of biology. This has been due to the
development of new techniques that have enabled scientists to analyse and manipulate
DNA in a quite unprecedented way. Genetically modified crops, DNA ‘fingerprinting’
and gene therapy are just three of the many applications made possible by these ad-
vances. The subject of ‘genetic engineering’ is too huge to be discussed here in detail,
and indeed it extends into areas far beyond the remit of this book. In this chapter, how-
ever, we shall examine some of the ways that microorganisms have contributed to the
genetic revolution. As we shall see, their role in the development of new techniques of
DNA manipulation since the 1970s has been just as important as their earlier contri-
bution to the elucidation of the structure, role and replication of DNA several decades
earlier.
Restriction endonucle-
ases do not destroy the
host bacterium’s own
DNA, because certain
nucleotides in the re-
cognition sequence are
modified by methyla-
tion. The REs are unable
to cleave the DNA at
methylated sites.
The beginnings of genetic engineering can be said to
date from the discovery, in the late 1960s, of a class of
bacterial enzymes called restriction endonucleases (REs).
These are enzymes that cleave DNA into pieces by mak-
ing breaks in the sugar-phosphate backbone; in nature,
they serve to destroy any foreign DNA that may enter
the cell. They do not cut the DNA in a random fash-
ion, however; their unique usefulness to the molecu-
lar biologist lies in the fact that they break the DNA
in a precise and reproducible manner. They do this by
cutting only at specific recognition sites, sequences of
typically four to six nucleotides (Figure 12.1). Thus,
under favourable conditions, a particular RE will di-
gest a given piece of DNA into an identical collection of fragments, time after time.
In the ensuing years, many hundreds of restriction endonucleases have been discov-
ered, many of which recognise different specific sequences, providing biologists with
a hugely versatile tool for the manipulation of DNA, often likened to a pair of
molecular ‘scissors’. Not long after REs were first isolated, they were used to create
the first man-made recombinant DNA molecule (Figure 12.2). This involved cutting
315
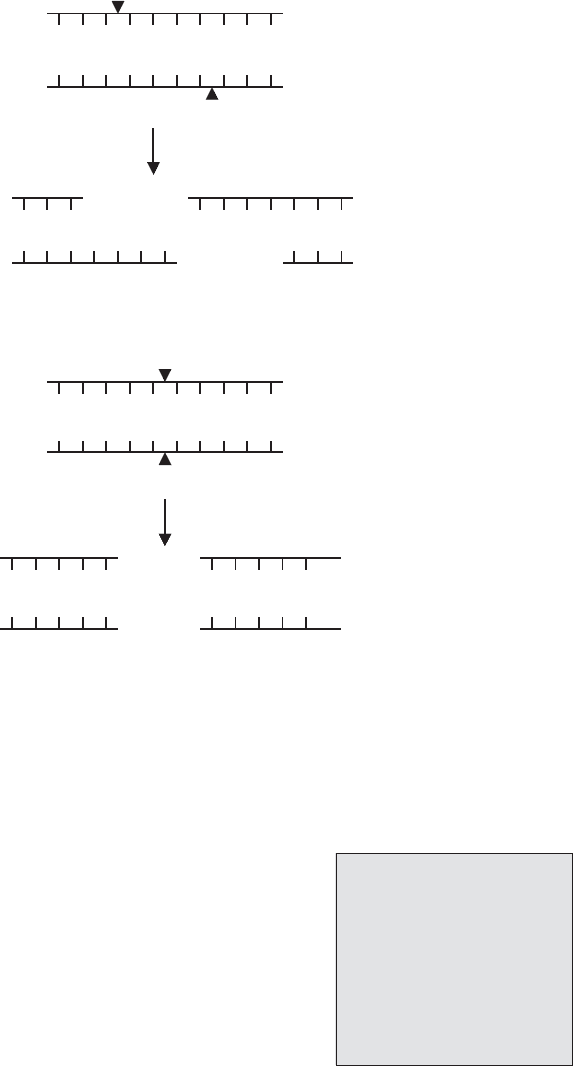
JWBK011-12 JWBK011-Hogg August 12, 2005 20:21 Char Count= 0
316 MICROORGANISMS IN GENETIC ENGINEERING
Site of cleavage
5’ 3’
A A G C T T
T T C G A A
3’ 5’
Site of cleavage
Hind III
5’ 3’ 5’ 3’
A A G C T T STICKY
T T C G A A ENDS
3’ 5’ 3’ 5’
Site of cleavage
5’ 3’
T G G C A A
A C C G T T
3’ 5’
Site of cleavage
Bal I
5’ 3’ 5’ 3’
T G G C C A BLUNT
A C C G G T ENDS
3’ 5’ 3’ 5’
Figure 12.1 Restriction endonucleases fragment DNA molecules by breaking the sugar–
phosphate backbone within a specific sequence of nucleotides. Depending on the site of
cleavage, the fragments so produced may be blunt-ended or ‘sticky’-ended
fragments of DNA from different sources, then using another enzyme, DNA ligase to
join them together, a process facilitated by using fragments with compatible ‘sticky’
Cloning is the produc-
tion of multiple copies of
a specific DNA molec-
ule. The term is also used
to describe the produc-
tion of genetically iden-
tical cells or even organ-
isms.
ends. Remember from Chapter 11 that A always pairs
with T and C with G; because of this, complementary
sequences that come into contact with one another will
‘stick’ together. DNA, it seems, is DNA, wherever it
comes from; consequently DNA from plants, animals,
bacteria or viruses can be joined together to create novel
sequences undreamed of by Mother Nature.
Of course, a single molecule of our newly recombinant
DNA is not much use to us. The important breakthrough
came with the development of cloning – the ability to
produce huge numbers of copies of a given molecule. To
do this, two further things are needed: a carrier DNA molecule called a vector, and a
host cell in which it can be replicated.
JWBK011-12 JWBK011-Hogg August 12, 2005 20:21 Char Count= 0
INTRODUCTION 317
SOURCE 1 SOURCE 2
A A G C T T A A G C T T
T T C G A A T T C G A A
Hind III Hind III
A A G C T T A A G C T T
T T C G A A T T C G A A
Mix fragments and join
with DNA ligase
A A G C T T RECOMBINANT
T T C G A A
DNA MOLECULE
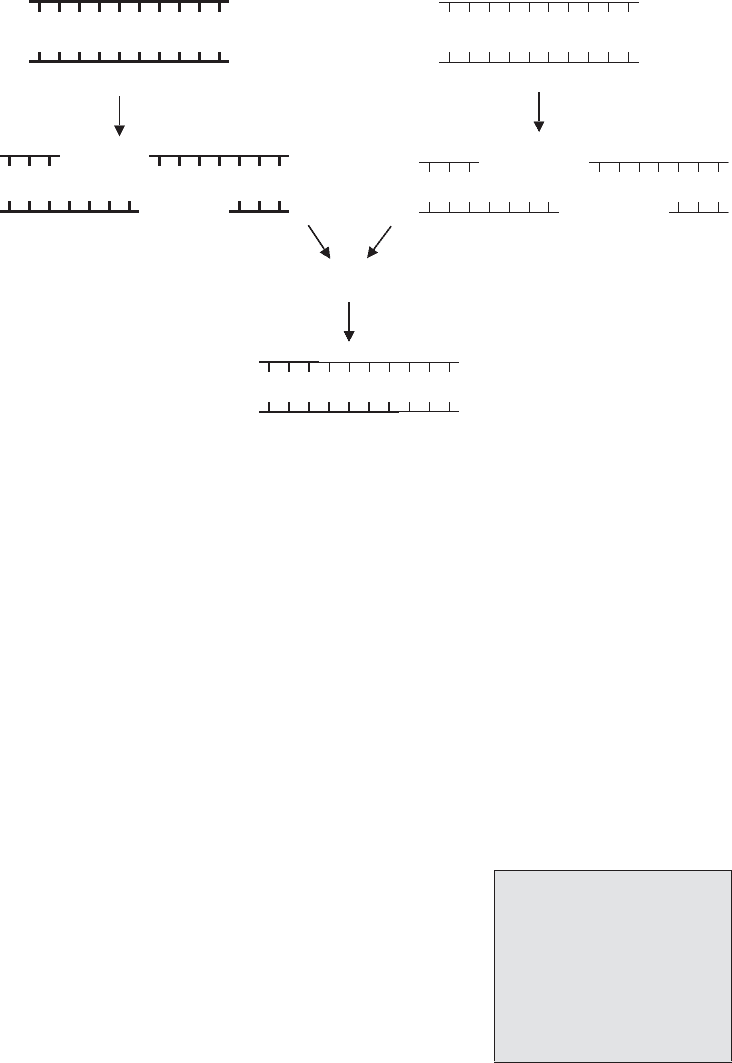
Figure 12.2 DNA from different sources can be joined together. ‘Sticky’-ended restriction
fragments from one DNA source have single-stranded sequences that are compatible with
fragments produced from another source by the same RE. Compatible base pairing attracts
the fragments together and the join is made more permanent by the action of DNA ligase
Figure 12.3 shows the main steps of a cloning protocol:
r
‘donor’ DNA and vector are digested with an RE to provide compatible sticky
ends
r
a fragment of donor DNA is spliced into the vector molecule
r
the recombinant vector gains entry to a host cell (e.g. E. coli )
r
the vector replicates inside the cell, making further copies of the inserted DNA
r
host multiplication results in the formation of a clone of cells, all containing
the same recombinant plasmid – we now have millions of copies of our donor
DNA ‘insert’. A collection of such clones is called a DNA library.
A vector is a self-
replicating DNA mole-
cule used in gene clon-
ing. The sequence to be
cloned is inserted into
the vector, and repli-
cated along with it.
Let us look at role of vectors in a little more detail. The
main features required of a cloning vector are:
r
it must be capable of replicating autonomously inside
a host cell – when it does so, any DNA it carries will
also be replicated. Vectors make multiple copies of
themselves inside the host cell.
r
it must be relatively small – to facilitate manipulation
and entry into a host cell, vectors must not exceed a
certain size.