Klipp E., Herwig R., Kowald A., Wierling C., Lehrach H. Systems Biology in Practice: Concepts, Implementation and Application
Подождите немного. Документ загружается.


Clb5 levels rise and Sic1 is degraded, while at Finish, Clb2 vanishes and Sic1 is newly
produced.
7.3
Aging
Aging is a complex biological process that affects practically all components of an or-
ganism. It is difficult to give a good definition of aging that holds for all organisms,
but for populations a very useful description is that aging consists of all those pro-
cesses that lead to a monotonic increase of mortality with time. Humans, for instance,
have a yearly mortality of roughly 0.1% at age 20, but of about 10% at 80 years of age!
There are two different aspects of the aging process that need to be understood.
First, there is the question of why the aging process has evolved. A process that leads
to an increase of mortality reduces the average life span of the organism and thus re-
duces the number of offspring the organism can have. This, however, should lead di-
rectly to a reduction of evolutionary fitness. It is therefore not trivial to understand
why such a biological trait should have evolved in practically all higher organisms.
The second important question is of course how the actual biochemical mechan-
ism works that leads to the gradual functional decline. Unfortunately, the evolution-
ary models do not predict specific mechanisms that might be involved in aging. As a
consequence a large number of mechanistic models have been proposed. A review
of the available literature showed that more than 300 different mechanistic theories
exist (Medvedev 1990). Figure 7.8 shows a small sample of the most popular me-
chanistic theories. The spatial arrangement of the diagram aims to reflect connec-
tions between the theories. For example, oxygen radicals can damage DNA, leading
to somatic mutations, or damage mitochondrial membranes and DNA, leading to
defective mitochondria. Radical reactions with cytoplasmic or mitochondrial pro-
teins or membranes can also lead to the accumulation of indestructible waste pro-
ducts. And finally, reactions of radicals with macromolecules can lead to the forma-
tion of cross-links that impair their biological functioning.
240
7 Selected Biological Processes
Fig. 7.8 Small selection of the large number of mechanistic theories
of aging. The topology of the diagram is intended to reflect overlap
and points of interaction between different theories.

It is therefore clear that this chapter cannot be a comprehensive treatment of the
aging process. Instead, we will discuss selected models. In Section 7.3.1 we will
study a model that deals with the evolution of the aging process, and in Section 7.3.2
we look at a model that deals with damage to mitochondria.
7.3.1
Evolution of the Aging Process
It is not immediately obvious why organisms should grow old and die. What is the
selective advantage of this trait? And if aging is advantageous, why do different spe-
cies have such widely differing life spans?
The first attempt to explain the evolution of aging was made by Weismann (1891).
He proposed that aging is beneficial by removing crippled and worn-out individuals
from the population and thus making space and resources available for the next gen-
eration. This type of reasoning is very similar to suggestions such as the prevention
of overcrowding or the acceleration of evolution by decreasing the generation time.
These ideas suggest that aging itself confers a selective advantage and that the evo-
lution of genes that bring life to an end is an adaptive response to selective forces.
All these theories have in common that they rely on group selection, the selection of
a trait that is beneficial for the group but detrimental to the individual. However,
group selection works only under very special circumstances such as small patch
size and low migration rates (Maynard Smith 1976). A second major argument
against adaptive theories is the empirically found paucity of old individuals in nat-
ural populations. Extrinsic mortality in natural populations is so high that only very
few individuals survive long enough to be killed by intrinsic mortality. But if intrinsic
mortality, which is caused by the aging process, is not the main cause of death under
natural conditions, it is difficult to see how aging could have evolved for the purpose
of removing animals from the population.
The weaknesses of adaptive theories have been recognized for some time, and
newer theories are no longer based on group selection but rather on the “declining
force of natural selection” with time. This important concept is based on the fact
that even in the absence of aging, individuals in a population are always at risk of
death due to accidents, predators, and diseases. For a given cohort this leads to an ex-
ponential decline over time in the fraction of individuals that are still alive. Events
(e.g., biochemical processes) that occur only in chronologically old individuals will
therefore affect only a small proportion of the whole population. The later the onset
of the events, the smaller the involved fraction of the population is.
Medawar (1952) was the first to present a theory for the evolution of the aging pro-
cess based on this idea. His “mutation-accumulation” theory states that aging might
be caused by an accumulation of deleterious genes that are expressed only late in
life. Because of the declining force of natural selection, only a small part of the popu-
lation would be affected by this type of mutation and the resulting selection pressure
to remove them would only be very weak. Mutations with a small selection pressure
to be removed can persist in a mutation-selection balance and thus explain the emer-
gence of an aging phenotype.
241
7.3 Aging

Another theory of this kind is the “antagonistic pleiotropy” theory (Williams
1957). Genes that affect two or more traits are called pleiotropic genes, and effects
that increase fitness through one trait at the expense of a reduced fitness of another
trait are antagonistic. Now consider a gene that improves the reproductive success of
younger organisms at the expense of the survival of older individuals. Because of the
declining force of natural selection, such a gene will be favored by selection and
aging will occur as a side effect of the antagonistic pleiotropy property of this gene.
Possible candidate genes might be found in males and females. Prostate cancer ap-
pears frequently in males at advanced ages, but it can be prevented by administra-
tion of female hormones or castration. It seems to be a consequence of long-term ex-
posure to testosterone, which is necessary for male sexual, and thus reproductive,
success. In older females osteoporosis is mediated by estrogens that are essential for
reproduction in younger women. In both cases, gene effects that are beneficial at
younger ages have negative consequences later in life.
Genes that trade long-term survival against short-term benefit are probably the
strongest candidates to explain the aging process. A specific version of this hypoth-
esis that connects evolutionary concepts with molecular mechanisms is the “disposa-
ble soma” theory (Kirkwood and Holliday 1986; Kirkwood and Rose 1991). The the-
ory realizes that organisms have a finite energy budget (food resources) that must be
distributed among different tasks like growth, maintenance, and reproduction. En-
ergy spent for one task is not available for another. Organisms have to solve this opti-
mal resource allocation problem such that evolutionary fitness is maximized. On the
basis of quite general assumptions, a mathematical model can be constructed that
describes the relationship between investment in maintenance and fitness. We will
have a closer look at this model as an example how to formulate a mathematical de-
scription of such a qualitative idea.
To get started we need a mathematical concept of fitness. A standard measure that
is often used in population genetics is the intrinsic rate of natural increase, r, (also
called the Malthusian parameter), which can be calculated by numerically solving
the Euler-Lotka equation (Eq. (7-16)). To calculate r for a given genotype, the survivor-
ship function, l(t), and the fertility function, m(t), have to be known. l(t) denotes the
probability that an individual survives to age t and m(t) is the expected number of off-
spring produced by an individual of age t.
R
1
0
e
rt
l tm t dt 1 : (7-16)
If the value of r that solves this equation is negative, it implies a shrinking popula-
tion; if it is positive, the population grows. Thus, the larger r is, the higher the fitness
is. An exact derivation of the Euler-Lotka equation is outside the scope of this chapter
but can be found in Maynard Smith (1989) or Stearns (1992). Investment in somatic
maintenance and repair will affect both survivorship and fertility, and the question
remains whether there is an optimal level of maintenance that maximizes fitness.
Unfortunately, the precise physiological tradeoffs are unknown, so we have to de-
velop some qualitative relationship. In many species mortality increases exponen-
242
7 Selected Biological Processes

tially according to the Gompertz-Makeham equation (Eq. (7-17)) (Makeham 1867).
m
0
, b, and g represent basal vulnerability, actuarial aging rate, and age-independent
environmental mortality, respectively.
m tm
0
e
b t
g : (7-17)
Mortality and survivorship are connected via the relation dl/dt =–m(t)7l(t). By sol-
ving this equation we obtain an expression for l(t) that depends on two factors that are
influenced by the level of maintenance, m
0
and b (Eq. (7-18)). We now define the vari-
able r to be the fraction of resources that are allocated for maintenance and repair,
r = 0 corresponding to zero repair and r = 1 corresponding to the maximum that is
physiologically possible. We also make the assumption that above a critical level of re-
pair, r*, damage does not accumulate and the organism has reached a non-aging state.
The rationale for this postulation is the idea that aging is caused by the accumulation
of some kind of damage and that by investing more in repair, the accumulation rate is
slowed down until finally the incidence rate is equal to the removal rate, in which case
the physiological steady state can be maintained indefinitely. The modifications to m
0
(Eq. (7-19) and b (Eq. (7-20)) are only one way to implement the desired tradeoff (de-
creasing m
0
and b with increasing r), but in qualitative models like this, the principle
results are often very robust with regard to the exact mathematical expression used.
l te
1e
b t
m
0
=bg t
; (7-18)
m
0
m
min
=r ; (7-19)
b b
0
r
r
1
r r
b 0 r > r
:
(7-20)
The level of maintenance also influences fertility, m(t). It is assumed that the age at
maturation, a, will increase with rising r and that the initial reproductive rate, f,isa
decreasing function of r. It is also assumed that fertility declines due to age-related de-
terioration with the same Gompertzian rate term as survivorship. From these condi-
tions equations can be derived for fertility (Eq. (7-21)), age at maturation (Eq. (7-22)),
and initial reproductive rate (Eq. (7-23)).
m tf e
e
b a
e
b t
m
0
=b
; (7-21)
a a
0
=1 r; (7-22)
f f
max
1 r: (7-23)
Now we have all the information necessary to solve the Euler-Lotka equation for
different values of repair, r (Fig. 7.9). The calculations confirm that an optimal level
243
7.3 Aging
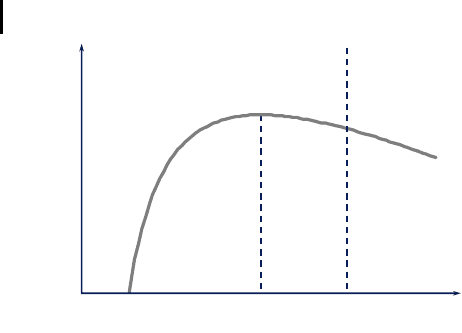
of maintenance, r
opt
, exists, which results in a maximal fitness. Depending on the
ecological niche (represented by environmental mortality, g), the optimal amount of
maintenance varies. In a risky environment the optimum is shifted towards lower
maintenance, and a niche with low external mortality selects for individuals with a
relatively high investment in maintenance.
This fits quite nicely with biological data. Species like mice or rabbits live in a
high-risk environment, and, as predicted, they invest heavily in offspring but have
little left for maintenance. Consequently, their aging rate is high and their life expec-
tancy is low (even under risk-free laboratory conditions). Humans or elephants, by
contrast, inhabit a low-risk environment, expend fewer resources for offspring, and
invest more in repair. Especially instructive are birds, which live two to three times
as long as mammals of comparable body weight. Again, this long life span can be
predicted by the enormous reduction of external mortality that accompanies the abil-
ity to fly and thus escape predators or starvation.
Another important result of the model is that the optimal level of maintenance
and repair is always below the critical level, r*, that is required for the evolution of a
non-aging organism. This result can also be understood intuitively. Since all species
have an external mortality that is above zero, they have a finite life expectancy, even
without aging. This means that even though it might be physiological possible to
have such an efficient repair system that damage does not accumulate with time, re-
sulting in a potentially immortal organism, this never results in a maximal fitness
value. Only in the limit of g = 0 does r
opt
approach r*.
Although the disposable soma theory is very successful in explaining the evolution
of the aging process, it unfortunately does not predict which specific molecular da-
mage accumulates or which repair systems are the most important. All possible me-
chanisms that somehow influence the steady state of the cell are viable candidates.
244
7 Selected Biological Processes
Fitness, r
Investment in Maintenance, U
Aging Non-Aging
U*
U
opt
Fig. 7.9 The disposable soma theory predicts
that the optimal investment in maintenance is
always less than what would be required to
achieve a non-aging state. The exact position of
this optimum (which maximizes fitness) de-
pends on the environmental risk a species is ex-
posed to over evolutionary times. Organisms liv-
ing in a niche with heavy external mortality
should invest less in maintenance than do or-
ganisms that are exposed to little external mor-
tality.

Indeed, this can be taken as an argument that many types of damage accumulate
with time and that many types of repair processes contribute to the rate of aging.
Under those conditions it is not fruitful to study individual biochemical mechanisms
in isolation, but rather as a network of connected processes. The investigation of the
aging process is thus a prime candidate for a systems biological approach.
7.3.2
Accumulation of Defective Mitochondria
Figure 7.8 shows that defective mitochondria play a prominent role in one of the
most favored theories regarding the biochemical mechanism of the aging process.
Mitochondria are not only the powerhouses of the cell, generating the majority of
the cellular ATP, but also are the main producers of reactive oxygen species (ROS).
These reactive molecules damage proteins, membranes, and the mitochondrial
DNA (mtDNA). The mitochondrial theory of aging is based on the fact that damage
to the mtDNA impairs the genes responsible for ATP production but not those in-
volved in the replication of the mtDNA, because they are located in the nucleus.
Thus ROS-induced damage to the mitochondria could turn a symbiont into a para-
site, leading to a progressive decline in the cellular energy supply. Experimental find-
ings of the past few years have confirmed that in aging post-mitotic cells there is in-
deed a clonal accumulation of defective mitochondria with time (Brierley et al. 1998;
Khrapko et al. 1999). This means that a single mutant appeared and expanded clon-
ally, out-competing the wild-type population. The actual molecular mechanism of
this accumulation, however, remains unclear, since it is difficult to see how energy-
starved mutant mitochondria can grow faster than wild-type organelles.
A possible solution to this problem rests on the fact that mitochondria, like pro-
teins, have a certain turnover rate. Newly synthesized mitochondria typically exist
within the cell only for a period of two to four weeks, after which they are degraded
(Huemer et al. 1971; Menzies and Gold 1971). Mitochondrial mutants can therefore
accumulate in a population either by increasing their division rate or by lowering
their rate of degradation. Therefore, it has been suggested that damaged mitochon-
dria accumulate because they have a slower degradation rate (de Grey 1997). If it is
furthermore assumed that defective organelles actually grow more slowly than wild
type (Kowald and Kirkwood 2000), this would explain clonal expansion and avoids
the energy paradox that arises if defective mitochondria are required to have a faster
proliferation rate. The idea is that individual mitochondria are targeted for turnover
in accordance with the level of oxidative damage to their inner mitochondrial mem-
brane. The more the membrane is damaged, the sooner the mitochondrion is de-
stroyed. Defective mitochondria have decreased respiratory activity, and it is there-
fore assumed that they inflict less oxidative damage to their membranes than do
wild-type mitochondria.
In the following sections we give an overview of this model, which has been called
“survival of the slowest” (SOS) hypothesis. For pedagogical purposes we will place
more emphasize on the development of the differential equations than on the actual
simulation results.
245
7.3 Aging
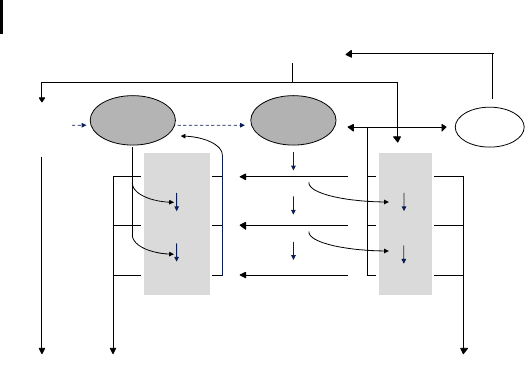
Figure 7.10 gives an overview of the model, which separates the mitochondrial
population into two major classes, intact mitochondria with no damage to their
DNA (M
Mi
) and defective organelles that have acquired some form of mtDNA da-
mage (M
DMi
). Furthermore, mitochondria also accumulate damage to their mem-
branes and a further subdivision is therefore necessary. Both major classes are di-
vided into three additional groups based on the amount of membrane damage.
M
M1
and M
DM1
stand for mitochondria with no or only slight damage to their
membranes, M
M2
and M
DM2
contain a medium amount of damage, and M
M3
and
M
DM3
contain a large amount of damaged membranes. Mitochondrial turnover is
proportional to the level of membrane damage, so the relation g > b > a holds for
the decay rates. RAD
M
represents the radical concentration in intact mitochondria,
and RAD
DM
represents the level in damaged mitochondria. RAD
M
can interact
with the membranes of intact mitochondria with a rate k
M
, shifting mitochondria
into a higher membrane damage class, or it can damage the mitochondrial DNA
(with a rate k
D
), converting intact mitochondria into defective ones. Once mito-
chondria have suffered DNA damage, further reactions with radicals (now RAD
DM
)
can only increase the amount of membrane damage. Finally, the model contains a
generic antioxidant species (AO
x
) that destroys radicals. The main reason for in-
cluding such a species in the model is to provide a sink for radicals, which other-
wise would increase without limit.
The model consists of nine ordinary differential equations describing the time
course of the different molecular species. Six equations are needed for the various
types of mitochondria, one for the level of antioxidants, one for the ATP level, and
one for the amount of radicals. Although the model contains different radical levels
246
7 Selected Biological Processes
G
D
E
J
D
E
J
ATP
mRNA
AO
x
DecayDecay
D
ecay
M
M3
M
M2
M
M1
M
DM3
M
DM2
M
DM1
RAD
DM
RAD
M
k
M
k
M
k
M
k
M
k
D
k
D
k
D
Fig. 7.10 Reactions described by the survival of
the slowest model. Two main classes of mito-
chondria are considered, with and without DNA
damage (M
DM
, M
M
). These are further divided
into subclasses containing little, medium, and
much membrane damage. The different classes
of mitochondria produce different amounts of
ROS, which cause the transition of mitochondria
from one damage class to another. Finally, the
mitochondria are degraded with a rate constant
that is proportional to the amount of membrane
damage. Further details are given in the text.

for damaged and intact mitochondria, it turns out that under certain assumptions
those values can be calculated from one equation (see Section 7.3.2.2).
7.3.2.1 Synthesis Rates
Mitochondria are continually being turned over, which means that new mitochon-
dria must be synthesized to balance degradation. It is assumed that the synthesis
rate is controlled by the cellular energy level. A low ATP concentration stimulates
mitochondrial growth, while a high concentration diminishes it. This is known as
product inhibition. The mathematical expression used to simulate this behavior is
given by Eq. (7-24).
k
1
1
ATP
ATP
c
n
: (7-24)
This artificial promoter has been used successfully in other models (Kowald and
Kirkwood 1994, 1996) and is also employed here. Depending on the amount of ATP,
it results in a value between zero and k
1
, which can be interpreted as the activity of
this promoter. If ATP is equal to ATP
c
, the promoter control parameter, activity is re-
duced to 50 %. The constant n determines how sensitive the promoter is to devia-
tions of the energy level from ATP
c
.
Although mitochondria contain their own genetic material, they cannot be treated
as self-replicating entities because the genes coding for the overwhelming majority
of their proteins are located in the nucleus and these proteins have to be imported
from the cytoplasm. Therefore, the synthesis rate is not proportional to the number
of existing mitochondria but has a fixed upper limit, k
1
.
Equation (24) is further modified to take account of the fact that the synthesis of
macromolecules requires energy and that the synthesis rate is proportional to the ATP
concentration in the cell. Hence the expression is multiplied by ATP/(ATP+ATP
c
).
This construct has the attractive property that new mitochondria are produced only if
energy levels are low, but that biosynthesis works best with a high-energy charge.
The resulting expression describes the total number of new mitochondria that are
synthesized per time interval, but it does not specify to which class they belong. If all
classes of mitochondria were to grow at the same rate, the total number would sim-
ply have to be distributed proportionally between the six different classes. However,
because of the assumed growth disadvantage of defective mitochondria, the contri-
butions to the different classes have to be weighted. All mitochondria with DNA da-
mage have the same growth disadvantage, GDF, because their oxidative phosphory-
lation is nonfunctional. GDF is the factor by which the growth rate is reduced com-
pared to intact mitochondria. The same growth disadvantage is assumed for mito-
chondria with intact DNA but with the highest amount of membrane damage
(M
M3
). For mitochondria with intact DNA and a medium amount of damage (M
M2
),
the growth disadvantage is assumed to be (1+GDF)/2. Taking these considerations
into account, the fraction of the total number of new mitochondria that belongs to a
given damage class is given by
247
7.3 Aging

1
M
M1
2
GDF 1
M
M2
1
GDF
M
M3
M
DM1
M
DM2
M
DM3
X ; (7-25)
where X specifies mitochondria of the given damage class. In the case of intact mito-
chondria, X = M
M1
, for intact mitochondria with a medium amount of membrane
damage, X =2/GDF +17M
M2
, and for all other classes, X = M/GDF.
7.3.2.2 Radical Levels
Figure 7.10 shows that the model includes two different radical pools. Rad
M
is the
number of radicals in a single intact mitochondrion and Rad
DM
is the number of ra-
dicals in a mitochondrion that suffered damaged to its DNA. It is assumed that radi-
cals are generated at a fixed rate and that their removal is proportional to the existing
amount of radicals. The equations can therefore be written as
dRad
M
dt
k
R
f Rad
M
(7-26)
and
dRad
DM
dt
RDF k
R
f Rad
DM
; (7-27)
where k
R
is the rate of radical production of intact mitochondria and it is assumed
that the production rate of defective mitochondria is increased by a factor RDF, the
radical difference factor. In the complete model, f is a function that depends on the
amount of antioxidants per mitochondrion (see Eq. (7-38)), but for any instant of
time it is identical for Eqs. (7-26) and (7-27). These equations can be solved analyti-
cally, and if Rad
M
(t=0) = Rad
DM
(t=0) = 0, it holds that Rad
DM
/Rad
M
= RDF. There-
fore, only one equation is necessary to describe the time course of radicals in this
model.
7.3.2.3 Dilution of Membrane Damage
An important aspect of the SOS hypothesis is that fast mitochondrial growth acts as
a rejuvenation mechanism because it dilutes membrane damage. When newly
synthesized components are incorporated into the mitochondrial membrane, the
preexisting level of membrane damage is reduced and the new damage level, NDL,
is the result of mixing existing membrane components with the new ones. If, for ex-
ample, the amount of new membrane components is equal to the existing ones,
NDL is 50%. This can be formalized and it holds that for all mitochondria with a
growth disadvantage of GDF (M
DM1
, M
DM2
, M
DM3
, M
M3
)
NDL
M
DM
S
M
DM
GDF
M
DM
GDF
S GDF
; (7-28)
248
7 Selected Biological Processes

where S is the total amount of newly synthesized mitochondria as defined by
Eq. (7-40) (see below). In this model, we used three different classes to group the
amount of membrane damage into low, medium, and high. If NDL is, for instance,
75%, we need to know how this affects the number of mitochondria in the three da-
mage classes. This situation is depicted in Fig. 7.11. The minimal amount of mem-
brane damage is zero and the maximum amount is defined as one. The different da-
mage classes are represented by dividing the range 0–1 into three equal segments.
Under the assumption that the membrane damage is evenly distributed between 2/3
and 1 (upper gray box), a 25% reduction (NDL = 0.75) means that the new distribu-
tion of membrane damage is given by the lower gray box ranging from 0.5 to 0.75.
From this diagram we can now calculate the fraction of mitochondria leaving M
3
(valid for NDL > 2/3):
2=3 2=3 NDL
1=3 NDL
2
S
GDF
: (7-29)
Of course, not only do some mitochondria leave M
3
and enter M
2
, but also some
mitochondria that were originally in M
2
become shifted to M
1
. Following the same
line of reasoning as above, this fraction turns out to be (valid for NDL >1/2)
1=3 1=3 NDL
1=3 NDL
S
GDF
: (7-30)
The calculation of NDL is slightly different for M
M2,
which has a growth disadvan-
tage of (GDF + 1)/2. Here NDL
M
M2
GDF 1
2 S GDF 1
and the fraction leaving M
M2
is
2 S
GDF 1
. We do not need to be concerned about M
M1
, since their membrane sta-
tus cannot improve.
7.3.2.4 The Equations
From the assumptions and details given in the previous sections, the differential equa-
tions of the model can be developed. Although the equations might in some respects
have been written in a more compact way, we chose this representation to improve
clarity. The different terms of Eqs. (7-31)–(7-36) describe the consequences of growth
and radical damage on the numbers of mitochondria in the various classes. To illus-
249
7.3 Aging
M1 M2 M3
0 1/3 2/3
1
NDL = 0.75
Fig. 7.11 Mitochondrial growth dilutes membrane
damage. The incorporation of new proteins and lipids
into existing mitochondrial membranes leads to a re-
duction in the level of membrane damage. The maxi-
mum amount of membrane damage is defined as 1,
and mitochondria are assigned to one of three classes
(M
1
, M
2
, M
3
), depending on the degree of damage.
The new damage level (NDL) can be calculated ac-
cording to Eq. (7-28) and the diagram shows how this
affects mitochondria that were originally in M
3
.