Vaccari D.A., Strom P.F., Alleman J.E. Environmental Biology for Engineers and Scientists
Подождите немного. Документ загружается.

monooxygenase pathways involve addition of a hydroxyl g roup to the ring in three dif-
ferent positions (ortho-, meta-, and para-), whereas the fourth involves oxidation of the
methyl group. For all of the pathways, the next step will be cle avage (breaking) of the
ring (usually between the two hydroxyl groups), leading to products (di carboxylic
acids) that after a few more transformations can enter the Krebs cycle. Detailed pathway
information is now available for many degradation reactions (e.g., see the University of
Minnesota Biocatalysis/Biodegradation Database Web site.)
Oxygenases, like most other enzymes, tend to be fairly specific. However, some degree
of nonspecificity can be beneficial, in that it can allow an organism to utilize a newly
available substrate, even if at a lower efficiency. Over an extended time, it can be expected
that new forms of the enzyme, better adapted to the new substrate, will emerge (through
random variation, genetic exchange, or rearrangement), giving the organism utilizing
them a selective advantage when the substrate is available.
On the other hand, a nonspecific enzyme may transform a nontarget substrate, yielding
a product that cannot be further metabolized by the cell. For example, a cell growing on
toluene might accidentally cleave an aromatic ring containing a chlorine substitution. The
resulting dicarboxylic acid may not be suitable for further metabolism because of the
large chlorine atom still present. Then the cell has wasted the material and energy it
invested in transforming a compound that provides it with no benefit. This type of activity,
in which a cell growing on one compound ‘‘mistakenly’’ transforms another without any
benefit, is referred to as cometabolism (Figure 13.12). In this case, over time it can be
expected that the organism will develop more specific enzymes to prevent the drain of
CH
3
H
Cl Cl
ClH
toluene
Cl
Cl
Cl
a
a
b
b
biphenyl
trichlorobiphenyl
pyrene
benzo(a)
py
rene
cc
trichloroethylene
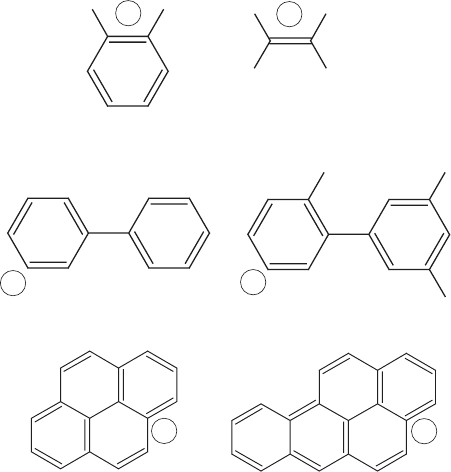
Figure 13.12 Possible cometabolism. The main (growth) substrate in each pair is located on the
left, with areas of similarity indicated by a, b, or c.
408
MICROBIAL TRANSFORMATIONS
cometabolism, or alternatively, it may develop new enzymes and pathways so that the
dead-end product can be further utilized. Although it is not beneficial to the microorgan-
ism, cometabolism may be desirable from a human point of view because it may be pos-
sible to exploit it to degrade otherwise nondegradable compounds.
Even with energy-yielding metabolism, dead-end products may form. In other cases,
intermediates may accumulate because their rate of production is greater than their rate of
further transformation. Occasionally, this accumulating compound will be more toxic than
the parent material, leading to at least a temporary increase in toxicity during biodegrada-
tion. The increased toxicity may have the effect of further slowing degradative activity, as
well as having other undesirable effects on the ecosystem.
In aero bic systems it is common for a single species of bacteria to be able to utilize a
single organic compound as its sole source of carbon and energy. However, occasionally,
organisms of one species can degrade the compound only partially, and those of another
species will further transform, and probably mineralize, the intermediate produced. Such a
combination of organisms of different species ‘‘working together’’ to metabolize sub-
strates is referred to as a consortium (plural, consortia).
The activities of consortia can make detection of cometabolism difficult, as a cometa-
bolic product may be degraded by other organisms before it accumulates sufficiently to be
noticed. Also, the fact that a compound can be completely degraded by a single organism
does not necessarily mean that this is the way it will be degraded in a particular
environment—the actual degradation still may be by a consortium.
Under anaerobic conditions, consortia are com mon. A wide variety of species may be
involved in hydrolyzing, or solubilizing, complex organics, followed by fermentations of
the subunits produced. Then sulfate-reducing or methanogenic organisms may utilize the
fermentation products, leading to mineralization.
The organisms in a microbial consortium may be only loosely associated, or they may
be so closely linked that they are difficult to separate. A classic example is the case of
methane production from ethanol. For many years this was attributed to Methanobacillus
omelianskii, an ‘‘organism’’ that could be grown in ‘‘pure’’ culture with ethanol as the
sole carbon and energy source. However, it was later demonstrated that, in fact, the cultur e
contained two species: a bacterium that converted ethanol to acetate, and a methanogen
(Archaea) that utilized the acetate, producing methane and carbon dioxide.
Occasionally, the balance among organisms in an anaerobic consortium is disturbed
and mineralization does not occur. In the absence of sulfate, anaerobic mineralization
is dependent on methanogens. Compared to the wide variety of organisms that are able
to hydrolyze and ferment organics anaer obically, methanogens are a relatively limited
group of strictly anaerobic archaea. Although most methanogens utilize acetic acid as a
substrate, as a group they are sensitive to low pH. If the acetic acid is produced too
quickly, the pH of the system will drop, inhibiting methanogenic activity. In turn, this
leads to a further buildup of acid, a greater drop in pH, and even lower rates of methano-
genesis and acid destruction.
Some human foods, such as pickles and sauerkraut, are preserved in this way. Ensi-
lage, the process of making silage, is a means of animal feed preservation utilizing
acid anaerobic conditions that has been employed in agriculture for many years. Fresh
corn, hay, or other feed crops are placed in a silo, where they quickly ferment. The
acid anaerobic conditions then prevent substantial further degradation. However, if oxy-
gen is introduced in large amounts, the organic acids are quickly mineralized, pH rises,
and biodegradation resumes.
CARBON 409
Acid anaerobic conditions are also responsible for the preservation of the large masses
of spongy rich organic material in bogs. This waterlogged peat soil, composed mainly of
dead sphagnum moss (an acid-loving plant), accumulates over hundreds or even thou-
sands of years. If the water is drained from such systems, aerobic conditions develop
and the organics are quickly oxidized.
During waste treatment, acid anaerobic conditions are undesirable because they slow
degradation, leading to preservation of the waste. In poorly run leaf ‘‘composting’’
(Section 16.2.3) operations (really, leaf dumps), large piles of leaves may be formed
and simply left unattended. Acid anaerobic conditions develop quickly, so that on opening
such piles 10 years later, the tree species of individual leaves can still be determin ed.
Similarly, in anaerobic digesters for sludge treatment (Section 16.2.1), overloading or
toxic shocks may lead to acid anaerobic conditions that prevent methane formation.
The digester under such conditions is referred to as having gone ‘‘sour.’’
Another problem with acid anaerobic conditions in waste treatment is the potential for
odors. The organic acids themselves may be volatilized, leading to a sour smell. Often,
some alcohols are also formed, and apparently react with the acids to give esters, which
may have a sweet smell. Thus, such systems often have a sweet–sour smell. This is not
necessarily unpleasant for silage in an agricultural setting, but it is often not appreci ated
near wastewater treatment, composting, and landfill sites.
If sufficient proteinaceous material is present in the waste, more severe odors may also
develop. Under most conditions the amino acids released by protein hydrolysis are dea-
minated; that is, the amino group is removed, releasing ammonium (Figure 13.13). This
can lead to an ammonia odor at high pHs where volatilization is favored. However, under
acid anaerobic conditions, amino acids may instead be decarboxylated. This leaves an
amine, some of which are highly odorous, with names such as putrescene and cadavarene.
Another odor concern with anaerobic conditions is hydrogen sulfide. This can be
formed through the reduction of sulfate during anaerobic respiration (Section 13.3.1) or
through the releas e of reduced sulfur from organic compounds such as the amino acids
methionine and cysteine.
In some cases, under both aerobic and anaerobic conditions, rather than b eing miner-
alized, organics are polymerized to form products with long-term stability. One example
is with nitroaromatic compounds such as trinitrotoluene (TNT). Because the aromatic ring
NH
2
CH
C
O
OH
R
CH
R
NH
2
H
H
CH
C
O
OH
R
amino acid
amine
or
g
anic acid
deam
ination
decarboxylation
+
CO
2
+
NH
3
Figure 13.13 Deamination and decarboxylation of an amino acid. (R represents the specific amino
acid side-chain.)
410
MICROBIAL TRANSFORMATIONS
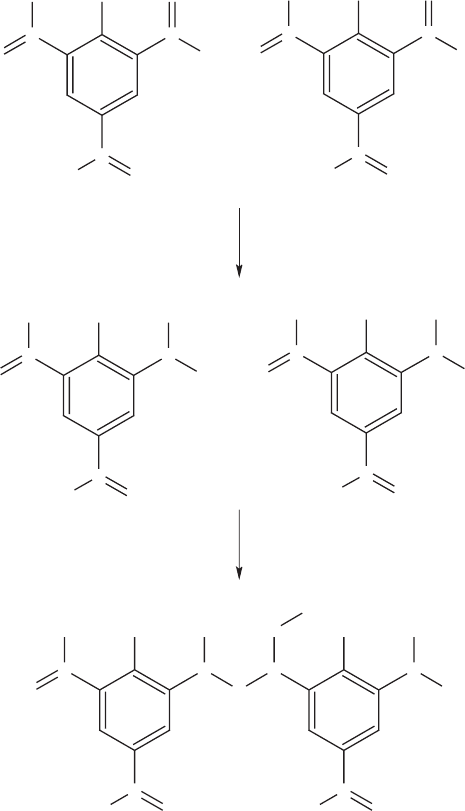
is stabiliz ed by the presence of the nitro groups, TNT is recalcitrant. Although it can be
mineralized, biodegradation usually involves reduction of one or more of the nitro groups
to an amine, which then reacts with a nitro group on an adjacent molecule (Figure 13.14).
A series of such reactions leads to the disappearance of TNT through the formation of a
polymer. The environmental significance of such polymers is still not thoroughly under-
stood, as there is concern that the monomers may be released under some conditions.
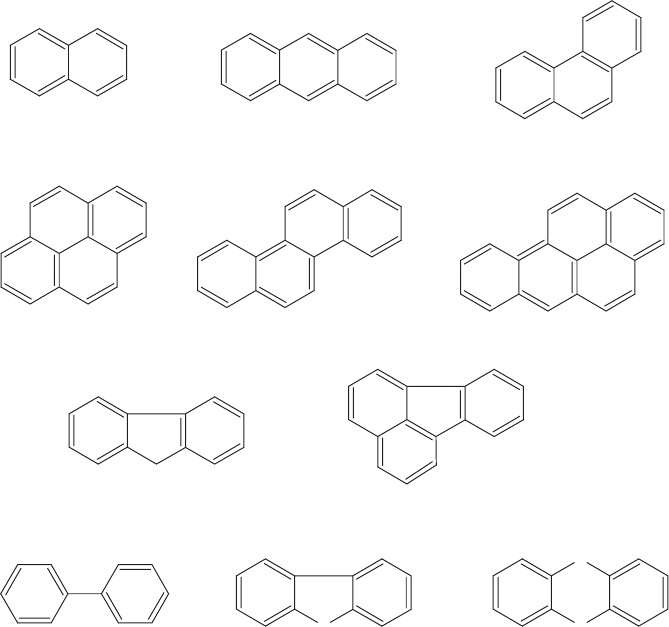
Polynuclear aromatic hy drocarbons (PAHs or PNAs) are compounds containing two or
more condensed (fused together) aromatic rings (Figure 13.15). They are common con-
stituents of coal, oil, and creosote (a wood preservative) and are also products of incom-
plete combustion. They include the first known human carcinogen, benzo[a]pyrene
CH
3
N
+
O
−
O
−
O
−
O
−
O
−
O
−
O
−
O
−
O
−
O
−
O
−
O
−
O
N
+
O
−
O
N
+
O
trinitrotoluene
CH
3
N
H
H
N
+
O
N
+
O
2-methyl-3,5-dinitroaniline
CH
3
N
H
H
N
+
O
N
+
O
CH
3
N
H
N
+
O
N
+
O
CH
3
N
H
H
N
+
O
N
O
O
H
CH
3
N
+
O
N
+
O
N
+
O
Figure 13.14 Polymerization of trinitrotoluene.
CARBON 411
(BAP), which was found to be a cause (because of its presence in soot) of testicular cancer
in English chimney sweeps.
Generally, the PAHs with two or three rings are readily biodegradable, at least under
aerobic conditions. The four-ring compounds pyrene and chrysene are also usually miner-
alized, although chrysene more slowly. However, there usually seems to be little miner-
alization (with occasional exceptions) of the higher-molecular-weight PAHs. BAP does
appear to be cometabolized to some extent. Probably as a result of initial cometabolic
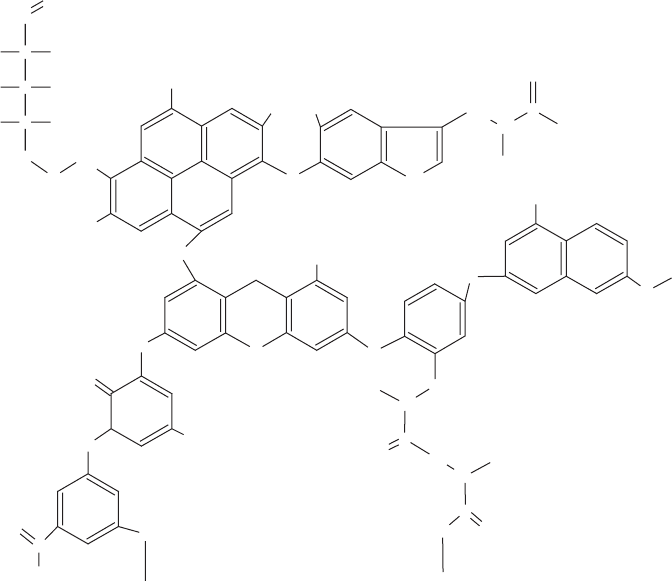
activity, it also appears to be incorporated into the soil humic materials. These complex,
irregular soil organic polymers, such as humic acid (Figure 13.16), include phenolics, het-
erocyclics, sugars, and amino acid residues as building blocks, some probably derived
from lignin degradation. Once a BAP or other PAH residue is incorporated into the poly-
mer, it probably cannot be distinguished from other subunits, and thus is no longer
expected to be of concern.
Over long periods of time, humic materials buried in anaerobic layers may be reduced
further. Eventually, after millions of years, a combination of microbial degradation and
abiotic processes at elevated temperatures and pressures has led to coal and oil formation.
These organic materials have thus been held out of the active cycling of carbon for
naphthalene
anthracene
phenanthrene
fluorene
O
O
O
dibenzofuran dibenzodioxin
biphenyl
pyrene
chrysene
benzo(a)pyrene
fluoranthene
Figure 13.15 Polynuclear aromatic hydrocarbons and related compounds.
412
MICROBIAL TRANSFORMATIONS
100 million years or more. The rapidly increasing use of these fossil fuels over the last
century is responsible for an increase in atmospheric CO
2
as this carbon reenters active
cycling.
Note that although humic materials are not usually referred to as recalc itrant, they do
not biodegrade rapidly. Rather, they are considered to be stable or to change only slowly.
Stabilization of wastes thus refers to elimination of the readily degradable components
through both mineralization and conversion to high molecular weight products such as
humics.
Greenhouse Gases In most cases, a desired major product of biodegradation is carbon
dioxide. However, it is now recognized that atmosp heric CO
2
concentrations have
increased dramatically during the industr ial age, from around 280 ppm prior to the late
eighteenth century to 380 ppmv today. Most of this increase is from the combustion of
fossil fuels, although some other human activities, such as deforestation, loss of soil orga-
nic matter, and draining of wetlands, may also have had a small role. The highe r concen-
tration of CO
2
acts to capture more of the infrared energy radiated from Earth’s surface.
This phenomenon, often referred to as the greenhouse effect, has contributed to an
increase in average temperature worldwide, or global warming. Thus, some thought is
now being given to slowing mineralization of certain organic materials and perhaps to
C
O
CH
C
C
C
CH
2
O
O
HOH
HOH
HOH
O
C
O
CH
CH
2
NH
2
NH
OH
OH
O
OH
OH
O
CH
2
O
C
O
CH
NH
CH
3
C
O
CH
NH
CH
3
O
O
O
OH
O
O
O
CH
3
CH
3
CH
3
CH
3
CH
3
O
OH
CH
3
Figure 13.16 Example of a possible humic acid chemical structure.
CARBON 413
rebuilding some stores of humic materials in soil, for example. This may be an area in
which environmental engineers and scientists can make contributions in the future.
Carbon dioxide is the most abundant, but it is not the only greenhouse gas. In fact,
methane has about 26 times the heat-trapping capacity of CO
2
. Additionally, its concen-
tration in the atmosphere has increased even more (proportionally) than that of carbon
dioxide, from about 0.7 to almost 1.8 ppm. Much of this increase is from fossil-fuel
extraction and use, but other major anthropogenic sources include cattle herds (digestive
tracts), rice fields (flooded soils), and solid waste landfills. Even landfills with gas collec-
tion systems produce fugitive emissions. This is another area in which environmental pro-
fessionals can expect to be called upon to help control greenhouse gas releases.
13.2 NITROGEN
Nitrogen is probably second only to carbon in the complexity of its cycling. Its roles in
living things are likewise diverse, its organic compounds contributing to physical form
(e.g., cells walls, exoskeletons, fingernails, hair), genetics (nucleic acids), and metabolism
(enzymes).
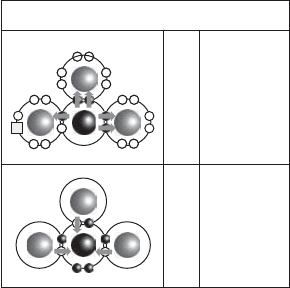
With AN ¼ 7, nitrogen has seven electrons, five of them in its outer shell. This results
in a range of oxidation states from 3toþ5 (Table 13.1). These extremes are represented
by ammonia- and nitrate-nitrogen, respectively (Figure 13.17). Many of the intermediate
forms exist naturally, but 3 (ammonia and organic nitrogen), 0 (elemental N
2
, also
called dinitrogen), þ3(nitrite), and þ5 (nitrate) are most common in soil, water, and
biomass. Other fairly common states include 1(hydroxylamine,NH
2
OH, an intermedi-
ate in the oxida tion of ammonia), þ1(nitrous oxide,N
2
O, also known as laughing gas,
which is an intermed iate in nitrite reduction), þ2(nitric oxide, NO, another intermediate
in nitrite reduction and also an important air contaminant), and þ4(nitrogen dioxide,
NO
2
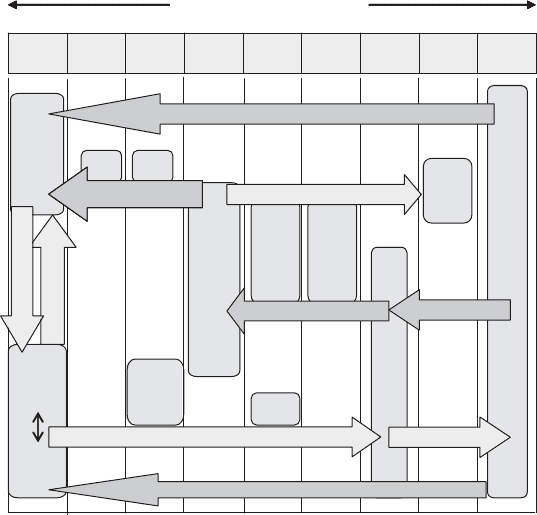
, another air contaminant). In Figure 13.18 these forms are shown as part of the nitro-
gen cycle schematic.
The major accessible source of nitrogen in the biosphere is the elemental nitrogen gas
(N
2
) that makes up 78% (by volume) of the atmosphere. However, the ability to utilize
elemental nitrogen is limited to a few groups of bacteria and archaea. Thus, nitrogen is
O
+5
-3
Nitrate
NO
3
Nitrate
NO
3
-
Ammonia
NH
3
NITROGEN
O
N
O
O
O
H
N
H
H
--
Figure 13.17 Nitrogen oxidation state extremes.
414
MICROBIAL TRANSFORMATIONS
a limiting nutrient in many environments, especially for photoautotrophs. Combined
nitrogen, whether organic or inorganic, is referred to as fixed-N. In addition to microbial
activity, nitrogen is naturally fixed in small amounts through photochemical reactions and
electrical discharges (lightning) in the atmosphere. Beginning in the early twentieth cen-
tury, large amounts of nitrogen have also been fixed through fertilizer production, and to a
lesser extent, through high-temperature combustion (automobiles, electric power genera-
tion, and industrial processes) and explosives manufacturing. These human activities now
account for almost 4 10
7
metric tons per year, more than 20% of all fixation. As of yet,
this large increase in nitrogen fixation on a global scale does not seem to have ‘‘over-
loaded’’ other parts of the nitrogen cycle.
In addition to the essential role of microorganisms in nitrogen fixation, there are only a
limited number of prokaryotes and a few fungi that can oxidize nitrogen. And although
many prokaryotic and eukaryotic microorganisms and plants can utilize inorganic nitro-
gen in the form of ammonium and/or nitrate, many other microorganisms and all animals
can obtain their nitrogen only from organic forms.
When referring to the concentrations of nitrogen-containing compounds, it is common
to report them on the basis of the amount of N present rather than the amount of the com-
pound. This makes it easier to keep track of the nitrogen as it is converted from one form
to another. Thus, if 1.0 mg/L of NH
3
-N is completely converted to nitrate, it yields 1.0
mg/L of NO
3
N. If this same conversion was reported as the concentration of the species
themselves, it would instead be
1:21 mg=LNH
3
! 4:43 mg=LNO
3
Ammonium
NH
4
+
-3
+4+3
+2
+1
0
-1
-2 +5
-3
+4+3
+2
+1
0
-1
-2 +5
-3
+4+3
+2
+1
0
-1
-2 +5
-3
+4+3
+2
+1
0
-1
-2 +5
Hydroxyl-
amine
NH
2
OH
Nitrogen
N
2
Nitroxyl
NOH
Nitrite
NO
2
-
Nitrate
NO
3
-
Nitrification Nitrification
Denitrification Denitrification
NH
3
Ammonia
Nitrogen Oxidation States
Nitrous
Oxide
N
2
O
Nitric
Oxide
NO
N
2
H
4
N
2
H
2
Organic-N
Assimilation
Ammonification
Nitrogen
Dioxide
NO
2
Abiotic
Nitrate Reduction
Nitrate Assimilation
Nitrogen Fixation
Figure 13.18 Biochemical nitrogen transformations.
NITROGEN 415

since for ammonia,
17
14
ð1:0mg=LÞ¼1:21 mg=L
and for nitrate,
62
14
ð1:0mg=LÞ¼4:43 mg=L
where the atomic weight of nitrogen ¼14, the molecular weight of NH
3
¼ 17, and the
formula weight of NO
3
¼ 62.
Example 13.3 The drinking water standard for nitrate-N is 10 mg/L. How much is this
when it is expressed as nitrate?
Answer The formula weight for nitrate ¼ 1ð14Þþ3ð16Þ¼62 g/mol:
62 g=mol nitrate
14 g=mol N
ð10 mg=LNÞ¼44:3mg=L nitrate
In addition to the oxidation and reduction reactions discussed in more detail below,
several other reactions are important in the nitrogen cycle. One of these is the con-
version of organic nitrogen to ammonia. As with any change from an organic form
to an inorganic one, this process can be referred to as mineralization. However, a
more specific name, ammonification, is often used in this case. The reverse react ion,
assimilation, refers to the uptake of an element for incorporation in cell material.
Ammonia assimilation does not represent a change in the oxidation state of the nitro-
gen. However, nitrate assimilation (as discussed below in Section 13.2.1) represents a
reduction and requires energy.
Another im portant nitrogen transformation without oxidation or reduction included
in Figure 13.18 is the acid–base reaction between ammonia (nonionized) and ammonium
(a cation):
NH
4
þ
, NH
3
þ H
þ
ð13:9Þ
When speaking of the ammonia-N concentration in water, it is usually the
total ammonia-N, or NH
4
þ
-N þ NH
3
-N that is meant, as this is what is measured
by all of the commonly used analytical methods. The pK
a
value (log K
a
)at25
C
for this reaction, at which half of the total ammonia would be in each form, is
about 9.3:
K
a
¼
½NH
3
½H
þ
½NH
4
þ
¼ 10
9:3
Since acid–base reactions are fairly rapid, at neut ral pH values, only a smal l per-
centage of the total ammonia is expected to be present as NH
3
(0.50% at pH 7,
4.8% at pH 8).
416 MICROBIAL TRANSFORMATIONS

Example 13.4 The pH of a water sample at 25
C is 7.5. If the measured total ammonia-
N concentration is 3.0 mg/L, what is the concentration of nonionized NH
3
-N?
Answer Let ½N
T
¼ NH
3
½þNH
þ
4
; then
K
a
¼
½NH
3
½H
þ
NH
4
þ
½
¼
NH
3
½H
þ
½
N
T
½NH
3
½
NH
3
½¼N
T
½
K
a
H
þ
½þK
a
NH
3
-N ¼ N
T
K
a
H
þ
½þK
a
¼ N
T
10
pK
a
10
pH
þ 10
pK
a
¼ N
T
1
10
pK
a
pH
þ 1
¼ 3:0
1
10
9:37:5
þ 1
¼
3
10
1:8
þ 1
¼ 0:047 mg=L
13.2.1 Nitrogen Reduction
Important nitrogen reduction reactions include nitrogen fixation, dissimilatory nitrate
reduction (including denitrification), and nitrate assimilation.
Nitrogen Fixation Although a gaseous sea of N
2
blankets Earth, it offers nothing in the
way of nutritional value except to a select few prokaryotes. The trivalent bond (N
N) of
this molecule is simply too strong for normal metabolic cleavage. Indeed, one of the
names that Antonie Lavoisier originally considered in the late eighteenth century for
this recently discovered gas was ‘‘azote,’’ meaning lifeless or inert.
Since the vast majority of life-forms are unable to utilize nitrogen gas, a critical step in
the global cycling of nitrogen is its fixation. Nitrogen fixation is the reduction of N
2
to
organic or ammonia nitrogen. As indicated in Figure 13.18, some nitrogen is also fixed
abiotically in natural systems through oxidation reactions involving combustion, photoly-
sis, or electricity (lightning).
A variety of nitrogen fixers are known, scattered among a number of taxa, but all are
prokaryotes. They are often separated into free-living vs. symbiotic forms, although this is
not a phylogenetic approach. Although Azotobacter and Beijerinckia are probably the best
known of the free-living, nitrogen-fixing aerobes among the proteobacteria, other exam-
ples are found among species of Klebsiella (but only when growing under anaerobic con-
ditions) and Citrobacter (members of the Enterobacteriaceae family), Methylomonas (a
methanotroph), and Thiobacillus (a sulfur-oxidizing autotroph). There are also aerobic
gram positives (e.g., a few Bacillus and the actinomycete Streptomyces) and many cyano-
bacteria, such as Anabaena (see Figure 10.20) and Nostoc. Free-living anaerobic nitrogen
fixers include members of the gram-positive spore-formers Clostridium and Desulfotoma-
culum (a sulfate-reducer); the sulfate-reducing proteobacteria Desulfovibrio; several
phototrophs (Table 10.4), such as Chlorobium (green sulfur bacteria), Chr omatium (purple
sulfur), Rhodospirillum (purple nonsulfur), and Heliobacterium (another gram-positive
spore-former); and a few methanogens, such as Methanococcus, which are archaea.
The best known symbiotic nitrogen fixers are rhizobia, proteobacteria such as Rhizo-
bium associated with leguminous plants (e.g., beans, peas, soybeans, peanuts, alfalfa, and
clover). Within the rhizospere (root zone in the soil), these bacteria infect the roots of the
NITROGEN 417