Vaccari D.A., Strom P.F., Alleman J.E. Environmental Biology for Engineers and Scientists
Подождите немного. Документ загружается.

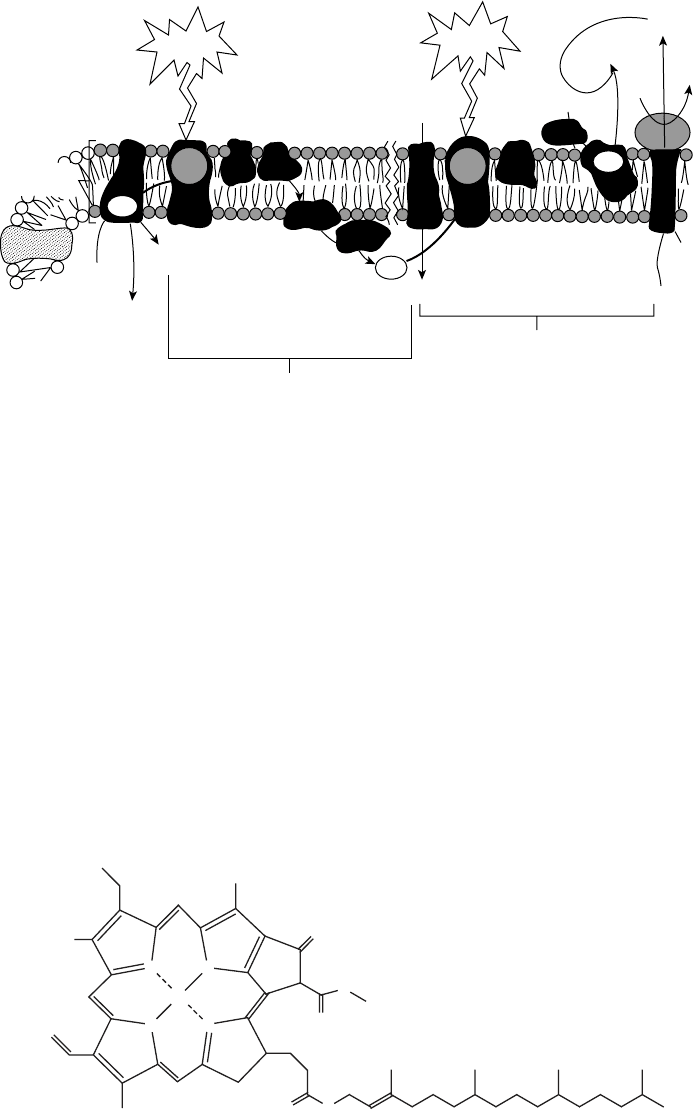
of chlorophyll molecules, called the reaction center, which uses the energy to promote an
electron to a higher energy level and transfer it to the electron transport system. The reac-
tions centers of photosystem I and photosystem II are known as P700 and P680, respec-
tively, for the optimum wavelengths at which they operate.
The chlorophyll molecule consists of a hydrocarbon tail connected to a ring system
with a nonionic magnesium atom at its center (Figure 5.10). The ring system has the alter-
nating single and double bonds that often characterize compounds that absorb visible
light. There are two main types of chlorophyll: a and b, but chlorophyll a is most common
in plants. Chlorophyll a absorbs most strongly in the blue (400 to 450 nm) and red (640 to
680 nm) regions of the spectrum, leaving green to please our eyes (Figure 5.11). However,
photosystem antennas contain other pigments, such as carotenoids, which capture green
and yellow light energy and transfer it to the chlorophyll. This is particularly important for
plants that live in deep waters, where the longer wavelengths do not penetrate. Diatoms,
the brown algae, and dinoflagellates (including the red tide organism) contain pigments
that absorb strongly in the range 400 to 550 nm. The carotenoids become visible when the
Photosystem II
Photosystem I
ATPase
H
+
H
+
H
+
high [H
+
]
2H
+
H
2
O
½O
2
Thylakoid
membrane
P680
P700
2e
2e
AT P
ADP
NADP
NADPH
CF
1
Photon
Photon
Stroma
(low [H
+
])
2e
P700
Figure 5.9 Photosynthetic apparatus. (From Fried, 1990. # The McGraw-Hill Companies, Inc.
Used with permission.)
Mg
N
NN
N
R
O
O
O
OO
Chlorophyll a : R = CH
3
Chlorophyll b : R = CHO
Figure 5.10 Structure of chlorophyll a and chlorophyll b.
108
ENERGY AND METABOLISM
chlorophyll degrades in deciduous tree leaves in certain areas in the fall, creating the spec-
tacular fall color displays.
Before examining photosynthesis in plants, it may be useful to consider bacterial
photosynthesis first, because it is simpler and because its mechanism is retained in
algae and plants. Bacteria have only one photosystem. Absorption of a photon by the
photosystem starts the process of cyclic photophosphorylati on, which results in the pro-
duction of a single ATP. The photon excites an electron in the chlorophyll, which transfers
to the electron transport chain. As the electron is passed down the chain, two protons are
secreted outside the cell using energy from the electron. At the end of the chain, the low-
energy electron returns to the chlorophyll, ready to start the cycle again. The proton secre-
tion creates a chemiosmotic potential across the membrane, which produces ATP as it
passes back into the cell through an enzyme complex. To form carbohydrates, the bacteria
need ATP and NADPH
2
. Bacteria require an external chemical reducing agent to form
NADPH
2
, such as H
2
,H
2
S, or organic matter. The ATP and NADPH
2
then feed the
dark reactions to produce carbohydrates as described below. No oxygen is produced as
it is by plants. Indeed, oxygen is toxic to these bacteria. An exception is the prokaryotic
nitrogen-fixing cyanobacter, which does produce oxygen.
Light Reactions Plants and algae retain the cyclic photophosphorylation pathway but
have developed noncyclic photophosphorylation , which has the advantage of not requir-
ing an external reducing agent to replace the electron given up by the chlorophyll
(Figure 5.12). Instead, in photosystem II, water is hydrolyzed to hydrogen and oxygen;
the hydrogens provide the needed electrons, and the remaining protons enter the thylakoid
lumen to augment the chemiosmotic gradient (ultimately for ATP production). The oxy-
gen formed by hydrolysis of the water is released. After passing through the electron
transport chain of photosystem II, using their energy to pump more protons, the low-
energy electrons can then replace electrons promoted by another photon absorbed by
photosystem I. The reenergized elect rons, now in photosystem I, pass through another
electron transport chain and ultimately are used to reduce NADP to NADPH
2
. (Note
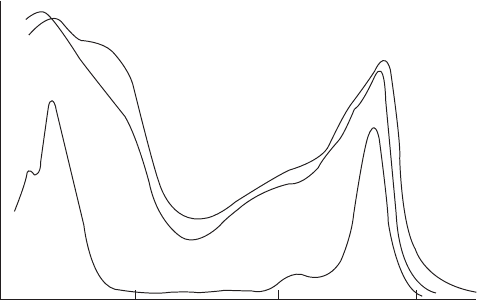
400 500 600 700
Wavelen
g
th (nm)
Action
spectrum
Absorption
spectrum
Chlorophyll a
Relative light absorption
Relative photochemical efficiency
Figure 5.11 Absorption spectrum of chlorophyll and of a green leaf, and the action spectrum for
the rate of photosynthesis vs. wavelength. (From Fried, 1990. # The McGraw-Hill Companies, Inc.
Used with permission.)
BIOCHEMICAL PATHWAYS 109

the use of NADP instead of NAD as in respiration. NADP is used preferentially when the
reducing power is to be used for synthesizing compounds, not just for transporting elec-
trons.) The sequence of events takes about 5 ms. The overall reaction for noncyclic photo-
phosphorylation is
12 H
2
O þ 48 hn þ 12 NADP þ 12 ADP þ 12 P
i
) 6O
2
þ 12 NADPH
2
þ 12 ATP
ð5:55Þ
Equation (5.55), as written, involves two electrons, each of which requires two
photons. The protons produced by photolysis of water, plus additional protons pumped
by the electron transport system (not shown), wind up in the thylakoid lumen. The che-
miosmotic gradient across the thylakoid membrane can be as much as 3 pH units, with an
electric potential of up to 100 mV. About one ATP is produced per NADPH
2
. Overall,
then, noncyclic photophosphorylation requires four photons to produce one ATP and
one NADPH
2
.
Noncyclic photophosphorylation produces NADPH
2
and ATP in about equal amounts;
but glucose production requires additional ATP, and the cell has othe r uses for ATP as
well. To get more ATP, plants are able to exploit cyclic photophosphoryla tion, similar
to bacteria, which does not produce oxygen or NADPH
2
. It does this by a sort of
‘‘short circuit’’ that shunt s electrons excited by photosystem I back to the electron trans-
port chain of photosystem II (see the dotted line in Figure 5.12). The process is controlled
by NADPH
2
levels; high levels hinder the flow of electrons out of photosystem I, forcing
them into the other pathway. Thus, the cell can control somewhat independently the rela-
tive amounts of NADPH
2
and ATP that it produces.
The energy of a mole of photons, E, is related to the frequency, n (or wavelength, l)by
E ¼ hn ¼
hc
l
ð5:56Þ
where h is Planck’s const ant and c is the speed of light. The two peak wavelengths shown
in Figure 5.11 are 430 and 650 nm. The corresponding energy levels are 67 and 43.5 kcal/
mol, respectively. Four moles of photons of light energy must be absorbed by each of the
−800
−600
−400
−200
0
200
400
600
800
1000
1200
1400
1600
Standard electrode potential (mV)
Photosystem
I
Photosystem
II
P700
P680
Z
ATP
Cyclic photophosphorylation
P430
Cyt b563
Two
photons
Two
photons
H
2
O
2H+
1/2O
2
2e-
2e-
NADP
NADPH
ADP
Figure 5.12 Noncyclic photophosphorylation.
110
ENERGY AND METABOLISM
two photosystems (eight photons in all) to produce one molecule of O
2
, two of NADPH
2
,
and two of ATP. To produce a six-carbon sugar by the dark reac tions, the cell will need
12 NADPH
2
and 18 ATP. This can be satisfied most efficiently by 48 photons in noncyclic
photophosphorylation (making 12NADPH
2
and 12ATP) and 12 photons in cyclic photo-
phosphorylation (six more ATP), for a total of 60 photons. Assuming that a conservative
40 kcal/mol of photons gives a total of 2400 kcal/mol glucose formed since the standard
Gibbs free energy of formation of glucose is 686 kcal/mol, the potential efficiency is 29%
(based on photons absorbed). The actual efficiency is less, due largely to a wasteful side
reaction called photorespiration, described below.
Dark Reactions The conversion of CO
2
to glucose is calle d CO
2
fixation. It is accom-
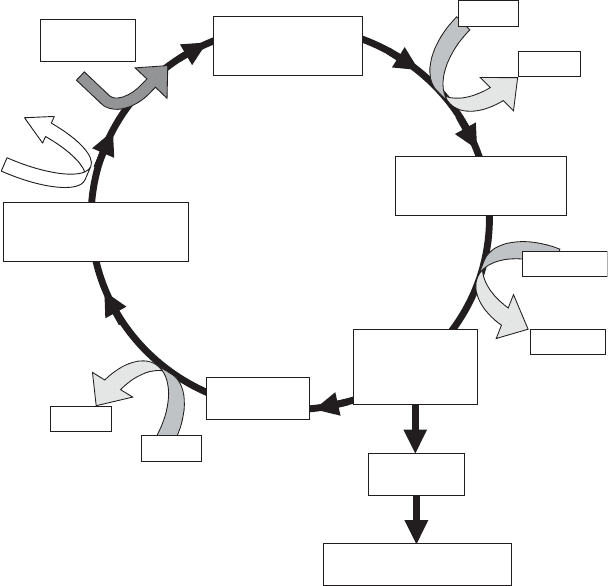
plished by two pathways. The first is a series of 12 reactions called the Calvin cycle
(Figure 5.13). The cycle starts in the stroma of the chloroplast when CO
2
combines
with a pentose (five-carbon sugar) to form two three-carbon acids:
ribulose-1;5 biphosphate þ CO
2
) 2;3-phosphoglycerate ð5:57Þ
Note that this step may be considered to be the actual point of carb on fixation. Yet
it does not require energy from light or even ATP. But ATP and the reducing agent
NADPH
2
are needed to ensure a supply of the ribulose to keep this reaction going. Several
6 Molecules of
Ribulose 1,5-diphosphate
(C-5)
12 Molecules of
3-Phosphoglycerate
(C-3)
6 Molecules
of CO
2
12 Molecules of
1,3-diphosphoglycerate
(C-3)
12ATP
12ADP
12 NADPH
12 NADP
The Calvin Cycle
12 Molecules of
Glyceraldehyde
3-phosphate
(PGAL) (C-3)
12ATP
12ADP
10 molecules
PGAL
2 molecules
PGAL
Carbohydrate Synthesis
(glucose, sucrose, starch)
O
2
(Photorespiration)
Figure 5.13 Carbon dioxide fixation.
BIOCHEMICAL PATHWAYS 111
subsequent reac tions use two NADPH
2
and two ATP to produce phosphoglyceraldehyde
(PGAL, or glyceraldehyde-3-phosphate). Most of the PGAL goes on to be converted back
into the ribulose, to keep the cycle go ing. This requires another ATP.
Two of every 12 PGALs formed are converted to another triose, which leaves the chlor-
oplast for the cytoplasm. There they can be converted to glucose or other carbohydrates in
the second pathway. The enzymes for forming glucose are the same as the ones in glyco-
lysis, acting in reverse. In fact, PGAL is one of the key intermediates in glycolysis. PGAL
can also be converted into glycerol and fatty acids for fats, or into amino acids for pro-
teins. Overall, since each turn of the cycle incorporates a single CO
2
, it takes six cycles to
produce one molecule of glucose. The overall stoichiometry for the Calvin cycle is
6CO
2
þ 12 NADPH
2
þ 18 ATP ) C
6
H
12
O
6
þ 12 NADP þ 18 ADP þ18 P
i
þ 6H
2
O
ð5:58Þ
Strangely, most plants have a wasteful side reaction, called photorespiration, which
uses the same enzyme as reaction (5.57). The reaction occurs during hot, dry conditions
when leaf pores close to conserve water. CO
2
is depleted inside the leaf and O
2
builds up.
The oxygen competes with the CO
2
for the enzyme, producing a side reaction with the
ribulose-1,5 biphosphate. Waste products are formed instead of carbohydrates, and ATP is
consumed. There seems to be no benefit to the plant. This is a major drain on biological
productivity for the world’s plants and can reduce the net efficiency of photosynthesis
below 1%.
A minority of plants (about 100 species are known) have developed a mechanism to tilt
the balance back toward normal CO
2
fixation. This has occurred in plants that grow in hot,
arid regions. Closing the stomates to limit water loss also limits CO
2
entry to the leaf. In
these plants, instea d of forming the three-carbon phosphoglycerate in reaction (5.57), cells
near the outside of the leaf use a different reaction that forms the four-carbon oxaloace-
tate, which is then reduced by NADPH
2
to the four-carbon malate. The malate diffuses to
other cells in the leaf interior, where it reacts to form pyruvate, CO
2
, and NADPH
2
.This
has the net effect of transporting CO
2
and NADPH
2
to the inner cells at a higher concen-
tration, where they enter the Calvin cycle. Twelve additional ATPs are used per glucose,
but the net efficiency increases because photorespiration is relatively low. Plants that do
this include the important food crops sorghum, corn, and sugarcane. Because they form a
four-carbon product with CO
2
, they are called C
4
plants. The more common plants with-
out this ability are called C
3
plants.
Actual maximum photosynthetic efficiencies of the C
4
crops sugarcane (Saccharum
officinale), sorghum (Sorghum vulgare), and corn (Zea mays) have been measured to
range from 2.5 to 3.2%; for the C
3
crops alfalfa, sugar beet, and the alga Chlorella, the
range is 1.4 to 1.9%. C
3
plants can fix 15 to 40 mg of CO
2
per dm
2
of leaf surface per
hour, while C
4
plants can fix 40 to 80 mg/dm
2
per hour. C
4
plants lose much less water by
transpiration than do C
3
plants. The optimum daytime temperature for growth of C
4
plants
is higher: 30
to 35
C vs. 20
to 25
C for C
3
plants. Examples of C
3
crop grasses are
wheat (Triticum aestivum), rye (Secale cereale), oats (Avena sativa), and rice (Oryza
sativa).
A third strategy for photosynthesis is carried out by succulents such as the jade
plant and some cacti. These are called CAM (for ‘‘crassulacean acid metabolism’’)
plants. CAM plants fix carbon at night with the stomata (pores in the leaves) open. During
the heat of the day the stomata close, and the Calvin cycle obtains its carbon from the
112 ENERGY AND METABOLISM
nighttime storage. Note that C
4
and CAM plants have all the photosynthetic machinery of
C
3
plants: namely, the photosystems and the Calvin cycle. However, they have additional
mechanisms that enable them to utilize CO
2
more efficiently.
The difference between C
3
and C
4
plants has caused concern about the effect of the
global CO
2
increase that is under way. Fossil fuel consumption has caused CO
2
in the
atmosphere to increase from 280 ppmv in preindustrial times to 356 ppmv in 1993. C
3
plants are more sensitive to low CO
2
levels. Experimental results have verified that
they can benefit more from an increase than C
4
plants. This has the potential to cause
ecological changes as the competitive balance between plant species changes. It is also
feared that some of the C
4
food plants could face increased competition from C
3
weeds. On the other hand, the lawn grasses Kentucky Bluegrass (Poa pratensis) and
creeping bent (Agrostis tenuis) are C
3
plants, whereas crabgrass (Digitaria sanguinalis)
is a C
4
plant, ordinarily giving it an advantage during hot, dry summers.
5.4.6 Biosynthesis
Many of the smaller molecules of the cell, whether those used directly or those used as
building blocks of macromolecules, are synthesized from precursors found among the
intermediates of respiration. Th ese anabolic pathways connect respiratory intermediates
with other sugars, amino acids, fatty acids and fats, and nucleic acids. Virtually all the
pathways in a cell are interconnected in a network of reactions.
Nitrogen is assimilated by cells by forming glutamat e and glutamine from ammonia
and ketoglutaric acid. In nitrogen-poor environments, energy must be expended. Rooted
plants and algae can use ammonia, but primarily take nitrogen in as nitrate. Some micro-
organisms can take in nitrate or fix atmospheric nitrogen, but first convert these to ammo-
nia. Other amino acids are then formed using the glutamate, possibly with other
precursors, many of which are intermediates in respiration, such as pyruvate or oxaloace-
tate. Formation of proteins from amino acids is discussed in Chapter 6.
Nucleotide synthesis begins with ribose for purines and glutamin e for pyrimidines. The
nitrogens are obtained from the amino acids glycine, glutamine, and aspartate. Fatty acids
are formed using acetyl-CoA, similar to a reverse of the fatty acid oxidation described
above. The process occurs in the cytoplasm and requires NADPH
2
,CO
2
, and Mn
2þ
.
Mammals can synthesize the saturated and monounsaturated fatty acids.
The biopolymers (starch, glycogen, protein, and nucleic acids) are produced by succes-
sive addition to the ends of the molecules. Often, ATP is converted to another nucleoside
triphosphate for the reaction, such as uridine triphosphate, cytosine triphosphate, guani-
dine triphosphate, or thymine triphosphate. The reaction usually splits the second phos-
phate bond, producing a monophosphate instead of a diphosphate. For example, if a
monomer M is joined to a polymer of n units,
ATP þ M þ½M
n
) AMP þ½M
nþ1
þ PP
i
Then the resulting pyrophosphate (PP
i
) is hydrolyzed to orthophosphate. By eliminating
the pyrophosphate reaction product, the first reaction is pushed toward completion by the
mass action law.
Sulfur in the form of sulfide (S
2
) is needed for the amino acid cysteine, which forms
the disulfide bonds that are so important to protein folding. However, our aerobic envir-
onment provides sulfur mostly in the form of sulfate, SO
2
4
. The sulfide can be formed
BIOCHEMICAL PATHWAYS 113
anaerobically by microorganisms that use the sulfate as a terminal electron acceptor in the
electron transport chain. Sulfate can also be reduced by a series of steps starting with ATP,
followed by use of one NADPH
2
to form sulfite ðSO
2
3
Þ and then three more NADPH
2
to
reduce the sulfite to sulfide.
PROBLEMS
5.1. Compute G
and K
eq
for the complete oxidation of ethanol.
5.2. Compute G
and K
eq
for methanogenesis C
6
H
12
O
6
, 3CO
2
þ 3CH
4
.
5.3. Compute the oxidation states of glucose, pyruvate, and lactic acid. Is the conversion
of glucose into two pyruvates an oxidation? How about the conversion of glucose
into lactate?
5.4. For pyruvic acid, glucose, ethanol, and glycine: Using the molecular formula,
compute the oxidation state, the theoretical oxygen demand, and the total organic
carbon content (TOC). Use equation (5.9) to compute the mean oxidation state of
carbon. Compare with the value computed directly from the formula. Discus s the
results.
5.5. Use the Arrhenius equation to compare the temperature increase that doubles the
reaction rate coefficient starting from 25
C for the hydrolysis of gluc ose example
given in the text, for cases with and without enzyme catalysis.
5.6. In Example 5.4, at what concentration will the reaction rate be double the first value
measured?
5.7. (a) Write a computer program or develop a spreadsheet to solve Michaelis–Menten
equations (5.30) to (5.32) numerically using Euler’s rule with t ¼ 0:001 min. Start
by writing a differential equation based on elementary kinetics for d½P=dt based on
equation (5.29). Use values for all rate coefficients equal to 1.0 (units are min
1
or
minL/mg), initial values are [S]
0
¼100, and [E]
0
¼50.0, and [ES] ¼ [P]
0
¼ 10.
Plot the values of [S], [ES], and [P] from 0.0 to 0.5 min. How accurate is the
quasisteady-state assumpt ion?
(b) Repeat part (a) using [E]
0
¼100. How is the rate of conversion of S to P
affected?
5.8. Using the data of Figure 5.4, equation (5.47) gives reaction rates of 145.2, 185.8,
and 234.2 mm
3
/min for temperatures 10, 20, and 30
C, respectively. Use the data
for 20 and 30
C in equation (5.48) and solve for y. Then use (5.48) with your value
of y to estimate the reaction rate at 10
C. How does this compare to the value from
equation (5.47)?
5.9. What is the difference between Le Cha
ˆ
telier’s principle and the law of mass action?
5.10. If a first-order reaction has a half-life of 1 h, what is the rate coefficient for that
reaction? How long will it take for the reactant to fall to 1% of its initial
concentration?
5.11. If K
m
in the Michaelis–Menten equation is equal to 0.02 M, at what concentration
will the reaction proceed at 90% of the maximum rate? 99%?
114 ENERGY AND METABOLISM
5.12. A sample of water containing photosynthetic algae (and negligible amounts of
heterotrophs) is placed in a sealed bottle in sunlight. After a time it is observed that
the dissolved oxygen level in the water has increased and dissolved carbon dioxide
decreases. How will the rate of change of these two substances change immediately
after the bottle is placed in the dark? (Don’t forget about the dark reactions of
photosynthesis.) What about several hours after being in the dark?
REFERENCES
Bailey, J. E., and D. F. Ollis, 1986. Biochemical Engineering Fundamentals, McGraw-Hill, New York.
Fried, G. H., 1990. Schaum’s Outline: Theory and Problems of Biology, McGraw-Hill, New York.
Grady, C. P., Jr., G. T. Daigger, and H. C. Lim, 1999. Biological Wastewater Treatment, Marcel
Dekker, New York.
Henze, et al., 1986. Activated Sludge Model No. 1, International l Association on Water, London.
Lange’s Handbook of Chemistry, 1987. McGraw-Hill, New York.
Lehninger Principles of Biochemistry, 3rd ed., W.H. Freeman, New York.
Smith, E. L., R. L. Hill, I. R. Lehman, R. J. Lefkowitz, P. Handler, and A. White, 1983. Principles of
Biochemistry: General Aspects, McGraw-Hill, New York.
Stryer, L., 1995. Biochemistry, 4th ed., W.H. Freeman, New York.
Vogel, F., J. Harf, A. Hug, and P. R. von Rohr, 2000. The mean oxidation number of carbon (MOC): a
useful concept for describing oxidation processes, Water Research, Vol. 34, No. 10, pp. 2689–
2702.
Watson, J. D., 1968. The Double Helix, Atheneum, New York.
Weast, R. C. (Ed.), 1979. CRC Handbook of Chemistry and Physics, 60th ed., CRC Press, Boca Raton,
FL.
REFERENCES 115

6
GENETICS
Genetics is one of the most fascinating areas of biology. It has effects at all scales from the
molecule to populations. Its study involves a wide variety of tools, from biochemical tests
to microscopy to breeding experiments. It relies on sophisticated mathematical analysis,
especially from probability and statistics. It is one of nature’s best examples of amplifica-
tion, in which a small change in a single molecule can spell the difference between
disaster and delight at the birth of a child.
Genetics is the science of heredity. Heredity is the transmission of traits from one
generation to another. By traits, we mean a distinguishing observable characteristic of
an organism, such as color or shape of one of its parts, or its ability to metabolize a
particular substance. Each trait is controlled by one or more genes.
6.1 HEREDITY
Gregor Mendel was one of the first biologists to use mathematical analysis to prove
hypotheses. He used elementary probability theory in his work. He was also inspired
by the atomic theory of matter (an appropriate example of cross-fertilization in science).
He wondered whether hereditary traits were also passed as particles to future generations.
Previously, it was thought that traits from parents blended to form a new mix in offspring.
Mendel proved his hypothesis and published his results starting in 1865. However, his
contribution was not recognized until the principles were rediscovered inde pendently
by others some 35 years later.
Mendel deduced several of the basic principles of heredity. Here we summarize all of
the principles as known today. Then several of Mendel’s and others’ results will be
described as examples. The principles of heredity can be summarized as eleven rules:
Environmental Biology for Engineers and Scientists, by David A. Vaccari, Peter F. Strom, and James E. Alleman
Copyright # 2006 John Wiley & Sons, Inc.
116
1. A gene is a sequence of nucleotides that codes for a specific polypeptide. [Recall
that a protein can consist of several polypeptides, and possibly other factors such as
metal atoms (e.g., hemoglobin, which has four polyp eptides and iron atoms).] Not
all genes in a particular cell actually produce their polypeptide. Cells that do
produce the polypeptide associated with a gene are said to express the gene.
2. The genes are contained in the chromosomes.
3. A gene may come in a variety of mutated forms, called alleles. For example, the
gene for eye color may have an allele for blue eyes and another allele for brown
eyes. Although all the organisms of a single species will have the same genes, each
individual can have a unique combination of alleles.
4. The particular combination of alleles that an individual has is called its genotype.
The particular set of genetically controlled traits possessed by an individual is
called its phenotype. That is, the phenotype is the result of expression of the
genotype.
5. Most higher organisms, in particular most plants and animals, are diploid, having
chromosomes in pairs. Thus, they have two copies of each gene.
6. The two copies can have the same or different alleles. If both chromosomes have
the same allele, the organism is said to be homozygous for that gene. If the alleles
are different, the organism is heterozygous. For example, a human can have a blue-
eyed allele on one chromosome and a brown-eyed allele on the other. That
individual would be heterozygous for eye color. Alternatively, both alleles could
be for blue eyes, and the person would be homozygous for eye color.
7. When the two alleles for a gene are different, several situations can occur:
a. Complete dominance: one allele is expressed; the other is not. An allele that is
expressed at the expense of another is said to be dominant. The allele that is not
expressed in a heterozygous genotype is said to be recessive. For example,
brown eye color is dominant and blue eye color is recessive. Blue eye color is
expressed only if both alleles are for blue eyes. When a gene is homozygous for
a recessive allele, it is said to be double recessive, and the recessive allele can be
expressed.
b. Incomplete, or partial, dominance : In this case, a mixed phenotype results in a
heterozygous individual. For example, if snapdragons with red flowers (homo-
zygous for red) are crossed with those with white flowers (homozygous for
white), all the offspring will be heterozygous and will have pink flowers.
However, if these are then crossbred with each other, about one-fourth of the
next generation will be red, one-fourth whi te, and one-half pink.
c. Codominance is when both traits are fully expressed in the heterozygous
organism. This is usually seen only in biochemical traits. An example is
human blood type. If a heterozygous individual has allele A and allele B,
their blood cells will have both type A and type B antigens.
8. Meiosis separates the two alleles of a diploid organism. Thus, gametes have only
one allele. This is called segregati on .
9. Random assortment in meiosis distributes the chromosomes to the gametes
independently. That is, the chromosomes of an individual’s gametes can have
any combination of its maternal and paternal chromosomes. In genetics, this is
HEREDITY 117