Vaccari D.A., Strom P.F., Alleman J.E. Environmental Biology for Engineers and Scientists
Подождите немного. Документ загружается.


transport, which requires energy. Water transport, on the other hand, is mostly passive,
powered by evaporation in the leaves.
The leaf is the main photosynthetic organ of the plant. Its large surface area is designed
to capture sunlight. A cross section through the leaf (Figure 7.4) shows that the upper and
lower surfaces of the leaf are formed of a layer of cells called the epidermis, which is
covered by a waxy cuticle to minimize water loss. The cells between these two layers
form the mesophyll, which has two parts. The palisade parenchyma form vertical col-
umns under the upper surface. They are the main photosynthetic tissues of the leaf. The
rest of the mesophyll consists of spongy parenchyma, which has a about 15 to 40% void
volume filled with air and a large surface area for exchange of carbon dioxide, oxygen,
and water vapor.
The epidermis is punctuated by special pores called stomata (singular, stoma or
stomate). There can be as many as 12,000 stomata per square centimeter, mostly on
the lower surface of the leaf, where they occupy about 1% of the leaf surface. Each
stoma is made up of a pair of banana-shaped guard cells. The guard cells have chloro-
plasts, whereas the other epidermal cells do not. The stomata control the passage of gases
into and out of the leaf, thus affecting photosynthesis and water use. They also affect the
transport of pollutants into the leaf. When the guard cells accumulate water, they swell,
causing them to bow outward, opening the pore. Conversely, loss of water causes the pore
to close. This occurs in response to several conditions. First, and most obviously, if water
is scarce, a general wilting will cause the stomates to close, limiting water loss. This gives
the plant control over transpiration , the evaporation of water from the plant. This ‘‘loss’’
is actually necessary for the plant’s survival, giving it the means to transport minerals
Figure 7.3 Microscopic view of root with root hairs. (From Simpson and Orgazaly, 1995.)
148
THE PLANTS
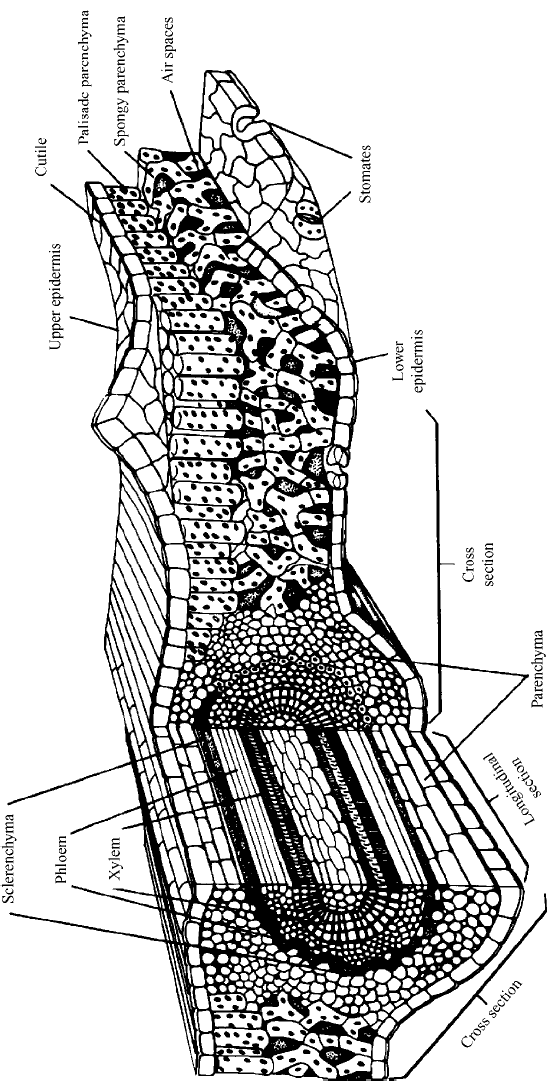
Figure 7.4 Cross section of a leaf, showing layers, a vein, and a stoma. (From Simpson and Orgazaly, 1995.)
149
from the roots. A field of corn in Kansas transpires the equivalent of 28 cm of rain per
year. A stand of red maple transpires an estimated 72 cm. These figures are far in excess
of the water actually used by the plant in its tissues or for photosynthesis.
When water is not limiting, the plant needs the stomata to open to admit CO
2
for
photosynthesis. When light is present, the plant actively transports salts into the guard
cells. This increases their osmotic potential, drawing in water and opening the pore.
Photosynthesis in the guard cells also contributes to their osmotic potential by producing
glucose in the cytoplasm. Temperatures above 30 to 35
C or high CO
2
levels can cause
stoma closing. Because of all these effects, a plant may close its stomata at night, open
them in the morning, then close them again in the heat of the midday, finally to open them
again when the afternoon cools. The arid-climate CAM plants are different: They
open their stomata at night and close them all day to conserve water. CO
2
is stored during
the night for use in photo synthesis d uring the day.
An interesting problem for tall plants is how they are able to move water from the soil
to great heights, as high as 111 m. Either the water is pushed by the roots, in which case
the water column would be under pressure, or it is drawn from the top, which would place
the water in the xylem under vacuum. Experiments show that when the xylem is punctu-
red, air is drawn in. This eliminates the pressure hypothesis. However, it is a simple
hydrodynamic fact that even a perfect vacuum pump cannot raise water much more
than about 10 m. The accepted explanation is called the cohesion-tension theory.
The suction is provided by evaporation of water from small pores in the leaves. The
water column is prevented from breaking because of adhesion to the walls of the
xylem. Experiments have shown that water in a fine capillary can withstand a tension
of up to 26.4 megapascal (MPa). But only 2.0 MPa (about 20 atm) is needed to
lift water to the top of a tall redwood. (Actually, the roots of many plants do produce
some pressure. Th ey do so by active tran sport of ions into the xylem, producing osmotic
pressure. Root pressures as high as 0.3 to 0.5 MPa have been measured.)
Transport of sugars via the phloem is described by the pressure-flow hypothesis. First,
the sugars (mostly sucrose) are moved into the phloem from leaves or storage by active
transport. This increases the osmotic potential, causing water to be absorbed into the
phloem, which generates hydrostatic pressure. The pressure causes the fluid to flow
toward tissues that are removing sugars. The removal of sugar at those sites causes
water to leave the phloem osmotically at that point. Most of the water then returns to
the xylem to complete the circulation. Thus, plants, like animals, can be thought of as
having a complete circulatory system, although it is not a single closed system.
7.2.2 Plant Growth and Control
Plants produce a number of chemical messengers, called hormones, which affect the way
the plant grows. There are five main hormones or groups of hormones:
1. Gibberellins promote growth, especially the lengthening of stems and formation of
fruit and leaves.
2. Auxins promote growth by stimulating cells to elongate. They control the bending
of plants toward light. They are used commercially to stimulate plant cuttings to
form roots. Auxins are produced by the growing tip of the plan t and diffuse
downward through the plant. Pruning encourages the formation of lateral buds by
150 THE PLANTS
removing the auxin-forming tip of the plant. Developing seeds produce auxin, which
stimulates the formation of the fruit. There is only one naturally occurring auxin:
indole-3-acetic acid, which is chemically similar to the amino acid tryptophan.
However, many synthetic analogs have been developed, including the herbicides
2,4-D and 2,4,5-T.
3. Cytokinins promote growth by stimulating cell division. They have an effect on
differentiation of plan t cells. Leaves that turn yellow after being picked can be kept
green longer by treatment with cytokinins. Chemically, they are derivatives of
adenine.
4. Abscisic acid has the opposite effect of some of the other hormones: It inhibits
growth and development. It is produced during water stress, causing the stomata to
close and thereby inhibiting photosynthesis. It is involved in causing the seed
embryo to become dormant so that it does not germinate prematurely. It also
stimulates. A some processes, such as protein storage in seeds.
5. Ethylene is the simple hydrocarbon H
2
C
CH
2
. It stimulates maturation, promoting
the ripening of fruit and the dropping of fruit, leaves, and flowers. Being a gas, it is
released to the air by plants and ripening fruit. This explains the adage ‘‘one bad
apple spoils the bunch,’’ as ethylene from o ne fruit hast ens ripening in the others.
Plants also release it when injured. Ethylene is used commercially to stimulate
ripening of fruit such as the tomato that were picked green so that they could be
transported to market while firm and less liable to damage. Ethylene is act ive at air
concentrations as low as 0.06 ppmv (parts per million by volume).
Plants also change their growth patterns in response to environmental stimuli, such as
by light and gravity. The response to gravity is called geotropism. If a potted plant is
placed on its side, the cells on the lower side of the stem will elongate, causing the
stem to bend upward. Auxin seems to be involved. The turning of plants toward the
light is called phototropism. Experiments have established that light coming from
the side, especially blue light, causes auxin to migrate to the shadow side of a stem.
This causes elongation on the shaded side, bending the plant toward the light.
Plants exhibit another behavior, familiar to those who raise houseplants, cal led photo-
periodism, in which the length of the night controls when the plant flowers. Plants exhibit
one of three photoperiodism behaviors: Short-day plants set flower only when the length
of the night exceeds a cri tical period, which varies from plant to plant. Examples include
some chrysanthemums, poinsettias, and strawberries. Short-day plants flower in the early
spring or fall. Ragweed, for example, blooms in the fall and needs at least 9.5 hours of
continuous darkness. Interestingly, a single flash of light in the middle of the night can
fool a short-day plant and prevent its flowering. Long-day plants require a period of dark-
ness less than some critical value and tend to flower in the summer. Sp inach, lettuce, and
some varieties of potato and wheat are long-day plants. Spina ch will bloom only if there
is less than 10 hours of darkness. In contrast to short-day plants, a flash of light during a
long night can fool a long-day plant into flowering. Finally, there are day-neutral plants,
such as cucumber, sunflower, rice, and corn, which are not controlled by photoperiod.
Photoperiodism is controlled by a membrane- bound protein complex called phytochrome
that acts as a detector for light. Plants also use phytochrome to detect if light is totally
absent, such as if the plant is shaded by other plants or by a fallen log. In response, plants
do not produce chlorophyll but instead, devote their energy to growing longer, seeking
STRUCTURE AND PHYSIOLOGY OF ANGIOSPERMS 151

TABLE 7.2 Elemental Plant Nutrients
Form in Which Typical Plant Concentration
Element Absorbed (% or ppm dry weight) Functions
Macronutrients
Carbon CO
2
44% Component of organic compounds
Oxygen H
2
OorO
2
44% Component of organic compounds
Hydrogen H
2
O 6% Component of organic compounds
Nitrogen NO
3
or NH
4
þ
1–4% Component of amino acids, proteins, nucleotides, chlorophylls, and coenzymes
Potassium K
þ
0.5–6% Involved in osmosis and ionic balance, stomata opening and closing, and coenzymes
Calcium Ca
2þ
0.2–3.5% Cell wall component; enzyme cofactor, involved in membrane permeability
Phosphorus H
2
PO
4
or HPO
4
2
0.1–0.8% Component of energy-storing compounds (ATP), nucleic acids, several essential
coenzymes, phospholipids
Magnesium Mg
2þ
0.1–0.8% Part of the chlorophyll molecule; activator of many enzymes
Sulfur SO
4
2
0.05–1% Component of some amino acids and coenzyme A
Micronutrients
Iron Fe
2þ
or Fe
3þ
25–300 ppm Required for chlorophyll synthesis; component of cytochromes and nitrogenase
Chlorine Cl
100–10,000 ppm Involved in osmosis and ionic balance; probably essential in photosynthetic reactions
that produce oxygen
Copper Cu
2þ
4–30 ppm Activator or component of some enzymes
Manganese Mn
2þ
15–800 ppm Activator of some enzymes; required for integrity of chloroplast membrane and for
oxygen release in photosynthesis
Zinc Zn
2þ
15–100 ppm Activator or component of many enzymes
Molybdenum MoO
4
2
0.1–5.0 ppm Required for nitrogen fixation and nitrate reduction
Boron B(OH)
3
or B(OH)
4
5–75 ppm Influences Ca
2þ
utilization, nucleic acid synthesis, and membrane integrity
Essential to some
plants
Sodium Na
þ
Trace Involved in osmotic and ionic balance; probably not essential for many plants;
required by some desert and salt marsh species and may be required by all plants
that utilize the C
4
photosynthesis pathway
Cobalt Co
2þ
Trace Required by nitrogen-fixing microorganisms
Source: Raven et al. (1992), Table 27.2.
152
the sunlight. The result can be seen in the spindly, yellow growth of grass that has been
covered by a board for some days. Some seeds require total darkn ess in order to germi-
nate; others, such as lettuce, grow only if exposed to light. The difference relates to
whether the seeds need to be buried or on the surface of the soil to germinate. This beha-
vior is also controlled by phyto chromes.
Plants also show 24-hour cycles in movements of their leaves that persist even if light
cues to day length are removed. Some plants open their flowers in the morning; others
fold their leaves at night. In the absence of light cues, the timing is not precise. It ranges
from 21 to 27 hours. These cycles are called circadian rhythms and have been referred
to as biological clocks. Circadian rhythms are now known to occur in all eukaryotes,
including animals. However, they are absent in prokaryotes.
The herbicides 2,4-dichlorophenoxyaceticacid (2,4-D) and 2,4,5-trichlorophenoxyace-
tic (2,4,5- T) were active ingredients in Agent Orange, the dioxin-contaminated herbicide
used in the Vietnam war to deprive the enemy of their forest cover. The use and manu-
facture of 2,4,5-T is now banned in the United States. 2,4-D is still used in household
broadleaf weedkiller, because dicots (such as dandelion) are much more sensitive to it.
They act by causing the plant to grow out of control, plugging the phloem. Another impor-
tant herbicide is glyphosate, which is sold by Monsanto under the name Roundup.
Glyphosate effectively kills all plants, although it is nontoxic to animals. It acts by
blocking the action of a single enzyme involved in the production of aromatic amino
acids.
7.2.3 Plant Nutrition
Plants, being autotrophs, do not require organic food sources. In fact, they do not need any
organic growth factors. They can synthesize all of the amino acids and vitamins they
require. Thus, their only known nutritional requirements are the inorganic nutrients listed
in Table 7.2. Besides roles as components of organic compounds, the elements are used as
enzyme cofactors, as intermediates in electron-transfer reactions, and for regulating
osmotic pressure and membrane transport.
Nitrogen is absorbed by plants as either nitrate or ammonium. If it is in the form of
nitrate, it is reduced to ammonium by the plant. This requires energy. The ammonium
is then transferred to organic compounds such as amino acids, in a process known as ami-
nation. These processes occur mostly in the roots. Forms of nitrogen rising toward the
shoot in the xylem are almost all organic, mostly amino acids.
Plants can also take up toxic minerals such as lead or some radionuclides from the
soil. This is being exploited in the phytoremediation of contaminated land (see
Section 16.2.4).
PROBLEMS
1.1. By looking at the Table 7.2, can you find any minerals that may be soil pollutants and
that might be removed from soil by plants? Are there any minerals not in the table
that are more toxic, yet have similar chemical behavior?
1.2. Sometimes industrial sites can be seen to have large areas of soil with few or no
plants. Other than the presence of toxic substances, what could account for this?
PROBLEMS 153
REFERENCES
Fried, G., 1990. Schaum’s Outline: Theory and Problems of Biology, McGraw-Hill, New York.
Raven, P. H., R. F. Evert, and S. E. Eichhorn, 1992. Biology of Plants, Worth Publishers, New York.
Simpson, B. B., and M. C. Ogorzaly, 1995. Economic Botany: Plants in Our World, McGraw-Hill,
New York.
Weier, T. E., C. R. Stocking, and M. G. Barbour, 1970. Botany: An Introduction to Plant Biology,
Wiley, New York.
154
THE PLANTS

8
THE ANIMALS
Animals are multicellular heterotrophs whose cells lack cell walls and that have a motile
stage at some part of their life cycle. They are diploid and reproduce primarily by sexual
reproduction. Some of the int eractions between animals and the environmental effects of
human activities are:
Loss of wildlife h abitat may cause extinction.
Overharvesting may lead to depletion or extinction of species.
Animals may be reservoirs for human diseases.
Control of pests may cause pollution.
The animal kingdom is divided into 33 phyla. A few of them include familiar groups or
organisms. However, quite a few are relatively unfamiliar, and most of these are small
marine organisms. About 1.5 million animal species have been named, but these are prob-
ably only a fraction of those that exist. Table 8.1 summarizes the animal phyla, grouped
according to an informal subdivision system.
8.1 REPRODUCTIVE STRATEGIES
Animals have a great variety of means of reproduction. Only a few of the simplest
animals reproduce asexually. Cnidarians, such as hydra, reproduce by budding, the
unequal division of an organism. Many invertebrates, such as sea stars, can reproduce
by fragmentation, in which the organism can simply be broken in two, with each part
then regenerating the missing portion. Asexual reproduction produces clones, individuals
Environmental Biology for Engineers and Scientists, by David A. Vaccari, Peter F. Strom, and James E. Alleman
Copyright # 2006 John Wiley & Sons, Inc.
155

TABLE 8.1 Summary of Animal Phyla
Phylum Description
Mesozoa Small worms 0.5 to 7 mm long, composed of only 20 to 30 cells; exclusively
marine parasites.
Porifera Sponges, 5000 species (150 freshwater) made of gelatinous matrix with
calcium carbonate or silicate skeleton. Flagellated cells pump as much as
1500 L of water per day to capture food.
Placozoa Contains a single species, Trichoplax adhaerens, a small (2 to 3 mm
diameter) plate like organism, it glides over food, secreting digestive
enzymes, absorbing the products.
Coelenterates (the Only Animals with Radial Symmetry)
Cnidaria Mostly marine, includes jellyfish, corals, anemones, and freshwater hydras.
First phylum with specialized tissues (e.g., nerve cells).
Ctenophora Common name comb jellies. Fewer than 100 species known. Only biradial
phylum.
Flatworms, etc.: More Complex Organ Specialization
Platyhelminthes Flatworms and flukes, turbellarians (e.g., Planaria), trematodes (e.g.,
Schistosomona), tapeworms.
Gnathostomulida Jaw worms: found in sediments, where they tolerate low oxygen levels; feed
on fungi and bacteria scraped from rocks.
Nemertea Ribbon worms: simplest organisms with a circulatory system.
Vermiform (Worm-Shaped)
Rotifera Rings of beating cilia draw food to the mouth, giving impression of rotating
wheels.
Nematoda Roundworms; includes organism that causes trichinosis.
Gastrotricha
Kinorhynca
Loricifera
These resemble rotifers, but without ‘‘wheel’’ of cilia. Together with rotifers
and nematodes, form a group called aschelminthes, and are common
aquatic organisms.
Nematomorpha Horsehair worms, include 250 species that are arthropod parasites.
Acanthocephala Spiny-headed worms; all are internal parasites that cause painful intestinal
injury.
Entoprocta Sessile organisms that superficially resemble hydra, but with ciliated
tentacles.
Priapulida 15 species of cold-water marine worm.
Development of a ‘‘True’’ Body Cavity
Mollusca Bivalves (e.g., clams), octopus and squid, snails and slugs.
Annelida Segmented worms: polychaetes (marine worms), oligochaetes (e.g.,
earthworms, tubifex), leeches.
Arthropoda Insects, crustaceans, spiders.
Lesser Protostomes: Minor Ecological Significance
Echiurida
Sipunculida
Tardigrada Water bears, common in aquatic habitats.
Pentastomida
Onychophora
Pogonophora Includes tube worms from deep-sea vent communities.
156
THE ANIMALS

that are genetically identical to the ‘‘parent.’’ It has the advantage that it can produc e new
individuals rapidly. However, it reduces genetic variablity and the ability of a population
to adjust to drastically changed environmental conditions.
Sexual reproduction is the formation of a new individual from the union of two
potentially genetically distinct gametes. The larger, nonmotile gamete is called the egg,
produced by meiotic division in female sex organs. In animals the smaller motile gam ete
is the sperm, which is produced by meiosis in male sex organs. The haploid gametes
combine in the process of fertilization to form a diploid single cell called the zygote,
which will develop into a new individual.
The most familiar form of sexual reproduction is bisexual reproduction, in which the
male and female organs that produce the gametes are on separate individuals. Species that
use this strategy are called dioecious. Almost all vertebrates, and many invertebrates, are
dioecious. Mating requires time, energy, and coordination and results in offspring that do
not share all the genes of either of its parents. Nevertheless, sexual reproduction confers
on a species a great ability to adjust to environmental change.
Some animals reproduce by parthenogenesis (virgin origin). No fertilization occurs,
although the diploid condition is restored by chromosome doubling. Some flatworms, roti-
fers, insects, and even fish reproduce parthenogenetically. Among the bees, a queen may
allow only some of her eggs to be fertilized. The fer tilized eggs develop into diploid
female worker bees; the haploid males are drones. Certain strains of turkeys have been
found capable of parthenogenetic reproduction. Even in mammals, such as mice, eggs
will on rare occasions start to develop into embryos and fetuses without fertilization.
However, they have never been found to survive to full term.
Some animals are hermaph rodites, having both male and female sex organs on the
same individual. They are also referred to as monoecious. This is a reproduction strategy
that is appropriate for organisms that are sessile, such as barnacles, or live in burrows or
parasitically within other organisms, such as the tapeworm. The advantage of this is that it
makes it possible for every individual to bear young, not just half the population. Some
hermaphroditic fish actually start out as a member of one sex, then change to the other
during their life span.
Animal Weeds: Sessile Wormlike Marine Animals
Phoronida Only 10 known species.
Ectoprocta Moss animals.
Brachiopoda The ‘‘lamp shells’’, resemble bivalves.
Deuterostomes
Chaetognatha Arrowworms: wormlike planktonic predators, 2.5 to 10 cm long.
Hemichordata Wormlike organisms.
Echinodermata Sea stars, sea urchins, etc.
Higher Animals
Chordata Tunicates, lancelets, vertebrates.
TABLE 8.1 (Continued)
Phylum Description
REPRODUCTIVE STRATEGIES 157